激光和LED补光对番茄幼苗生长、光合作用及生理生化特性的影响
2022-05-24陈美香申宝营邱雯婷林火养刘银春
陈美香,陈 雄,申宝营,邱雯婷,王 洪,郭 建,林火养,刘银春
(1. 福建信息职业技术学院物联网与人工智能学院,福建 福州 350001;2. 福建农林大学机电工程学院,福建 福州 350002;3. 福建江夏学院电子信息科学学院,福建 福州 350108;4. 福建农林大学园艺学院,福建 福州 350002)
0 引言
【研究意义】番茄育苗是番茄生产过程中的关键技术环节,幼苗的品质决定后期果实的产量和质量。传统的设施番茄育苗全靠太阳光照,光强极不稳定,特别是我国南方常遇阴雨天气,幼苗因缺乏光照长势不佳。工厂化育苗是现代农业发展的必然趋势,它利用人工光源,在人工控制的最佳环境条件下育苗,是培育番茄壮苗的有效途径之一。找到最佳的光配方是番茄工厂化育苗的关键。【前人研究进展】光是影响植物生长的最重要的环境因子之一,它影响植物光合作用、形态建成、生理生化和基因表达[1-3]。研究表明,光质和光强作为光合作用的能源影响叶绿体结构、光合色素合成和气孔运动[4,5];同时,光质还作为信号影响植物的生长和生理生化。例如,红光可以激发叶绿素分子进行光合作用,蓝光既可以激发这种作用又可以触发隐花色素和向光素,进而影响植物的形态建成和其他生理活动[6-8]。有研究发现,红光有利于提高草莓叶片的净光合速率与蒸腾速率[5];蓝光有利于增大草莓叶片的气孔导度与胞间CO2浓度[5]、增加竹叶兰过氧化氢酶与过氧化物酶含量,增强竹叶兰抗氧化能力[3];红蓝黄复合光有利于提高紫苏叶片的光合速率[9],红、蓝和白光的优化组合能促进金线莲的生长和提高黄酮含量[10]。近年来,发光二极管(Light-emitting diode,LED)因单色性好、光强易调等优点被广泛应用于植物生长、光合作用和生理生化研究[11-13]。激光是单色性最好的光源,和LED相比,光谱能量更集中、光辐照距离更远、光能在空气中损失更小。因此,激光特别适合在大田、温室或植物工厂等大面积种植环境中补光。已有研究表明,激光促进草莓光合作用使草莓植株的重量指数增加[14];低剂量He-Ne激光照射能有效地缓解UV-B 辐射对小麦、水稻和花生幼苗叶绿体产生的胁迫损伤从而促进光合作用[15-17];Nd:YAP红外激光可刺激亚心形扁藻的叶绿体发育,改善其光合作用活性[18]。【本研究切入点】现有对番茄育苗光环境调控的研究大部分使用荧光灯、LED灯等,用激光做补光光源鲜有报道。【拟解决的关键问题】本文以番茄幼苗为研究材料,先探索各3种单色激光和LED光(2红1蓝)辐照下叶片的光合作用特性,选出幼苗光合能力较强的红光,再将其和蓝光作为补光光源设计3个处理,进行番茄补光育苗试验,以未补光的LED白光育苗为对照,探明不同补光方法对番茄幼苗生长、光合作用及生理生化特性的影响,找到最佳的番茄育苗补光方案,为提高番茄幼苗品质提供科学依据。
1 材料与方法
1.1 测试材料
供试番茄(Solanum lycopersicum)选用白果强丰品种,试验分两个阶段。第一阶段:测定各3种激光和LED单色光辐照下番茄幼苗叶片的光合响应曲线。2018年6月1日播种,置暗室内4 d后幼苗出土,移至LED光下培养。距育苗盘(32孔穴)底部13.4 cm处的光合光量子通量密度(PPFD)为 100 μmol·m-2·s-1, 光 照 时 间 12 h·d-1, 室 内 温 度 (25 ±2)°C、相对湿度(70 ± 10)%。30 d后选 30株一致性好的植株作为供试植株,10株为1个重复,共3个重复,每个重复选一株测定光合响应曲线。第二阶段:色温6 000 K的LED白光补激光和LED光育苗试验。选相同的番茄品种,2020年7月1日播种,置暗室内4 d后幼苗出土,以白光补少量蓝光和第一阶段选出的光合能力较强的红色激光和LED光作补光光源,设计3个光处理(表1),以未补光的LED白光育苗为对照。光照时间、温度和湿度条件同第一阶段,距育苗盘(32孔穴)底部13.4 cm处的 PPFD = 88 μmol·m-2·s-1。栽培 30 d 后,设置 3个重复(方法同第一阶段),测定生长、光合作用和生理生化指标。
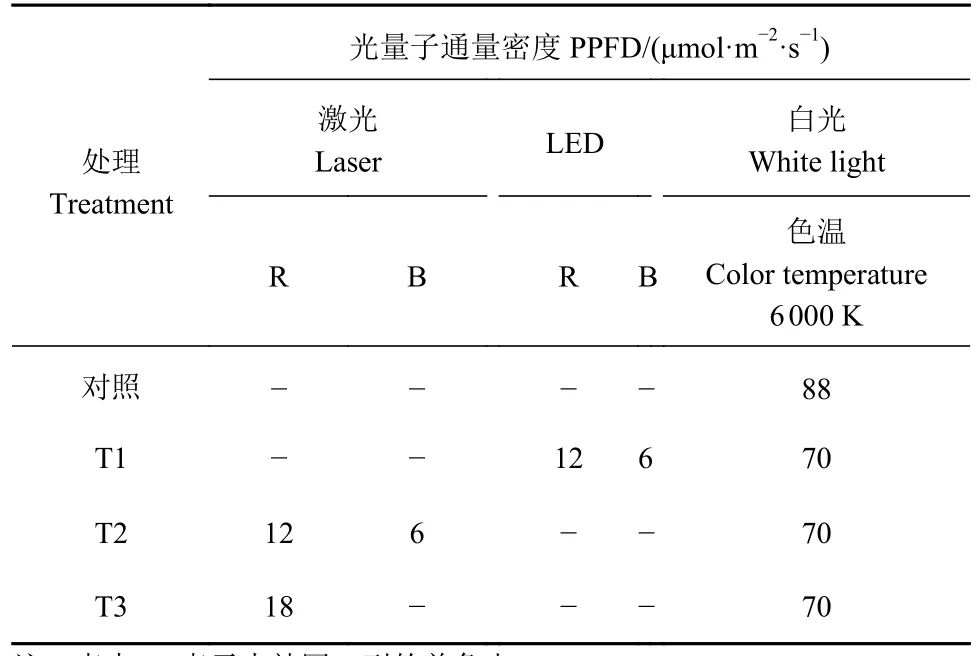
表1 激光和LED补光栽培试验光源设计Table 1 Design of supplementary laser and LED light for tomato cultivation experiment
1.2 测定方法
选择第一阶段培养的植株自上而下第3片叶片,使用CIRAS-3光合测定系统(美国,PPSYSTEMS公司),分别测定3种激光(波长450 nm、633 nm、650 nm,半波宽8~10 nm)和3种单色LED光(波长450 nm、630 nm、650 nm,半波宽18~22 nm)辐照下,光强 PPFD(μmol·m-2·s-1)分别为 0,20,40,60,80,100,150,200,250,300,400,···,最大值时,其对应的净光合速率(Pn)、气孔导度(Gs)、 胞 间 CO2浓 度 (Ci)、 蒸 腾 速 率 (Tr)和水分利用率(WUE)。测试环境温度(26 ± 2) ℃,CO2浓度(450 ± 30) μmol·mol-1。各光源下Pn值稳定后开始测量,测量时每5 s记录1次,共5次,取平均值。
第二阶段栽培结束,同样选择植株自上而下第3片叶片,采用光合测定系统自带的光源,调至红光:绿光:蓝光=1:1:1,总PPFD的梯度和第一阶段相似,从0至Pn的饱和光强,依次测定各光合作用参数值。
采用WinRHIZO根系分析系统(加拿大Regent)测定叶面积。定义壮苗指数和根冠比如下:

叶绿素和类胡萝卜素含量测定采用Arnon[19]的方法。超氧化物歧化酶(SOD)活性、过氧化物酶(POD)活性、丙二醛(MDA)含量测定分别采用WST-1 法、比色法和TBA法(试剂盒购于南京建成生物工程研究所)。定义在37 °C条件下,SOD抑制率达50%时所对应的酶量为一个SOD活力单位(U),每克叶片样本每分钟催化1微克底物的酶量为一个POD活力单位(U)。叶绿素、类胡萝卜素含量和POD活性使用UV-2600型紫外可见分光光度计(日本,岛津公司)测定,SOD活性和MDA含量使用DNM-9602酶标分析仪(北京普朗)测定。
1.3 数据处理
采用Excel2010软件进行数据处理和作图分析,采用SPSS Statistics17.0统计分析软件进行统计和差异显著性检验。
2 结果与分析
2.1 激光和LED光辐照对番茄幼苗叶片光合特性的影响
不同激光和LED光质辐照番茄幼苗叶片的光合特性参数值存在差异(图1)。由图1-A可以看出,相同光强,激光辐照幼苗的净光合速率Pn(650 nm)>Pn(450 nm)>Pn(633 nm);LED辐照幼苗 当 PPFD< 250 μmol·m-2·s-1时Pn(650 nm)>Pn(630 nm)>Pn(450 nm),当 250 μmol·m-2·s-1<PPFD ≤ 400 μmol·m-2·s-1时Pn(650 nm)≈Pn(630 nm)>Pn(450 nm), 当 PPFD > 400 μmol·m-2·s-1时Pn(630 nm)>Pn(650 nm)>Pn(450 nm)。激光辐照幼苗的Pn(650 nm)和Pn(450 nm)均大于相同波长的LED辐照,且这两波长激光辐照和LED辐照的比值最大分别达到3.5和1.7倍(PPFD = 300 μmol·m-2·s-1时),激光辐照幼苗的Pn(633 nm)在PPFD 大于 150 μmol·m-2·s-1之后小于对应波长的 LED辐照。不同光质,激光辐照幼苗的光补偿点均低于对应波长的LED辐照。所有红光辐照幼苗的光饱和点均为 700 μmol·m-2·s-1,450 nm 激光辐照幼苗的光饱和点为 800 μmol·m-2·s-1,低于同波长 LED 辐照的饱和点 1 100 μmol·m-2·s-1。
激光和LED辐照番茄幼苗叶片的胞间CO2浓度Ci(λ)随光强增加先快速下降,在PPFD达到700 μmol·m-2·s-1后,所有红光和 450 nm 激光辐照幼苗的Ci慢慢趋于稳定,但450 nm LED辐照幼苗的Ci继 续 下 降 (图 1-B)。 PPFD ≤ 400 μmol·m-2·s-1时,激光辐照幼苗的Ci(650 nm)>Ci(633 nm)>Ci(450 nm),但LED辐照幼苗的Ci(450 nm)>Ci(630 nm)>Ci(650 nm);PPFD>400 μmol·m-2·s-1后,激光辐照幼苗的Ci(633 nm)>Ci(650 nm),但LED辐照幼苗还是Ci(630 nm)>Ci(650 nm)。

图1 不同光处理下番茄幼苗叶片光合作用的光响应曲线Fig. 1 Light response curves on leaf photosynthesis of tomato seedlings
气孔导度Gs(λ)均随光强增加而增加,但激光辐照幼苗Gs(650 nm)和Gs(450 nm)在最高光强时有所下降(图1-C)。相同光强,除450 nm外,激光辐照幼苗Gs(λ)均大于对应波长的LED辐照,且激光辐照幼苗Gs(650 nm)>Gs(633 nm)>Gs(450 nm);LED 辐照幼苗在 PPFD ≤ 500 μmol·m-2·s-1时,Gs(450 nm)>Gs(630 nm)和Gs(650 nm),在PPFD>600 μmol·m-2·s-1时,Gs(650 nm)>Gs(630 nm)>Gs(450 nm)。幼苗的蒸腾速率Tr(λ)和水分利用率WUE(λ)随光质、光强的变化规律分别与气孔导度Gs(λ)和净光合速率Pn(λ)非常相似(图 1-D、E),可见,Tr(λ)和WUE(λ) 分别与Gs(λ)和Pn(λ)呈正相关关系。
综合以上分析,650 nm激光辐照番茄幼苗叶片的光合能力大于633 nm辐照,650 nm LED辐照番茄幼苗叶片的光合能力大于(PPFD <250 μmol·m-2·s-1)和约等于(250 μmol·m-2·s-1≤ PPFD ≤ 400 μmol·m-2·s-1)630 nm辐照,故以650 nm激光和LED光为红光补光光源。
2.2 激光和LED补光对番茄幼苗生长的影响
以白光补少量650 nm和450 nm激光和LED光为育苗光源,设计3个光处理培育幼苗,以LED白光培育幼苗为对照(表1)。结果表明(表2),各生长指标在处理组间存在显著差异(P<0.05)。T2处理的番茄幼苗除株高和地上部鲜重外,其余生长指标值都最高;T3处理的番茄幼苗株高最高但叶面积最小,幼苗发生徒长;T1处理的番茄幼苗株高、地下部鲜重和干重最小。对照幼苗茎直径、地下部鲜重和干重最小。壮苗指数和根冠比在处理组间也存在显著差异(P<0.05),所有试验组的壮苗指数值和根冠比从大到小排列分别为T2(8.8 mg)>T1(5.5 mg)>T3(4.0 mg)>对照(3.7 mg)和 T2(0.13) >T3(0.09)>对照(0.08)>T1(0.07)。

表2 西红柿幼苗的生长指标Table 2 Growth indicators of tomato seedlings
2.3 激光和LED补光对番茄幼苗叶片光合参数的影响
3个处理叶片的净光合速率Pn在光强超过光补偿点后快速增大、光强达到 200 μmol·m-2·s-1后缓慢增加到各自的光饱和点后趋于稳定(图2-A)。所有试验组光补偿点都是 20 μmol·m-2·s-1,光饱和点 T2(800 μmol·m-2·s-1)< 对照、T1(1 000 μmol·m-2·s-1)<T3(1 400 μmol·m-2·s-1)。T2 处理幼苗的最大净光合速率(光饱和点对应的净光合速率)分别是对照、T1、T3处理的1.5倍、1.3倍、1.2倍。
幼苗叶片胞间CO2浓度Ci随光强变化的规律和Pn相反(图2-B),先随光强增加快速下降,在光强达到 200 μmol·m-2·s-1后缓慢下降并趋于稳定(除了T3处理的Ci缓慢下降后缓慢增加、T2处理的继续呈下降趋势)。所有试验组Ci在各光强下差异较小。
气孔导度Gs和蒸腾速率Tr随光强的变化规律为在 PPFD < 800 μmol·m-2·s-1时相似(图 2-C,D),都是随光强增加而增加,当 100 μmol·m-2·s-1< PPFD < 800 μmol·m-2·s-1时,所有试验组按Gs和Tr值从大到小排列依次是对照>T1>T2>T3;在强光(PPFD>1 000 μmol·m-2·s-1)辐照下,T3 处理的Gs和Tr继续较快地增加,其他试验组Gs缓慢下降、Tr缓慢增加后下降,其中T2的Gs下降最快。
水分利用率WUE随光强的变化规律和净光合速率Pn相似(图2-E),在较低光强(PPFD < 400 μmol·m-2·s-1)时随光强快速增加、中等光强(400~800 μmol·m-2·s-1)时 缓 慢 增 加 (除 了 T3 处 理 在PPFD> 600 μmol·m-2·s-1时 开 始 下 降 )、 高 强 光(PPFD>800 μmol·m-2·s-1)时缓慢下降并趋于稳定(除了T2处理下降后又上升)。T2处理WUE最大值分别是对照、T1、T3处理最大值的1.6、1.4、1.1倍。

图2 不同光处理下番茄幼苗叶片光合作用的光响应曲线Fig. 2 Light response curves on photosynthesis of tomato seedlings under treatments
2.4 激光和LED补光对番茄幼苗生理生化特性的影响
3个光处理对番茄幼苗叶片的光合色素含量的影响存在差异(图3-A)。白光补激光(T2、T3)处理幼苗的叶绿素a、类胡萝卜素含量显著大于未补激光处理(P<0.01),但叶绿素b含量则无显著差异(P>0.05);白光补LED光(T1)处理的叶绿素a含量也显著大于对照,但其他色素含量无显著差异;T3、T2处理的叶绿素a含量无显著差异(P>0.05),分别是对照(叶绿素a含量最低)的1.7、1.6倍;叶绿素a/b的比值存在显著差异(P<0.01),按比值从高到低依次为T3(5.6)>T2(5.5)>T1(2.0)>对照(1.7)。T2处理的类胡萝卜素含量最高,分别是对照、T1和T3处理的7.0倍、7.7倍和2.0 倍。
3个光处理对番茄幼苗叶片中SOD、POD酶活性和MDA含量的影响存在差异(图3-B)。3个处理幼苗的SOD活性无显著差异(P>0.05),但POD活性和MDA含量有极显著差异(P<0.01)。所有补激光和LED光处理的POD活性和MDA含量显著大于对照,其中T2处理的值最大; T2、T3 和T1处理的POD活性分别是对照的1.5倍、1.4和1.3倍,MDA含量是对照的8.6倍、3.5倍和2.4倍。

图3 不同光处理番茄幼苗的生理生化指标Fig. 3 Physiological and biochemical indices of tomato seedlings under treatments
3 讨论与结论
植物在不同的光照条件下会调整自身的生理状态以获得最佳的光合作用[20]。在弱光条件下,植物的光合速率或CO2同化速率较低;当环境光强超过植物光合作用的光补偿点后,光合速率随着光强的增加而增加,植物开始积累有机物质并生长;在最佳光质和光强下,植物处于最佳生长和代谢状态,代谢产物含量最高[21]。
本研究的结果表明650 nm和450 nm激光是最佳的补光光源,它们优于同波长LED光;白光同时补少量这两种波长的激光进行育苗能大大提高幼苗的光合能力。Hall等[22]研究表明,叶绿素a和叶绿素b的吸收光谱在蓝光波段和红光波段各有2个吸收峰,蓝光波段峰值波长是430 nm和453 nm,红光波段是643 nm和660 nm。激光单色性比LED光好,其光能比LED更集中于中心波长,因此,同光强的450 nm激光比LED光有更多光子处于叶绿素的蓝光吸收峰,光合作用效率更高。650 nm处于红光波段叶绿素的2个吸收峰之间,同光强的激光和LED光,激光有更多光子远离两吸收峰,但它仍能产生比LED光高得多的净光合速率,说明650 nm激光除了为光合作用提供能量,还有其他作用,如可以提高叶绿体电子传递速率,激活叶绿体酶活性,从而提高光合能力。这和高丽美等[23]对小麦的研究结果一致。因此,植物生长的最佳光源的光谱,应和植物光合作用的吸收光谱匹配,而这吸收光谱应是光合色素(主要是叶绿素a/b)的吸收谱和光合作用酶等能促进光合作用的吸收光谱的叠加。本研究还表明白光同时补650 nm和450 nm激光处理光饱和点比其他试验组低,而最大净光合速率却比其他试验组高很多,说明同时补650 nm和450 nm激光能大大提高幼苗的光合作用光能利用率。
本研究还表明,白光补少量激光和LED光都能显著提高叶绿素a含量和叶绿素a、叶绿素b的比值a/b,但同时补650 nm和450 nm激光的效果更佳。光是叶绿体发育和叶绿素合成必不可少的条件,在叶绿素a的合成过程中,有2个直接需光过程:谷氨酰-tRNA(Glu-tRNA)在光照的刺激下生成δ-氨基乙酰丙酸、原叶绿素酸酯经光还原成叶绿酸酯[24]。此外,一些酶的激活需要光。因此,本文推测激光更能促进这些需光反应,进而促进叶绿素a的合成。本研究的第一阶段番茄幼苗叶片在450 nm激光辐照下的净光合速率远小于650 nm激光辐照,但第二阶段白光同时补这两种波长激光后,叶片净光合速率显著大于白光补650 nm激光,这说明单独450 nm激光辐照叶片产生了光胁迫,抑制了光合作用,但增加650 nm激光后缓解了光抑制,提高了光合能力。这很可能是650 nm激光修复了番茄叶片光合系统由于光抑制造成的损伤。这与张美萍等[16]研究结果相似。本文中白光同时补650 nm和450 nm激光还能显著提高幼苗的生长指标,这正是光合能力提高的结果,和陈晓栋等[14]对草莓的研究结果相似。
类胡萝卜素是植物中普遍存在的光合色素,它通过与其他有机物特别是叶绿素a、叶绿素b的直接相互作用,参与光的收集和能量传递,并通过消耗多余能量来防止光合系统的光氧化损伤[25]。此外,植物在受到干旱等环境胁迫时叶片中的类胡萝卜素含量会升高[26]。本研究中白光补激光特别是650 nm和450 nm激光能显著提高类胡萝卜素含量,这与金丽虹[27]对大豆幼苗的研究结果一致。说明激光特别是具有高能光子的蓝色激光(450 nm)促进了类胡萝卜素的合成,可能是因为番茄幼苗通过合成更多类胡萝卜素来防止激光对光合系统造成光氧化损伤,是其应对激光胁迫的生存策略。
SOD、POD是植物保护酶体系的关键组成部分,是植物体内主要的抗氧化酶,其中SOD能催化超氧化物阴离子自由基发生歧化反应生成O2或H2O2,消除自由基保护细胞膜;POD能清除活性氧,催化过氧化氢的分解,防止膜脂过氧化和过氧化氢毒害细胞;MAD是反映膜脂过氧化程度的重要指标,其含量高低反映膜脂过氧化程度大小。本研究表明,白光补激光和LED光都能显著提高番茄幼苗POD活性和MAD含量,提高其抗氧化能力,但白光同时补650 nm和450 nm激光效果更显著,说明同时补红蓝两种激光虽加速了膜脂过氧化,但促使番茄幼苗保护酶系统活性升高,加快细胞清除自由基的速度以减轻膜系统的受损程度。这和对大豆幼苗[27]、青钱柳[28]、曼地亚红豆杉[29]、远志[30]等的研究结果一致。
综上所述,白光补650 nm和450 nm的激光能提高番茄幼苗的生长指标、光合能力和抗氧化酶活性,是适合番茄幼苗生长的优质光源,可用于生产中做补光光源。
