生物信息学分析萝卜硫素上调RAB7交互溶酶体蛋白与非小细胞肺癌细胞自噬的关联
2022-05-21张彤,周妍,吴巍
张 彤,周 妍,吴 巍
(首都医科大学基础医学院生物化学与分子生物学系,北京 100069;*通讯作者,E-mail:weiwu207@ccmu.edu.cn)
萝卜硫素(sulforaphane,SFN)是一种从十字花科植物中提取具有抗癌潜力的化合物[1]。研究表明,在非小细胞肺癌(non-small cell lung cancer,NSCLC)中,SFN能触发微管断裂,抑制自噬溶酶体的形成导致细胞凋亡,但SFN抑制自噬溶酶体形成的机制尚未明确[2]。
研究表明,自噬参与囊泡的形成、运输和融合,而膜转运蛋白作为介导溶质、离子或药物跨生物膜选择性转运的重要跨膜蛋白,与囊泡运输和融合密切相关[3]。其中,RAB7蛋白作为转运蛋白,在细胞内的准确定位决定其所介导的囊泡能否正确抵达目的细胞器。RAB7交互溶酶体蛋白(RILP)是RAB7蛋白下游的效应蛋白,定位在溶酶体[4]。RAB7与GTP结合将RILP运送到细胞膜中,共同控制晚期内体和溶酶体的转运[5,6]。
溶酶体的功能与溶酶体膜蛋白有着密切的关系。其中,ATP6V0D1为溶酶体膜蛋白,具有质子泵的作用,为溶酶体水解酶提供酸性环境,影响溶酶体功能进而调控自噬途径[7]。我们推测,SFN通过调控RILP及溶酶体膜蛋白ATP6V0D1抑制自噬溶酶体的形成。本研究通过生物信息学结合免疫荧光染色及蛋白免疫印迹技术研究SFN对RILP及ATP6V0D1的调控作用,探讨在非小细胞肺癌中SFN抑制自噬溶酶体形成的分子机制,以便为肿瘤靶向治疗提供有效策略。
1 材料与方法
1.1 细胞株与试剂
人非小细胞肺癌细胞株A549细胞购自中国医学科学院基础医学研究所基础医学细胞中心;SFN购自美国Santa Cruz公司,β-actin抗体购自美国Cell Signaling Technology公司;RILP抗体、LAMP1抗体购自美国Proteintech公司。
1.2 激光扫描共聚焦显微镜分析
将处于对数生长期的A549细胞接种于免疫荧光专用的共聚焦小皿内。待共聚焦小皿中A549细胞密度达到60%左右后,弃去旧培养基,置换成新鲜的含10%胎牛血清的培养基,对照组不做任何处理,实验组加入SFN,使其终浓度为20 μmol/L,诱导细胞24 h。弃去旧培养基,经固定、透化、封闭操作后,根据实验要求将抗RILP抗体(1 ∶200)、抗LAMP1抗体(1 ∶200)加入共聚焦小皿在4 ℃冰箱静置孵育过夜。随后加入二抗室温避光孵育1 h,然后滴加三到四滴含DAPI的防荧光淬灭封片剂封片,于湿润避光处保存。在激光扫描共聚焦显微镜下随机寻找视野观察目的蛋白的荧光强度及定位,并拍照。
1.3 高效液相色谱与质谱联用分析蛋白表达差异
用高效液相色谱和质谱联用分析SFN处理后肺腺癌细胞A549中蛋白差异表达。将肺腺癌A549分为SFN组与未处理组(对照组)。使用20 μmol/L SFN处理肺腺癌细胞A549 24 h,收集细胞裂解液并定量。对照组与SFN组使用等量的蛋白上样,每组样品有3个重复样本。通过Orbitrap Fusion Lumos质谱仪和EASY-nLC 1000液相色谱系统进行色谱分离和分析,该系统配备了电喷雾电离源。使用C18(1.9 μm,100 A)毛细管柱,流动相为0.1%甲酸/水和0.1%甲酸/乙腈进行分离,并以正电离模式进行检测。收集数据后,通过Uniprot网站分析蛋白质的细胞定位和功能。
1.4 蛋白免疫印迹技术检测RILP蛋白的表达
分别加入0,10,20,30 μmol/L SFN处理A549细胞24 h后,使用含有蛋白酶抑制剂的RIPA缓冲液(Thermo Fisher Scientific,USA)裂解细胞,通过12% SDS-PAGE电泳分离对应样品中等量的蛋白质分子(20 μg),根据目的蛋白分子量观察对应蛋白Marker的分离情况,一般在溴酚蓝到达分离胶底部时转膜,最后使用Odyssey Software Version启动成像系统检测不同浓度SFN处理后RILP(1 ∶1 000),β-actin(1 ∶5 000)蛋白的表达。
1.5 生物信息学分析
使用UALCAN数据库分析肺癌与癌旁组织蛋白差异表达;用ONCOLNC数据库分析肺癌患者中RILP、ATP6V0D1蛋白表达与生存期的关联;用STRING数据库(10.5版)来分析RILP与微管蛋白TUBB/TUBA1A、LC3Ⅱ之间的相互作用;用GEPIA数据库分析RILP、ATP6V0D1、LC3Ⅱ蛋白间的相关性。
1.6 数据处理和统计学检验
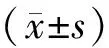
2 结果
2.1 SFN诱导溶酶体核周聚集
免疫共聚焦荧光显微镜观察显示:对照组溶酶体散在、均一分于细胞质中;SFN组SFN处理细胞后溶酶体团成一簇聚集在核周(见图1)。

红色荧光表示溶酶体相关膜蛋白LAMP1;蓝色荧光表示细胞核染料DAPI图1 SFN对溶酶体核周聚集的影响Figure 1 Effect of SFN on the perinuclear aggregation of lysosomes
2.2 SFN诱导上调的蛋白富集到膜转运相关通路
液相色谱和质谱联用分析表明:在SFN处理的A549细胞中,380个蛋白表达上调(红色),同时278个蛋白表达下调(绿色),差异有统计学意义(P<0.05,见图2A);利用Uniprot网站预测这些差异蛋白的功能与亚细胞定位,同时对上调的蛋白进行聚类分析发现SFN处理后蛋白富集到膜转运相关通路(见图2B)。

A.火山图分析蛋白表达差异 B.Uniprot网站中蛋白聚类分析图2 液相色谱和质谱联用分析差异蛋白的功能Figure 2 Function of differential proteins by liquid chromatography and mass spectrometry
2.3 SFN上调RILP及RILP与微管蛋白的相互作用
使用蛋白免疫印迹技术检测SFN处理A549细胞后RILP蛋白的表达情况,发现随着SFN浓度的升高RILP的表达水平逐渐上升,差异有统计学意义(P<0.05,见图3A),表明SFN对RILP表达的调节呈浓度依赖性。同时该蛋白与微管蛋白TUBB/TUBA1A有相互作用(见图3B)。

与0 μmol/L比较,*P<0.05B.STRING数据库中PPI网络分析A.SFN上调A549细胞中RILP蛋白表达图3 SFN处理后RILP的表达及RILP与TUBB/TUBA1A蛋白表达关联Figure 3 The expression of RILP after SFN treatment and the association of RILP with TUBB/TUBA1A protein
2.4 RILP的表达与肺癌患者生存率有相关性
TCGA数据分析显示RILP在肺腺癌和肺鳞癌组织中较正常组织表达显著降低(P<0.05,见图4A)。且ONCOLNC数据库显示高表达RILP的肺癌患者生存率高(P<0.05,见图4B)。

A.TCGA数据库中RILP蛋白的表达B.ONCOLNC数据库中NSCLC患者生存曲线分析图4 RILP在肺癌中的表达及RILP蛋白高表达的肺癌患者生存曲线Figure 4 The expression of RILP in lung cancer and the survival curve of lung cancer patients with high RILP expression
2.5 RILP与自噬相关蛋白LC3Ⅱ的相关性
STRING数据库显示,RILP与自噬相关蛋白LC3Ⅱ(基因名:MAP1LC3B)存在相互作用(见图5A),TCGA数据库分析显示在肺鳞癌与肺腺癌中RILP与LC3Ⅱ的表达呈正相关(肺腺癌:r=0.20,P<0.01;肺鳞癌:r=0.37,P<0.01,见图5B)。
2.6 肺癌中RILP与ATP6V0D1的相关性
TCGA数据库显示在肺鳞癌和肺腺癌中RILP与ATP6V0D1呈正相关(肺腺癌:r=0.37,P<0.01;肺鳞癌:r=0.54,P<0.01,见图6)。
2.7 ATP6V0D1上调自噬相关蛋白LC3Ⅱ的表达
ONCOLNC数据库显示ATP6V0D1在肺腺癌和肺鳞癌中表达较正常组织显著降低(P<0.05),且ONCOLNC数据库显示ATP6V0D1高表达的患者生存率显著提高(P<0.05,见图7)。

A.STRING数据库中PPI网络分析B.TCGA数据库中基因相关性分析图5 肺癌患者中RILP与LC3Ⅱ的相关性分析Figure 5 Correlation between RILP and LC3Ⅱ in lung carcinoma patients

A.肺腺癌B.肺鳞癌图6 TCGA数据库分析RILP与ATP6V0D1在肺腺癌和肺鳞癌中的相关性Figure 6 Correlation between RILP and ATP6V0D1 in lung adenocarcinoma and lung squamous cell carcinoma by TCGA database

B.ONCOLNC数据库中NSCLC患者生存曲线分析A.TCGA数据库中ATP6V0D1蛋白表达分析图7 ATP6V0D1在肺癌的表达及ATP6V0D1高表达的肺癌患者生存曲线Figure 7 The expression of ATP6V0D1 in lung cancer and the survival curve of lung cancer patients with high ATP6V0D1 expression
2.8 肺癌中ATP6V0D1与自噬相关蛋白LC3Ⅱ的相关性
TCGA数据库显示在肺鳞癌和肺腺癌中,ATP6V0D1与自噬标志蛋白LC3Ⅱ呈正相关(肺腺癌:r=0.48,P<0.01;肺鳞癌:r=0.57,P<0.01,见图8)。

A.肺腺癌患者基因相关性分析B.肺鳞癌患者基因相关性分析图8 TCGA数据库分析肺腺癌和肺鳞癌患者ATP6V0D1与LC3Ⅱ的相关性分析Figure 8 Correlation between ATP6V0D1 and LC3Ⅱ in lung adenocarcinoma and lung squamous cell carcinoma by TCGA database
3 讨论
既往研究发现SFN是一种有潜力的抗癌活性物质,通过调节自噬诱导凋亡并能抑制多种肿瘤细胞增殖、浸润[8]。本研究我们进一步发现SFN可以上调RILP的表达,增强RILP与溶酶体ATP6V0D1蛋白及LC3Ⅱ的关联,从而抑制非小细胞肺癌细胞自噬体与溶酶体融合。这些研究结果对于我们分析SFN调节的自噬机制及其潜在的抗癌靶点提供了新的思路。
自噬是一种重要的细胞降解过程,其中功能失调的细胞质内容物被溶酶体水解酶消化。在自噬过程中,伴随着一些囊泡的融合,包括自噬体和溶酶体的融合,导致自噬溶酶体中的内容物降解和清除[9]。实验室前期结果显示,SFN及其代谢物可以诱导肿瘤细胞形成自噬体,同时抑制自噬体与溶酶体的融合,阻碍肿瘤细胞内的错误折叠蛋白和受损细胞器的降解,从而诱导肿瘤细胞凋亡[10]。
RILP蛋白作为溶酶体膜蛋白,通过与其上游蛋白RAB7相互作用,调节核内体和溶酶体之间的信号转导并参与自噬小体的成熟,同时有文献报道其在自噬溶酶体形成中也扮演着至关重要的作用[11]。生物信息学分析显示RILP高表达的肺癌患者生存率显著升高,这意味着RILP是一个抑癌蛋白,上调RILP可能成为治疗肺癌的新方向。蛋白免疫印迹结果显示:SFN上调RILP蛋白的表达;同时免疫荧光实验发现SFN诱导溶酶体在核周聚集,这提示我们在非小细胞肺癌细胞中,SFN可能通过调节RILP蛋白诱导溶酶体向核周聚集。研究表明,SFN靶向细胞中多种蛋白,其中以α-tubulin蛋白为主。SFN可以与α-tubulin共价结合,破坏其二级和三级结构,抑制微管聚合,从而诱导肿瘤细胞凋亡[12]。为了研究RILP蛋白与自噬之间的关联,我们使用TCGA数据库分析RILP蛋白与自噬相关蛋白LC3Ⅱ的关系,发现RILP蛋白可与LC3Ⅱ、tubulin相互作用,这进一步证明在非小细胞肺癌中,RILP蛋白能够调控自噬溶酶体形成,并且微管蛋白tubulin可能调控RILP蛋白介导的自噬。
溶酶体作为细胞内的“清道夫”,可以与自噬体融合,从而消化细胞内受损的细胞器,与溶酶体膜上蛋白质复合体V-ATPase密切相关。研究发现,V-ATPase具有质子泵的活性,能为溶酶体水解酶提供酸性环境,从而维持溶酶体正常功能[13]。有文献报道RILP可以通过调控V-ATPase进而影响溶酶体的功能,而ATP6V0D1作为V-ATPase中的一种亚基,在膜转运以及膜融合方面扮演着重要的角色[14,15]。生物信息学分析显示ATP6V0D1与LC3Ⅱ在肺腺癌和肺鳞癌的表达均呈正相关,LC3Ⅱ蛋白作为一种微管相关蛋白,主要存在于自噬体膜上,并在自噬溶酶体的形成中发挥作用,这提示我们ATP6V0D1在自噬溶酶体形成中也扮演着重要的角色。同时,ATP6V0D1高表达的肺癌患者生存率显著升高,这表明ATP6V0D1是一个抑癌蛋白,可能成为治疗肺癌的新靶点。
综上所述,我们证明SFN在亚细胞水平抑制自噬溶酶体形成的机制,这些结果可以为开发新的抗癌疗法提供理论支持。
