重金属污染土壤植物修复中细胞分裂素的作用与机制
2022-05-20方治国谢俊婷都韶婷
方治国,杨 青,谢俊婷,都韶婷
1 浙江工商大学环境科学与工程学院,杭州 310016 2 浙江树人大学交叉科学研究院,杭州 310015
土壤是生态系统中物质与能量交换的重要场所,是人类赖以生存和发展的根本基础,土壤环境中重金属污染已受到各国科研人员的高度关注。重金属具有高毒性、持久性、隐蔽性和难降解性,进入环境后不断迁移,通过生物链富集作用直接或间接地危害人体健康[1]。目前,重金属污染土壤的修复技术主要有物理修复、化学修复和生物修复等,在实践过程中也逐渐开展了多种修复方式的联合应用。植物修复因投资成本低、二次污染易控制、环境扰动少等优点受到研究学者的青睐[2]。强化植物修复效果的技术手段主要有水肥调控、菌剂接入、基因工程、螯合钝化剂和土壤动物添加等,这些技术措施改变了植物的生长状态、生化特征和抗性能力等,进而会影响植物修复的整体效果[3]。
植物内源激素是植物体内存在调节生长发育的低浓度小分子有机物,共分为七大类:细胞分裂素(cytokinin,CTK)、生长素 (auxin,IAA)、乙烯 (ethylene)、脱落酸 (abscisic acid,ABA)、赤霉素 (gibberellin,GA)、油菜素甾醇 (brassinosteroids,BR)及其它植物激素 (水杨酸、茉莉酸等)[4]。随着科研人员对植物内源激素系统性研究的不断深入,发现其在重金属污染土壤植物修复中具有较强的应用潜力。近年来,IAA、ABA、GA等内源激素在植物修复中的应用及作用机理已开展了大量研究,但与CTK相关的研究内容与作用机制报道较少[5-7]。论文以CTK在调控植物生长发育及强化重金属污染土壤植物修复中所取得的研究成果为基础,重点整合了CTK对植物机体生理生化过程中的调控机理,系统论述了CTK对强化重金属污染土壤植物修复中的作用与机制,并展望了CTK在植物修复中的研究方向。
1 细胞分裂素的种类与作用机制
1.1 细胞分裂素的概念和种类
1957年Skoog等[8]通过对鲱鱼精子 DNA 样本进行高压灭菌纯化出一种名为激动素 (kinetin,KT)的物质,用该物质对烟草 (Tobacco)愈伤组织进行体外培育发现可促进愈伤组织细胞分裂;1963年,Letham等[9]从玉米种子中提取出类似KT的物质,并发现该物质是一种天然存在高活性的细胞分裂刺激剂,最终命名为玉米素 (zeatin,ZT)。随着科学研究不断深入,Skoog等提出将所有与KT有相同作用的可促进植物细胞分裂、诱导叶芽形成、调控生长发育的化合物,统称为细胞分裂素 (cytokinin,CTK)[10]。
目前,CTK主要分为自然存在和人工合成两大类,天然细胞分裂素是在N6位具有类异戊二烯或芳香族侧链的腺嘌呤衍生物[11],高等植物体内最丰富的类异戊二烯类CTK主要包括:异戊烯腺嘌呤 (isopentenyl adenine,iP)、顺式玉米素 (cis-zeatin,cZ)、反式玉米素 (trans-zeatin,tZ)和二氢玉米素 (dihydrozeatin,DHZ)[10],而天然芳香族类CTK在植物体内含量较低,其中活性最高的CTK主要是6-苄基腺嘌呤 (6-Benzylaminopurine,6-BA)以及它的相关衍生物[12]。常见的人工合成CTK主要有KT、6-BA、ZT、四氢吡喃苄基腺嘌呤(tetrahydropyranyl benzyladenine,PBA)等[13],针对CTK对植物生长发育的作用机制与效果,人工合成的CTK外源物质在农业以及园艺方面有着广泛的应用。
1.2 细胞分裂素在植物生长过程中的作用
CTK对于植物生长发育的调控作用主要包括:促进植物细胞分裂与分化、调节叶芽和根部生长、减轻顶端优势、延缓叶片衰老、促进叶绿体发育等。研究表明,植物七大类内源激素的主要作用机制与影响效果虽各不相同,但对于植物生理过程的调控往往涉及多种激素的共同参与[14],其中CTK便与其他内源激素存在信号串扰与相互作用。例如:乙烯在植物体内调控衰老过程,诱导果实成熟,而Zdarska等[15]研究发现植物根部的乙烯通过信号传递调节CTK多步磷酸化正调节剂arabidopsisresponseregulator10 (ARR10)表达,激活了植物根部合成CTK,进而促进根尖分生组织 (root apical meristem,RAM)细胞分裂与发育,这表明乙烯与CTK彼此间的相互作用,调节着根部萌发与生长;ABA主要是一种引起植物幼芽休眠、叶片脱落、生长延缓等生理作用的内源激素,而在干旱条件下,CTK会与ABA产生拮抗作用,通过诱导叶片气孔闭合,提高植物对干旱胁迫的耐受性[16];GA在高羊茅 (Tallfescue)体内通过刺激蘖芽负调节基因teosintebranched1 (FaTB1)表达,抑制分蘖芽生长发育[17],而Zhuang等[18]通过外源施加10 μmol/L的CTK,发现蘖芽抑制作用得到缓解,分蘖芽长度显著提升。以上结果皆表明CTK与内源激素间会通过信号传导、协同与拮抗等作用共同调控植物生理活动[19]。
2 重金属污染土壤植物修复中细胞分裂素的作用与机制
2.1 增强光合作用,延缓叶片衰老,提升植物抗性能力
植物体内叶绿体是CTK 作用的主要靶标之一,内源CTK激活叶绿体基因转录,外源CTK可提高叶绿体合成蛋白质水平,促进叶绿素合成酶的形成,增强植物光合作用[20]。Lu等[21]发现在拟南芥 (Arabidopsisthaliana)内源CTK应答基因cytokinin-responsivegata1 (CGA1)信号传导的下游,硝酸盐诱导的碳代谢基因GATA,nitrate-induciblecarbon-metabolism-involved(GNC)受到刺激并转录,CGA1和GNC作为叶绿体发生的两个主要转录调节因子,彼此相互作用促进了叶绿体分裂与生长;同时Criado等[22]发现外源6-BA刺激小麦 (TriticumaestivumL.)叶绿体基质膨胀,促进叶绿素含量增加,抑制蛋白质在体内降解,显著提高了叶片蛋白质和淀粉含量。内源CTK调节植物光合作用,而外源根部施加、叶面喷施CTK皆会促进体内叶绿素含量显著上升,降低叶绿体损伤,增强植物光合作用能力[23]。
在叶片衰老初期,衰老启动子senescence-associatedgene12 (SAG12)会被激活,促进异戊烯基转移酶基因isopentenyltransferasegene(IPT)合成CTK[24],生成的CTK会促进体内与脂肪酸代谢相关的蛋白质,如:脂氧合酶、酰基载体等蛋白表达,清除叶面细胞的活性氧自由基,同时CTK还会促进氮转运基因Ntransport-regulatedgene1.1 (TaNRTs1.1,TaNRT1.3,TaNRT1.4)表达,提高硝酸还原酶活性,减轻细胞膜氧化损伤[25—26]。同时,Liu等[27]采用0.01、0.1、1、10μmol/L的6-BA进行水稻 (OryzasativaL.)叶片培育,发现叶片衰老速度与浓度呈负相关,且6-BA下调了氨基酸代谢基因Osl20和Osh36、应激反应基因Osl43等SAGs代表性基因的转录水平,抑制了衰老基因表达,同时植物体内6-BA与茉莉酸调节叶绿素降解基因chlorophyll-degradation-relatedgenes(CDRGs)表达,彼此间通过拮抗作用,调控叶片衰老过程[28]。
因重金属胁迫致使植物体内产生大量活性氧自由基(reactive oxygen species,ROS),引起植物代谢产物紊乱、脂质过氧化、酶活性降低、细胞膜损伤、细胞失活甚至死亡[29],此时植物激活抗氧化系统,通过释放抗氧化物质抵御ROS损害。其中,Zhou等[30]在镉和锌复合污染条件下,对海滨锦葵 (Kosteletzkyapentacarpos)施加10 μM的t-ZR,显著降低了丙二醛 (malondialdehyde,MDA)含量;Wang等[31]在砷 (As)污染土中种植0、0.1、0.5、1.5mg/L的KT浸泡后的玉米 (Maize)种子,收获后测定发现超氧化物歧化酶 (superoxide dismutase,SOD)活性在0.5mg/L的处理组下提高50%,其后逐渐下降;在1.5mg/L处理时,体内过氧化物酶 (peroxidase,POD)、过氧化氢酶 (ctalase,CAT)活性提高约50%,体内As含量提高3倍。以上结果表明外源CTK能 提高SOD、POD、CAT 等抗氧化酶活性,增加抗坏血酸、脯氨酸、可溶性蛋白等非酶抗氧化剂含量,同时降低MDA、H2O2、超氧阴离子自由基 (O2-)等膜脂质过氧化指标,减轻ROS积累与损伤,提高重金属胁迫抗性,这一作用保障了植物在重金属胁迫下良好生长状态,为重金属污染土壤植物修复奠定了坚实基础[32—33]。
2.2 调控根茎叶发育,增加植物生物量,强化植物富集效果
植物内源CTK主要在根部产生,其会抑制根部生长因子表达,并与IAA产生拮抗作用,负调控根部生长。一方面,当CTK与IAA比值在根部低时才会促进细胞增殖,根部器官主是由RAM分化而来[34],在RAM发育初期,首先CTK信号表达增强[35],在IAA接收信号后,会直接抑制A类CTK响应调节因子ARR7和ARR15表达,促进CTK负调节因子arabidopsishistidinephosphotransferprotein6 (AHP6)转录上升,其后CTK含量逐渐下降,与此同时IAA负调节蛋白shortpypocotyl2 (SHY2)活性被抑制,IAA外输载体蛋白PIN-formed(PIN)表达上升,IAA合成逐渐上升[36]。其后,CTK又会通过B型ARR刺激SHY2信号表达,抑制PIN活性,阻碍IAA合成,因此,CTK与IAA二者主要通过信号传递以及拮抗作用,协调根部发育[37]。另一方面,内源CTK还会下调根部促生长因子stuntedplant1 (STP1)表达,Werner等[38]发现在拟南芥细胞分裂素氧化酶/脱氢酶cytokininoxidasesdehydrogenase(CKX)过表达的突变体中,CTK含量的降低导致STP1蛋白合成上升,根部生长显著优于野生型,因此CTK对于植物根部生长发育往往起到负调控作用。
植物内源CTK通过与茎尖分生组织 (shoot apical meristem,SAM)相关基因的相互作用,调控植物SAM区域活性与大小。首先,在SAM中心与分生组织稳态和叶片发育有关的I类家族基因knotted1-likehomeobox(KNOX)转录因子会直接激活IPT基因[39],促进区域内CTK合成,促进SAM细胞分裂,Kurakawa等[40]发现在水稻SAM中当CTK激活酶基因LONELYGUY(LOG)表达后,会促进无活性的CTK转化为具有生物活性的游离碱形式,致使CTK在特定发育域中被激活,进而直接提高SAM活性。其次,同源结构域转录因子WUSCHEL(WUS)是定义干细胞生态位的关键调节剂,并且WUS是诱导SAM增殖分化的直接媒介[41],CTK通过B型ARR激活SAM中WUS的转录,刺激分生组织启动与发育[42]。同时,Chatfield等[43]发现在拟南芥ARR7和ARR15过表达突变体内CTK含量显著下降,与野生型相比SAM细胞活性、茎部发育都受到显著抑制,由此表明CTK在茎尖分生组织局部激活、分裂分化、活性提高等方面发挥着关键作用。
植物富集是利用植物根系从土壤中吸收重金属并将其转移到可收获的地上部分,并且可从植物的可收获部分中回收金属,但如今许多超级累植物如印度荠菜 (Brassicajuncea)、拟南芥、东南景天 (Sedumalfrediihance)、天蓝遏兰菜 (ThlaspicaerulescensL.)等自身生物量较小[44],且多数植物在生长过程中存在一定程度的顶端优势,抑制了侧枝发育[45],这更加减小了重金属蓄积空间,导致植物修复效果难以达到预期要求。外源CTK的应用可减轻植物顶端优势,促进叶片和侧芽细胞的分裂,增加植物生物量,扩大重金属蓄积体,增强重金属富集效果。Luo等[46]通过对桉树 (Eucalyptusglobulus)进行叶片喷施20 mg/kg的商业产品CTK,发现实验组整株生物量与对照相比增长了39.1%,同时Thomas[47]等在铜 (Cu)胁迫下,发现在CTK过表达烟草中生物量显著增加,成熟叶片中Cu富集量提高8倍。其实在植物茎叶部位,当CTK与IAA比值高时,会促进顶端以下腋芽枝条发育[48],此时外源CTK会转化为内源t-ZR、IP,而内源CTK的B型ARR信号因子会直接激活CTK受体蛋白组氨酸蛋白激酶arabidopsishistidinekinase4 (AHK4)、CKX4/5、WUS等基因表达,进而促进叶片幼芽细胞分裂增殖[49]。植物体内CTK对于根茎叶发育起到的基础调控作用,为外源CTK应用增加植物生物量提供理论支持,因此CTK能够通过扩大重金属植物蓄积体进而提高重金属富集量[49]。
2.3 增强转运蛋白表达,提高叶面蒸腾作用,促进重金属吸收转运
重金属在植物体内的转运过程直接影响着根部对重金属的吸收程度,转运蛋白是植物体内一类膜蛋白,可实现跨膜转运养分、代谢产物、有毒污染物等生物分子,转运蛋白对植物营养吸收、生长发育、体内稳态和胁迫反应至关重要[50]。CTK通过影响部分转运蛋白的表达,调控植物对于重金属的吸收与转运。研究表明,拟南芥对砷 (As)的吸收量与体内磷酸盐转运蛋白phosphatetransporter1;1 (PHT1;1)的表达密切相关[51],Castrillo等[51—52]在As胁迫下通过对拟南芥进行qRT-PCR分析发现,当体内CKX1基因过表达时,体内PHT1;1表达被抑制了2.7倍,且体内As含量与对照组相比显著下降;Jiang等[53]发现拟南芥功能缺失突变体ipt 1 3 5 7体内硒 (Se)含量降低47.8%,而受体缺失突变体AHK2和AHK3 体内Se含量增加35.4%、24.1%,由于亚硒酸盐被植物中的磷酸盐 (Pi)转运蛋白吸收[54],通过对比分析发现ipt 1 3 5 7体内PHT1;1、PHT1;8转录分别降低了57.4%、62.7%,而突变体AHK2、AHK3 体内PHT1;1转录提高1.22、1.11倍,PHT1;8转录增加0.86、0.76倍,PHT1;9增加0.69和0.50倍,这些结果表明内源性CTK可通过促进转运蛋白的表达,促进植物对污染物的吸收和积累。其实,在植物机体内也存在着与重金属排毒有关的转运蛋白,这些转运家族主要包括:ATP结合盒式蛋白ATP-bindingcassette(ABC)、金属耐受蛋白metaltoleranceprotein(MTP)、多药及有毒化合物外排蛋白multidrugandtoxinextrusion(MATE)、锌铁调控转运蛋白ZRT and IRT-likeprotein(ZIP)等[55-57],能够调控根部对重金属的吸收,协调机体内部重金属的转运,维持着胞内胞外毒物的平衡。目前,CTK与各类转运蛋白相关性研究报道较少,其作用机制仍需进一步探索和研究。
植物叶面气孔影响着植物蒸腾作用,外源CTK可通过调控叶表皮气孔细胞活动,增强蒸腾作用[58],促进重金属吸收。一方面,CTK与气孔发育关键转录因子speechless(SPCH)相互作用,调节气孔发育的起始,同时CTK通过A型ARR16、ARR17信号调控气孔谱系细胞的分裂,增加气孔数量[59];另一方面,植物自身通过促进ZjPYR1、SmGRP1等基因的表达,增强气孔细胞对ABA的敏感性,抑制叶片气孔开放[60-62],而CTK通过B型ARR1,11,12等抑制ABA正调节因子蔗糖非发酵-1相关蛋白激酶基因sucrosenon-fermenting-1-relatedproteinkinase2 (SnPK2)活性,与ABA产生拮抗作用,进而促进气孔开放[63],同时Marchadier等[64]通过对拟南芥外源施加6-BA,发现降低了气孔细胞对于ABA的敏感程度,延缓气孔闭合,提高蒸腾作用。研究表明,植物蒸腾作用产生的向上作用力,影响着根部对可溶性污染物的吸收程度,且蒸腾强度与植物吸收呈正相关[65—66],这意味着CTK能够通过正调节植物蒸腾作用,促进植物对于重金属的吸收。然而,Arnaud等[67]对拟南芥野生型和突变体AHK3外源施加0.01—10μmol/L的t-Z,发现野生型随着浓度的上升,孔径逐渐减小,而AHK3缺失突变体孔径没有显著变化,这表明外源t-Z通过与AHK3作用诱导气孔关闭,抑制了蒸腾作用。因此,内源CTK调控植物气孔细胞分裂,提高气孔密度,同时促进气孔开放,增强了蒸腾作用,促进了植物对于重金属的吸收。然而,外源CTK的应用所产生的作用,则与种类、浓度、植物特性等有较大关系[68]。同时,目前针对CTK与蒸腾作用以及植物修复之间的相关性研究还较少,彼此间的相互影响与关联机制仍需进一步研究。
2.4 参与解毒过程,降低重金属毒性,调控重金属体内转化
金属离子进入植物体内,会改变植物细胞现有的动力学和氧化还原系统,并对植物细胞膜、叶绿体、细胞核等产生毒性和损伤[69],而CTK信号会被激活并且参与植物体内自身解毒过程,植物自身会释放谷胱甘肽 (glutathione,GSH)、植物螯合素 (phytochelatins,PCs)、半胱氨酸 (cysteine,Cys)等非蛋白巯基,与重金属发生络合反应,将毒性更低的络合物储存在液泡中,降低重金属离子的迁移[70]。Mohan等[52]发现,在As胁迫下,通过对比研究CKX1基因突变体和野生型拟南芥体内PCs合成基因表达情况,发现突变体内部CTK含量下降,同时phytochelatinsynthase1 (PCS1)、glutathionesynthetase2 (GSH2)、γ-glutamylcysteinesynthetase(γ-ECS)等PCs合成基因的表达下降。Singh等[71]在Cd胁迫下对茄子 (SolanummelongenaL.)幼苗喷施10μM的KT,发现KT处理组与对照相比,体内非蛋白巯基含量提高了26%,这说明外源施加CTK能促进体内非蛋白硫醇的合成。Cassina等[72]在镍 (Ni)胁迫下,通过对超富集植物庭荠 (Alyssummurale)进行叶片和根部施加CTK,测定发现CTK并未使体内Ni积累量及硫醇浓度发生显著增加。这意味着CTK调控植物体内非蛋白巯基基因表达,调控解毒物质合成,参与植物体内重金属的解毒机制,但作用效果的强弱与浓度有一定关系[73]。
植物体内金属硫蛋白是富含半胱氨酸的金属结合蛋白,维持金属稳态和应激反应,也是清除植物体内活性氧自由基的重要物质,在植物遭受重金属胁迫时可减轻毒性[74]。Yuan等[75]通过外源施加6-BA、t-Z和KT影响了水稻根部初期发育时金属硫蛋白metallothionein2b(OsMT2b)基因的表达;Thomas等[47]在Cu胁迫下以IPT转基因与非转基因烟草为实验对象,对比分析了两者体内金属硫蛋白metallothionein-likegene2 (MT-L2)基因表达水平,发现对照组烟草体内MT-L2 mRNAs转录水平显著下降,而实验组中由于IPT的过表达促进了CTK生物合成,在转基因烟草体内检测到MT-L2转录水平上升,同时在成熟叶片中Cu积累量增长了8倍。由此可见,内源CTK调控体内硫蛋白基因表达,而外源CTK施用则会影响体内金属硫蛋白含量,参与重金属解毒过程,提高植物对于重金属的积累。目前,有关CTK与多种金属硫蛋白家族相关基因如:AsMT、PoMT、HcMT、SpMT2等的作用机制研究还不够充分深入[76],仍需要进一步加强。
3 总结与展望
论文综述了植物内源激素CTK在调控植物生长发育及强化重金属污染土壤植物修复中的作用,明确了在植物修复中外源CTK能够有效促进植物生长、调节光合作用、促进重金属转运、强化抗性系统等 (图1)。总体而言,低浓度CTK的应用能促进植物生长发育并强化植物修复效果,而高浓度CTK会对植物生长产生不利的影响。CTK作为植物外源激素已在农作物生产中广泛应用,它可显著提高农作物抗盐、抗旱等抗胁迫能力,且人工合成CTK具有价格低廉、种类广泛、应用便捷等特点,因此在重金属污染土壤植物修复中具有良好的应用前景。目前,CTK在植物修复中的应用还存在不足之处,将来可以从以下方面开展深入研究:
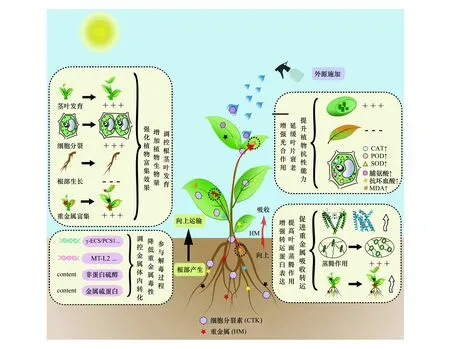
图1 细胞分裂素在重金属污染土壤植物修复过程中的作用机制Fig.1 The mechanism of cytokinin in the process of heavy metal phytoremediation
(1)完善CTK对重金属污染土壤植物修复的作用机制。目前多数研究侧重于CTK对植物生长过程的调控作用,忽略了CTK在植物修复过程中作用机理的研究。在植物修复中,植物挥发机理是通过将汞、砷、硒等重金属吸收至体内再经内部转化释放到大气中,内源CTK对体内金属离子转化的调控机制及外源CTK的应用是否会影响植物挥发的作用过程,目前都不得而知;在植物根际降解机制中,内源CTK对根系分泌物柠檬酸、苹果酸、草酸等产生是否具有调控作用,外源CTK的施加是否会影响植物根系对重金属的降解效果及相关的作用机制,目前都还缺乏确切的解释。因此,应加强CTK对植物修复机理影响作用的研究,为CTK在重金属污染土壤植物修复中的应用提供更为完整的理论依据。
(2)重点关注CTK对土壤结构、性质、酶活、微生物群落等的影响作用,总体把控CTK对污染土壤植物修复系统产生的效应。土壤中有机质、氮磷钾含量、pH等决定着植物营养元素的供给情况;土壤酶活性则是反映土壤质量和肥力的重要生物指标;而植物根系与土壤微生物构成的联合修复体系影响着植物对重金属的吸收积累。但目前的研究重点仍是CTK对植物地上部产生的影响作用,忽略了CTK对于土壤根系及微生物的研究。因此,应深入研究CTK对土壤生态系统的影响机制,探索CTK对于植物-土壤联合系统的作用。
(3)拓展CTK联合超级累植物在实际修复工程中的应用,明确CTK对不同植物修复的强化效果。一方面,工程项目中实施条件复杂,难以控制,这将导致CTK在实际应用时的降解速度、转化程度、强化效果的等均有改变。另一方面,采用超级累植物作为蓄积体进行修复应用较为广泛,但目前对超级累植物联合CTK进行修复的研究较少,且不同植物适用的CTK互不相同。因此,应将CTK联合植物修复技术应用于实际工程项目中,根据应用效果不断完善适应植物修复的外源CTK种类、施加浓度以及施加方式等因子。
(4)明确重金属胁迫下外源CTK的应用对内源激素信号系统的作用机制。内源激素对于植物生长发育具有不可替代的作用,彼此间可通过代谢转化、协同拮抗、彼此调控等作用调节激素合成及植物生理活动,在诱导植物对生物及非生物胁迫的耐受性中发挥着重要作用。因此,明确重金属胁迫下CTK与各激素间的因子响应、基因转录、信号传递等过程,对深层次掌握CTK在植物修复中的作用有着重要意义。
