土壤动物肠道微生物多样性研究进展
2022-05-20ChenTingWen吴东辉
郝 操,Chen Ting-Wen,吴东辉,*
1 东北师范大学环境学院;国家环境保护湿地生态与植被恢复重点实验室,长春 130117 2 中国科学院东北地理与农业生态研究所;湿地生态与环境重点实验室,长春 130102 3 Biology Centre of the Czech Academy of Sciences,Institute of Soil Biology,Na Sádkách 7,37005 é Budějovice,Czech Republic
动物肠道是一个复杂而又稳定的微生态系统,其内定居着包括细菌、病毒、真菌及一些小型原生动物、显微藻类等生物群体[1]。肠道微生物与宿主互相依存并互相制约,对动物宿主健康发挥着重要作用[2],同时肠道微生物又是联系动物参与生态系统物质循环与能量流动的重要纽带[3—4]。近年来动物肠道微生物研究已从肠道微生物多样性组成[3],拓展到肠道微生物驱动机制[5]及其群落构建过程[6],以及肠道微生物与宿主生长、繁殖、寿命及疾病等方面之间联系和作用机制研究[7—8],备受各界科学家的关注。随着高通量测序手段及组学技术的发展,应用分子生物学手段以快速、定性和定量研究动物肠道微生物,已成为微生物生态学研究热点领域。
土壤动物指生活史有一段时间稳定地在土壤中度过并且对土壤有一定影响的无脊椎动物[9],是研究陆域生态系统土壤生物学效应、土壤健康水平、生态系统演替或受干扰程度的重要材料[7,10—11]。土壤动物及其共生微生物构成一个独特微生态系统,这一独特微生态系统与土壤动物宿主相互作用、协同进化。鉴于土壤动物肠道微生物在与宿主长期协同进化中形成了复杂的群落结构,其组成变化受宿主种类、生理状态、发育阶段、食性、栖息环境等因素共同塑造[12—14]。肠道微生物则影响着土壤动物个体食物消化、营养吸收、繁殖、寿命及免疫调控[4,15—16]与种群和群落的稳态和功能实现。因此,土壤动物肠道菌群能够间接调控陆地生态系统碳氮循环过程[3]。基于目前影响土壤无脊椎动物肠道菌群的主要因素,阐明不同因素对土壤动物肠道菌群多样性和群落结构组成的影响,对理解土壤动物肠道微生物多样性及其对土壤生态系统功能的影响和肠道菌群与宿主的互作机制有着重要意义。
本文总结了土壤动物肠道微生物领域近30年的研究现状,土壤动物肠道微生物群落组成特征和研究方法,以及其生物和生态学功能(协助宿主消化、提供宿主营养、影响宿主寿命和繁殖、提高宿主免疫能力及参与生态系统碳氮循环),重点介绍了土壤动物肠道微生物的影响因素(肠道结构和生理状态、微生物相互作用、食性、发育阶段、宿主系统发生和环境因子),以期为学界提供土壤动物肠道微生物领域研究的全面认识。
1 土壤动物肠道微生物多样性领域研究现状
为了更加准确表达土壤动物肠道微生物领域近些年所取得的研究成果,本文基于Web of Science核心数据库,以“soil invertebrate”和“gut microbiota”为核心关键词检索文献,文献类型为“Article”和“Review”,并利用CiteSpace软件[17]进行文献去重处理及分析。共检索到1990—2019年土壤动物肠道微生物文献342篇,土壤无脊椎动物肠道微生物领域每年发文量虽少,却呈现增长趋势(图1)。对所有文献进行主题及关键词共现分析发现,16S rRNA、bacteria和diversity是重要连结点,说明16S rRNA高通量测序技术对推动土壤无脊椎动物肠道微生物领域的发展产生了重要影响,而肠道细菌及肠道微生物多样性则是土壤无脊椎动物肠道微生物领域研究主要内容。
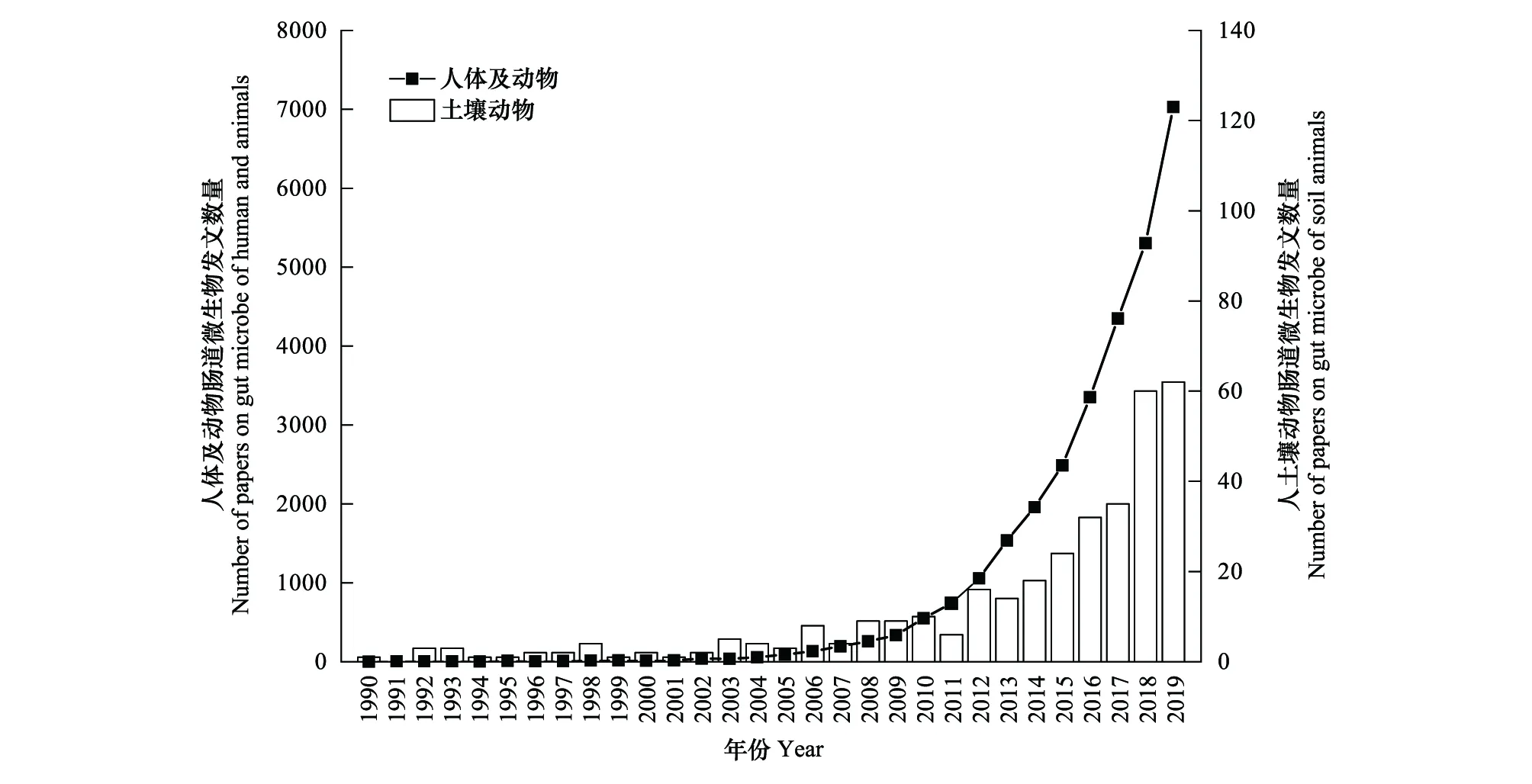
图1 1990—2019年“肠道微生物”领域SCI期刊每年发文量Fig.1 Number of papers published per year in SCI journal in the field of “gut microbe”in 1990—2019
目前土壤无脊椎动物肠道微生物研究现状可总结为以下几点:1)土壤动物肠道微生物研究主要集中在模式动物类群。秀丽隐杆线虫Caenorhabditiselegans、白符跳Folsomiacandida和赤子爱胜蚓Eiseniafetida等模式动物具有生命周期短、繁殖快、科研成本低等优势,是科研人员研究肠道微生物与动物宿主及人体互作关系重要的模式生物[18—19];2)土壤动物肠道微生物多样性及驱动机制[14,20]、共存机制[21]及群落构建[12]的理论研究是微生物生态学领域的前沿;3)土壤动物肠道微生物已由群落多样性组成拓展到功能基因和代谢产物研究,揭示肠道微生物与土壤动物宿主之间相关性及其作用机制[22—23];4)肠道微生物介导土壤动物对人类活动的响应。人类活动造成土地利用方式的改变,施肥、农药、重金属、微塑料及抗生素的环境残留等问题备受世界关注;从肠道微生物角度理解土壤动物对人类活动的响应机制,对土壤生物多样性维持与人类健康及生态系统可持续发展意义重大[24—26]。
2 土壤动物肠道微生物多样性组成与研究方法
2.1 土壤动物肠道主要微生物
细菌因其众多的数量及多样的种类和功能[27],是土壤动物肠道微生物的主要研究对象。土壤动物肠道细菌优势类群以变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)和拟杆菌门(Bacteroidetes)为主,且变形菌门是线虫、螨类、跳虫、蚯蚓等多数土壤动物类群肠道的公有优势细菌门(表1),它们对个体生长、发育十分重要。如Zheng等[20]研究发现,长期施肥下土壤线虫肠道核心细菌属包括变形菌门的肠杆菌属(Enterobacter)和杆菌属(Ancylobacter),它们参与线虫有机食物基质的分解。Holmes等[28]研究发现,南极地区螨虫Alaskozetesantarcticus体内优势细菌纲有Actinobacteria、Flavobacteriia、Sphingobacteriia、Gammaproteobacteria和Betaproteobacteria,且幼螨肠道细菌除Actinobacteria外,其他四个细菌纲的相对丰度均高于成螨,推测这种差别由不同发育阶段螨虫食性和微生境的不同所导致。Xiang等[29]发现不同食性跳虫的肠道细菌群落结构和优势类群组成均有显著差异,但变形菌门和拟杆菌门是所有食物处理跳虫肠道优势细菌门,且其肠道Thermogemmatisporaceae、Brevibacillus和Novosphingobium的相对丰度与跳虫体重呈显著正相关。Liu等[30]以不同类型土壤培养蚯蚓,发现矿质土壤和钾矿石粉蚯蚓肠道优势细菌Proteobacteria和Bacteroidetes的相对丰度显著高于腐殖土壤,而其肠道Firmicutes、Actinobacteria、Chloroflexi和 Candidate division TM7的相对丰度则显著低于腐殖土壤。另外,拟杆菌门被认为是降解环境和动物肠道中高分子有机物的重要细菌。以上研究结果表明,土壤动物肠道菌群结构组成不尽相同,而优势细菌种类却有一定趋同性,其与宿主个体生长、发育、抗逆息息相关。

表1 土壤无脊椎动物肠道优势微生物组成Table 1 Composition of dominant microorganisms in the gut of soil invertebrates
真菌是土壤动物如线虫、螨类和跳虫重要食物资源[31],而土壤动物肠内真菌群落的组成也得到了研究。Gong等[32]采集了山毛榉林地7种甲螨,通过扩增真菌ITS2区域解析了甲螨肠道真菌,结果发现甲螨肠内真菌以子囊菌门(Ascomycota)、担子菌门(Basidiomycota)和接合菌门(Zygomycota)为主,且相比肠道细菌,肠内真菌与宿主碳氮稳定同位素值联系更密切,该研究首次揭示了肠道微生物与土壤捕食者营养生态位的关系。Anslan等[33]研究了森林表栖型跳虫肠道真菌群落,研究表明Ascomycota和Basidiomycota是跳虫肠内优势真菌门,且跳虫肠内真菌群落结构随季节有所差异,表明跳虫在不同季节对真菌食物偏好性可能不同。此外,Sapkota等[21]以草地土壤蚯蚓为研究对象,通过18S rRNA基因测序技术研究了蚯蚓肠内真核生物群落结构,发现除真菌外,Cercozoa、Apicomplexa、Metazoa、Ochrophyta亦是蚯蚓肠内优势真核生物,同时揭示了蚯蚓肠道微生物间存在较多正向交互作用,肠道菌群的共存模式对土壤动物宿主-肠道微生态系统的稳态意义重大。
2.2 土壤动物肠道微生物研究方法
传统土壤动物肠道微生物分析方法建立在分离及纯化培养的基础上[44],通过对菌株进行显微特征观察和生理生化试验来认识微生物多样性及其功能。Ineson等[45]对大型土壤节肢动物Oniscusaseilus和Giomerismarginata食物、肠道及粪便微生物分离并计数,发现从食物、肠道到粪便的细菌数目显著增加;生理生化试验表明,从食物、肠道及粪便分离的菌株生化功能有显著差异。虽然纯培养技术是研究土壤动物肠道微生物多样性及验证其功能的重要手段,但由于绝大多数微生物目前尚不能在实验室培养、以及受到样品及数据通量低等局限性,使得该技术难以成为微生物生态研究的主流方法。而自1985年Lane等[46]使用16S核糖体RNA序列研究细菌进化关系以来,聚合酶链式反应(PCR)、基因克隆文库分析法(GCL)、变性梯度凝胶电泳法(DGGE)、温度梯度凝胶电泳法(TGGE)、限制性酶切片段长度多态性分析法(RFLP)、磷脂脂肪酸法(PLFA)、荧光原位杂交法(FISH)、基因芯片、16S文库测序等分子生物学技术逐渐发展且被广泛应运于微生物群落结构分析[47]。近些年来,以高通量测序技术为基本手段,宏基因组、宏转录组、宏蛋白组、宏代谢组等新兴分子生物学技术快速推动了整个微生物生态学领域研究发展[1]。这些方法突破传统纯化培养方法的限制并较能准确认识自然界中微生物的群落结构、进化、演替及功能等[23,48]。在土壤动物肠道微生物多样性的研究中,Anslan等[49]结合纯培养和高通量测序技术研究土壤表栖型跳虫肠內及其体表真菌群落结构与组成,发现高通量测序技术相比培养技术能够获得更高的真菌多样性。Dirksen等[50]利用高通量测序技术研究自然生境线虫肠道菌群,发现Pseudomonas是优势细菌属,进一步对Pseudomonas细菌进行分离培养并进行抗菌试验,发现该属的三种菌株有抵抗真菌病原体现象。因此,结合新兴分子生物学技术如高通量测序和分离培养技术对研究土壤动物肠道微生物菌群结构及功能验证十分必要。
对于较大体型土壤动物如蚯蚓[37],马陆[41,51],通常先把其肠道组织置于无菌环境中解剖提取,进而研究肠道组织内部微生物。对于中型土壤动物如跳虫和螨类,解剖获得肠道微生物是传统方法,延续至今[24,52—53]。而对小型土壤动物如线虫及部分中型土壤动物而言,它们体型较小,其肠道组织提取困难。近几年一些研究者用洗脱掉体表微生物的虫体来研究其内部肠道菌群[32],而洗脱去除虫体表面微生物方法不尽相同。Zheng等[20]研究线虫肠道菌群先将线虫放入2%的次氯酸钠溶液处理10s,后用无菌磷酸盐缓冲溶液漂洗4次,并将最后一次洗脱液涂于LB平板无菌环境培养24h,未观察到菌落,这表明线虫体表微生物已几乎被完全去除。Gong等[32]研究甲螨肠道菌群先将螨虫转移至1% Tween 20中涡旋2min,然后转移至98%乙醇中旋涡5min,接着转移至1% Tween 20中旋涡2min,再转移至5%漂白剂中旋涡5min,最后在1% Tween 20中涡旋2次后研究其肠道微生物。Hao等[54]先将雪地生活跳虫附肢移除以减少虫体表面微生物的携带,接着用次氯酸钠溶液及无菌水进行洗脱来研究其体内微生物。需注意的是,虽然去除虫体体表微生物为研究土壤动物肠道菌群带来巨大便利,但其同时也存在争议。利用洗脱法研究土壤动物关联的微生物会包含虫体内部其他非肠道部位的微生物,如生殖组织及脑部的胞内细菌Wolbochia[55]。另外,由于这些非肠道的微生物可能大量存在于虫子体内,导致肠道部位的微生物无法被有效扩增,从而掩盖肠道微生物群落结构及组成的真实特征。因此,用该方法进行研究肠道微生物时应根据具体情况考虑引物的选用、测序深度,以及下游数据分析时对非目标序列的去除等细节。
3 土壤动物肠道微生物的生态功能
3.1 协助宿主消化
土壤动物肠道微生物能够协助宿主消化食物基质。芽孢杆菌纲细菌被报道普遍存在于取食凋落物土壤动物如跳虫、白蚁、等足类、多足类及蚯蚓肠道中[4,56],与木质纤维素食物基质的分解联系密切[57]。土壤动物肠道中的肠杆菌科及黄单胞菌科细菌也被报道与有机物质分解有关[58—59]。Yang 等[60]发现EnterobacterasburiaeYT1和Bacillussp.YP1是印度谷螟Plodiainterpunctella幼虫肠道降解高分子聚乙烯重要细菌,该研究首次证明节肢动物肠道细菌对塑料降解的重要性。Agamennone等[23]利用宏基因组鸟枪法测序技术研究了自然生境中白符跳体内微生物组成及功能基因多样性,发现跳虫体内微生物具有极其丰富的联系碳水化合物代谢的功能基因,揭示了肠道微生物对多糖物质分解的重要性。肠道微生物源代谢产物亦是降解大分子物质的关键物质[61],影响着宿主食物消化。
3.2 提供宿主营养
土壤动物肠道微生物能影响宿主的营养利用与生长发育。Borkott等[62]研究发现Xanthomonasmaltophilia和Curtobacteriumsp.是白符跳肠道降解几丁质的优势细菌,用添加四环素几丁质饲喂跳虫后,发现跳虫生物量显著降低,这可能是四环素抑制了肠道细菌活性进而影响跳虫对几丁质的营养吸收。Xiang等[29]研究发现白符跳肠道微生物Thermogemmatisporaceae、Brevibacillus和Novosphingobium的相对丰度与跳虫体重呈正相关。其他无脊椎动物如昆虫纲和甲壳纲的相关研究也证实其肠道微生物能够促进宿主体重的增加[63—64]。此外,部分土壤动物如线虫、跳虫及螨类等类群会主动摄食环境中的微生物[18]。如跳虫偏好取食某些真菌[33],而跳虫肠道内的几丁质降解酶能帮助分解消化真菌的细胞壁[65],这些微生物被消化分解后进一步能提供宿主碳水化合物、脂肪和氨基酸等必要营养物质。Wüst等[39]研究也发现蚯蚓肠道中梭菌科和大肠杆菌科细菌是葡萄糖衍生物的重要贡献者,能为宿主合成所需的营养物质。
3.3 影响宿主寿命和繁殖
土壤动物肠道微生物影响宿主的寿命和繁殖。Houthoofd等[66]研究发现,无菌培养上调了秀丽隐杆线虫抗氧化和抗热应激体细胞维护功能,且无菌条件线虫的发育时间和成体寿命增加了两倍。MacNeil等[67]用三种细菌喂食秀丽隐杆线虫,相比喂食大肠杆菌OP50和大肠杆菌HT115,喂食丛毛单胞菌DA1877显著减少了线虫的繁殖率,同时喂食丛毛单胞菌DA1877线虫的寿命相比其他两种细菌也大大缩短。胞内共生菌Wolbachia也被报道能够调控多数节肢动物的繁殖行为[68],Wolbachia产生的核黄素和生物素代谢物是调控宿主繁殖行为重要物质[69]。此外,肠道微生物也参与调控宿主其他行为,如Akduman等[70]以线虫为研究对象,发现线虫Pristionchuspacificus摄入细菌Novosphingobium能够增加其对秀丽隐杆线虫的捕食能力,且揭示细菌产生的维生素B12是造成捕食行为增加的重要原因。
3.4 提高宿主免疫能力
Dirksen等[50]研究自然生境秀丽隐杆线虫肠道微生物,发现其肠道内假单胞菌MYb11,MYb187和MYb193对真菌生长有抑制作用,提高了线虫对真菌病原体的抵抗与防御。Agamennon等[15]从5种跳虫Folsomiacandida,Folsomiafimetaria,Orchesellacincta,Sinellacurviseta和Tomocerusminor肠道分离得到46种肠道细菌并对其进行抗菌活性研究,发现分离的肠道细菌中,76%微生物能抑制细菌或真菌病原体活性,抑制活性肠道细菌中有51%对细菌和真菌病原菌均有抑制作用。Iatsenko等[71]研究发现秀丽隐杆线虫肠道枯草芽孢杆菌(GS67)增加了线虫对革兰氏阳性病原菌的抗性,且脂肽风霉素是枯草芽孢杆菌产生的主要抑制物质。此外,抗生素和抗菌化合物也是抑制病原菌定植生长的肠道微生物源代谢产物[72]。这些研究揭示了土壤动物肠道微生物间抵抗抑制作用能提高宿主的免疫能力。
与人类肠道类似,土壤动物肠道除共生菌和益生菌外,也存在致病菌。这些菌的多少和之间的互作模式决定了它们整体对宿主个体健康的影响。研究发现,森林病原真菌Heterobasidionannosum被检测存在于自然生境中跳虫的肠道及体表[33],对跳虫生长发育产生不利影响。Garigan等[16]研究发现线虫肠道致病细菌的增殖与累积导致了宿主的死亡,推断这是微生物源致病代谢物质作用的结果。当土壤动物面临众多寄生微生物和致病菌威胁时,能够通过共生菌的屏蔽作用如对病原菌产生高度的定植位置竞争、营养竞争,以及增加宿主免疫竞争力与疾病对抗从而对机体进行有效保护[72],且物种多样的肠道菌群更利于抵抗病原菌入侵。
3.5 参与生态系统碳氮循环
土壤动物参与陆地生态系统碳氮循环过程,离不开宿主肠道微生物。土壤动物取食、消化及排泄行为与土壤有机质分解和矿化过程紧密联系[73]。动物食性影响了肠道菌群结构及组成[29],同时肠道微生物又能够协助宿主对各种复杂食物如真菌、细菌、植物残体及动物体等进行分解消化[4,74],经吸收利用后被排遗于环境,进而影响碳氮循环。另外,研究表明,螨类和跳虫是微生物扩散与传播重要媒介[49,75—76],摄入土壤动物体内微生物会随宿主的活动迁移而被传播扩散,这对环境微生物群落结构起到了间接调控作用,进而影响微生物分解有机质的过程。土壤动物肠道微生物能促进气体排放[77],如蚯蚓反硝化释放的N2O与其肠道Rhizobiales细菌有关[78],Methanobacteriaceae和Methanoregulaceae肠道细菌也被报道与CH4释放有关[38,79]。此外,土壤动物肠道微生物能影响土壤中有害元素的转化,如蚯蚓肠道微生物能够介导土壤中砷的还原[26]。因此,肠道微生物能介导土壤动物对生态系统中元素循环过程。
4 土壤动物肠道微生物多样性的主要影响因素
4.1 肠道结构和生理状态
不同动物物种的肠道或不同区段的肠道微环境在形态和理化性质上存有很大差异,能影响微生物数量及群落结构。相比哺乳动物,土壤无脊椎动物消化道结构相对简单,其肠道微生物种类较少且群落多样性较低[3]。而拥有复杂消化道结构的土壤动物,如白蚁,其肠道微生物则较复杂多样。白蚁后肠存在特定膨大结构,其内包含大量微生物,对木质纤维素的降解发挥着重要作用[80]。另外,不同肠道部位的微生物数量及群落结构有所不同,如Cazemier等[81]以7个目节肢动物为研究对象,发现从前肠、中肠到后肠的微生物数量依次增加,这可归因于宿主对微生物选择作用[82]。Hu等[37]发现,相比于后肠,蚯蚓前肠微生物群落结构与环境微生物群落结构组成更相似,而后肠微生物群落结构则更稳定,也是宿主营养吸收主要部位[83]。此外,动物肠道生理状态影响其内微生物的定居。土壤动物肠道是一个厌氧环境,从而筛选体内微生物的群落结构[40]。Brune等[84]使用微电极方法研究了5种食土白蚁肠道pH特征,发现白蚁中肠到后肠的pH值显著增加,中肠pH值接近中性而后肠呈碱性。由于pH是影响微生物群落结构重要因子[85],因此推测食土白蚁不同肠道部位pH亦会影响其内微生物的定殖。
4.2 微生物相互作用
微生物间的相互作用在微生物群落构建过程中起重要作用。Berg等[12]以秀丽隐杆线虫为研究对象,发现线虫肠道微生物群落构建过程是非随机的,通过物种共存网络分析发现线虫肠道微生物间的负向作用比土壤微生物间的多,揭示物种间竞争是线虫肠道微生物共存的主要现象。另外,Ding等[36]研究长期施肥处理下土壤跳虫Folsomiaquadrioculata体内细菌微生物间相互作用,发现在跳虫体内黄单胞菌科、黄杆菌科和假单胞菌科与其他细菌科的负向相互作用比在土壤中强,表明跳虫体内细菌群落与土壤细菌群落存在不同生物相互作用模式。Sapkota等[21]研究草地土壤中蚯蚓发现,蚯蚓的种类相比于生态型对肠道微生物群落结构影响更大;而对蚯蚓肠道细菌及真核生物进行跨界共存网络分析,发现真核生物和细菌相互作用中,正向作用显著多于负向作用,Cercozoa是与细菌相互作用数目最多的真核生物,而Heteromita是与细菌作用最密切的真核生物。由于生物间相互作用是微生物物种共存和群落构建的重要过程[86],现有的研究更多是揭示相互作用的模式,土壤动物肠道微生物间的相互作用具体机制则需要结合多组学技术与操控实验进一步研究。
4.3 土壤动物食性
动物食性是决定其肠道微生物群落结构和组成的重要因素。Xiang等[29]研究发现,以植物凋落物、细菌和酵母菌混合食物饲喂跳虫时,跳虫肠道微生物多样性最高,而以单一凋落物饲喂跳虫时的肠道细菌群落多样性最低。其他动物如昆虫的研究,也证实了杂食性昆虫肠道微生物多样性显著高于肉食和植食昆虫肠道的微生物多样性[87]。多样的肠道菌群可能与多样复杂食物基质的分解消化密切联系[88]。Kudo等[14]研究不同食性甲虫肠道微生物结构,发现腐食性甲虫肠道中黄单胞菌科是优势细菌类群,而肉食性和植食性甲虫肠道肠杆菌科是优势细菌类群,显示食物类别影响了肠道中某些微生物的富集。Knapp等[89]通过使用不同食物资源饲养蚯蚓Lumbricusrubellus,也发现不同食物处理下蚯蚓肠道菌群结构差异显著。Mikaelyan等[90]利用16S RNA基因测序了18种高等白蚁肠道菌群,发现食性是驱动肠道菌群分化的重要因素,食木高等白蚁相比取食土壤和腐殖质高等白蚁,其肠道内含有更多螺旋体菌门(Spirochaetes)、纤维杆菌门(Fibrobacteres)和TG3门细菌,这对降解木质纤维素食物资源发挥着重要作用[91]。
4.4 土壤动物发育阶段
不同发育阶段土壤动物个体,其生理状态、食性和栖息微环境皆可能影响肠道菌群结构与组成。Dirksen等[50]以线虫为研究对象,发现其肠道微生物群落结构受宿主发育阶段影响。另外,Holmes等[28]研究发现,螨虫Alaskozetesantarcticus末龄幼体与成体体内微生物种类无显著差异,而在微生物的相对丰度上有较大差异,推测这些重叠微生物是由于幼体和成体拥有相似的栖息生境和取食偏好。Shukla等[92]以金龟子科物种Euoniticellusintermedius和Euoniticellustriangulatus为研究对象,发现它们的卵内部共生微生物群落结构与雌性成体肠道微生物群落结构更相似,说明卵内部共生菌主要受遗传因素影响并以垂直传播方式为主[93];而幼体肠道微生物群落结构和成体肠道微生物群落结构有差别,也许是幼体和成体的食性及栖息环境的不同所导致[94]。因此,土壤动物通过垂直和水平传播方式获取其肠道微生物,与其他动物类群规律类似。然而,对于不同发育阶段的土壤动物个体而言,其肠道菌群以垂直和水平方式获取的种类及比例目前仍不清楚。
4.5 宿主系统发生
土壤动物肠道微生物群落结构具有一定的宿主物种稳定性,其受宿主系统发生影响,且与宿主表现出协同进化的特征。Hao等[54]发现两种德跳属雪地生活跳虫体内菌群结构相比另一鳞跳科物种更相似,可能是亲缘相近物种的栖息生境及取食特性较相似所导致。Yun等[87]以18S rRNA基因分析了21目218种昆虫的系统发生,同时利用16S rRNA基因通过454焦磷酸测序手段测试了昆虫肠道微生物群落,发现昆虫系统发生、生存环境、食性及发育阶段均显著影响了肠道厌氧菌相对丰度。Gong等[32]通过研究7种甲螨系统发生与其体内菌群之间关系,发现相比于甲螨肠道真菌,其肠道细菌群落与甲螨的系统发生更相关,表明甲螨肠道微生物群落结构受其系统发育的影响,且肠道细菌与甲螨宿主可能存在更密切的协同进化关系。Kudo等[14]研究了5科46种甲虫肠道微生物,通过分析宿主和肠道微生物群落的系统发育,发现亲缘关系密切的甲虫物种,其肠道肠杆菌科群落系统发育也相似,揭示出肠杆菌科细菌与甲虫宿主存在长期进化关系。上述研究显示,亲缘关系密切土壤动物的肠道菌群结构相似,微生物类群能否存在于宿主肠道与宿主系统发育有关,且肠道菌群与宿主存在协同进化关系。
4.6 环境非生物因子
土壤动物肠道菌群影响因素除宿主因素外,环境因子也是重要影响因素,如温度、时间、空间、微塑料、抗生素等。温度引起机体肠道微生物的变化是温度变化影响宿主适应能力的一种重要机制[95]。有关两栖动物、鱼类的研究表明,外界温度改变了肠道微生物群落组成,并介导了外界温度与宿主消化能力之间关系[96—97]。Berg等[12]研究温度对土壤线虫肠道微生物群落有影响,发现线虫肠道Sphingobacterium细菌属的相对丰度随着温度升高而增加,Agrobacterium相对丰度则随着温度升高而降低。Bing等[98]研究发现,夏季果蝇肠道细菌多样性显著高于秋季。Anslan等[33]发现跳虫肠道真菌群落丰富度和组成随季节呈现一定动态变化,推测跳虫肠道微生物随季节的变化与自身取食偏好特征有关。Hao等[54]研究发现,冬季雪地生活跳虫肠道微生物如Cyanobacteria是宿主重要食物资源,推测其能够为低温环境中生活的跳虫提供机体能量,对维持跳虫生命活动十分重要[99—100]。然而,温度对土壤无脊椎动物冬季和生长季间肠道菌群结构的影响目前尚不清楚,肠道微生物介导土壤动物适应低温的机理是未来必要的研究。
此外,局域尺度上空间因子及生境类型均影响土壤动物肠道菌群结构。Kudo等[14]对不同栖息生境甲虫肠道微生物研究发现,所有营养类型甲虫的肠道细菌群落结构在林地和草地间皆是有差异的。Zhu等[25]研究了不同土地利用方式下跳虫Orchesellidessinensis肠道菌群,发现公园用地、公路用地及农业耕地之间的跳虫肠道细菌alpha和beta多样性差异显著,公园用地跳虫肠道细菌alpha多样性最高,而耕地alpha多样性最低。此外,Bahrndorff等[58]采集土壤类型和植被相似的四个地点跳虫Orchesellacincta,研究发现不同地点跳虫种群肠道微生物群落组成有一定差异。然而,在区域甚至是全球尺度上土壤动物肠道微生物群落格局目前尚不清楚。
其他研究则表明微塑料、抗生素及农业施肥等改变了土壤动物肠道微生物群落多样性和结构组成[24,40,101]。如Zhu等[24]以白符跳为研究对象,结果发现:比较对照组,微塑料处理显著增加了白符跳肠道细菌Alpha多样性,且显著增加了优势细菌厚壁菌门的相对丰度但降低了拟杆菌门的相对丰度。另外,微塑料处理显著抑制了跳虫繁殖行为;相比对照组,微塑料处理的跳虫拥有更高的δ15N 和δ13C,微塑料也许通过影响跳虫肠道菌群而影响了宿主对元素的同化及其生长和繁殖。Zhu等[101]研究发现,抗生素降低了土壤跳虫肠道微生物多样性,显著增加了优势细菌放线菌门和拟杆菌门的相对丰度,且显著降低了厚壁菌门相对丰度,抗生素能通过改变跳虫肠道微生物群落结构进而影响跳虫体内抗生素抗性基因的富集。此外,农业施肥亦改变土壤动物肠道微生物群落结构及组成,研究表明,相比空白处理,添加无机肥处理降低了土壤线虫肠道微生物多样性,但却增加了变形菌门的相对丰度[34]。另有研究表明,与对照相比,无机肥对土壤跳虫肠道微生物群落结构有显著影响,而有机肥对土壤跳虫肠道微生物群落结构无显著影响,且变形菌门对施肥处理的响应最为敏感[36]。土壤动物肠道微生物多样性及群落组成对不同环境污染物及施肥管理方式的响应模式有所不同,宿主能够通过调控自身肠道菌群结构组成进而帮助宿主适应环境。
5 结语与展望
土壤动物种类众多,食性复杂,其肠道微生物也丰富多样。土壤动物肠道微生物与宿主互相作用、协同进化,影响土壤动物自身的生长发育及其生态功能的实现。土壤动物肠道微生物的种属阶元数量、类别以及每种微生物含量存在很大差异,而宿主和环境因素则共同影响土壤动物肠道菌群组成与多样性。土壤动物肠道微生物领域研究已取得显著成果,且日渐得到重视。在研究技术上,目前土壤动物肠道微生物领域研究以传统分离培养和扩增子测序技术为主,多数研究主要揭示肠道微生物多样性、群落结构组成,以及肠道微生物与宿主食性、生长、繁殖和寿命等之间的相关性。然而,对土壤动物肠道微生物功能基因及其代谢产物在这些过程中的具体作用机制知之甚少,未来应突破技术瓶颈,结合多组学技术如宏基因组、宏转录组及代谢组等手段,以进一步阐明土壤动物肠道微生物基因及代谢产物与宿主分子及生理的互作机制。在研究对象上,土壤动物肠道微生物研究主要聚焦在土壤模式生物,应加强对野外土壤动物种群和群落肠道微生物多样性的研究。另外,目前对土壤动物肠道微生物研究主要集中在单一微生物类群,如多数研究重在调查肠道细菌群落多样性,未来应关注土壤动物肠道多界微生物如肠道细菌,真菌甚至原生生物等研究,以全面解析土壤动物肠道微生物多样性和跨界物种共存互作模式及其群落构建机制。在研究尺度上,现有研究主要通过控制实验揭示宿主或环境因子对土壤动物肠道微生物多样性及群落结构组成的影响,未来应关注自然生境中大尺度范围土壤动物及其肠道微生物的调查研究,揭示两者在局域和区域尺度上的多样性格局,以探究动物-微生物互作机制对土壤生物地理分布格局与生态系统功能的影响。此外,土壤动物肠道菌群的季节动态研究将能增进我们对土壤生物适应极端环境与反映气候变迁的理解。
致谢:东北师范大学环境学院管平婷博士帮助写作,特此致谢。
