利用花培与辐照诱变培育粳型两系不育系江79S
2022-05-20谭瑗瑗富昊伟张渭章吴三玲舒庆尧
谭瑗瑗 汪 庆 富昊伟 张渭章 吴三玲 舒庆尧,*
(1 浙江大学作物科学研究所/国家水稻生物重点实验室,浙江 杭州 310058;2 无锡哈勃生物种业技术研究院有限公司, 江苏 无锡 214100;3 嘉兴市农业科学研究院,浙江 嘉兴 314024;4 浙江之豇种业有限责任公司, 浙江 杭州 311107;5 浙江大学农生环测试平台,浙江 杭州 310058)
杂交水稻的大面积应用极大地推动了我国及印度、越南、缅甸、印度尼西亚等亚洲国家的水稻生产[1]。近十多年来,涌现了一批以粳稻不育系与籼稻广亲和恢复系配制的粳籼亚种间杂交稻,如宁波市农业科学院、中国水稻研究所、浙江省农业科学院等单位先后育成了春江16A[2]、浙粳7A[3]、甬粳15A[4]等系列粳型不育系,配制出春优84[5]、浙粳优6153[3]、甬优1540[4]等一批强优势品种并在生产上应用。李铮友于在1973年实现了粳型杂交稻三系配套,开创了我国杂交粳稻的研究,并选育出“滇型”系列不育系[6]。国内科研单位也于上世纪80年代开始了两系杂交粳稻育种和两系粳型不育系的研究[7]。
与三系法相比,两系法杂交水稻具有配组自由、育种程序简单、育种周期短、能克服三系不育细胞质负效应等优点[8]。两系法水稻研究始于石明松在1973年发现的光敏不育水稻农垦58S[9],之后国内开始大规模鉴定和培育光温敏不育水稻材料,育成了如衍生自农垦58S的培矮64S[10]、株1S[11]和安农S-1[12]等两系不育系。上世纪90年代中期,我国实现了基于两系杂交水稻的生产应用[13]。目前审定的两系水稻品种主要来自Y58S、培矮64S、株1S等籼型不育系[14]。
在培育出第一个粳型两系不育系农垦58S后,辽宁省水稻研究所等陆续育成了108S、7001S、C407S、L62S等粳型不育系[7]。与籼稻两系不育系相比,粳型两系不育系数量较少,多数未能在生产上大面积应用,且存在遗传基础较狭窄、优质不育系少、制种产量低等问题[15]。因此,选育新的优良粳稻不育系已成为推动水稻杂种优势利用和杂交粳稻品种培育的当务之急。
为促进两系法杂交稻的培育和利用,本研究综合运用花药培养和诱发突变技术育成一个早熟、优质、抗性好的两系粳稻不育系江79S,以江79S为母本,与广亲和恢复系江恢1501杂交,育成一个杂交水稻新品种江两优7901,以期本研究为综合运用花培和辐照诱变技术培育新两系不育系材料提供研究思路。
1 材料与方法
1.1 选育方法
江79S来自杂交组合培矮64S/粳稻H179,由浙江大学作物科学研究所先进育种实验室通过花培与诱变育成,表1为其培育流程。水稻花药培养方法参照文献[16]。诱变处理经中科院近代物理研究所兰州重离子研究装置(Heavy Ion Research Facility Lanzhou, HIRFL)完成,S79干种子用300 MeV·u-1的12C6+离子束辐射处理,剂量率为0.5 Gy·min-1,辐射剂量为80 Gy。
1.2 育性转换特性鉴定
1.2.1 田间育性转换鉴定 2018年在浙江嘉兴(120°43′E、30°50′N)进行,从5月15日开始每隔10 d播种,至7月5日结束,单本移栽,开花期田间观察花粉育性并镜检,确定育性转换特性。
1.2.2 人工气候室育性鉴定 根据《NY/T 1215-2006水稻光、温敏核不育系育性鉴定规程》[17],在中国水稻所人工气候箱设置不同光温条件(表2),对江79S进行育性测试,观察花粉育性和自交套袋结实率。
1.3 稻瘟病抗性与稻米品质测定
1.3.1 稻瘟病抗性鉴定 由浙江省种子管理总站统一委托浙江省农业科学院植物保护与微生物研究所鉴定江79S苗期稻瘟病抗性。2018年检测单位对供试材料浸种、催芽,每份材料穴播10~15粒,水稻秧苗3~4叶期时接种。将接种后的秧苗置于25~28℃的恒温室内,遮光保湿24 h。去除遮光条件后,定时喷雾保湿。试验设2次重复。苗叶瘟在感病对照达7级或以上时进行调查,每个品种取发病最重的3片叶的结果作为抗性评价的依据,苗叶瘟调查分级标准参照《NY/T 2646-2014 水稻品种试验稻瘟病抗性鉴定与评价技术规程》[18]。
1.3.2 稻米品质测定 2018年春在海南收获成熟江79S种子,晒干后取500 g用于稻米品质检测,检测单位为中国水稻研究所-农业部(现农业农村部)稻米及制品抽量监督检验测试中心。依据《NY/T 83-2017米质测定方法》[19]检测糙米率、垩白度、透明度、碱消值、精米率、垩白粒率、国家标准《GB/T 21719-2008稻米整精米率检验法》[20]检测胶稠度、直链淀粉、整精米率,并依据《NY/T 593-2013食用稻品种品质》[21]标准确定样品品质等级。
1.4 全基因组测序和性状等位基因鉴定
随机选取10株生长30 d左右的江79S植株叶片,等量混合10个单株叶片提取DNA,利用NovaSeq6000高通量测序仪对样品进行测序(Illumina,北京百迈克生物科技有限公司),测序深度为30×。测序完成后按以下步骤对数据进行分析:(1)利用NGSQCtookit v2.3.3软件对原始测序数据进行质控;(2)利用BWA v0.7.17软件将质控后质量合格的读序映射到水稻参考基因组IRGSPv1.0(https://plants.ensembl.org/Oryza_sativa/Info/Index);(3)通过Samtools v1.2将映射后得到的SAM文件转成BAM文件并删除潜在的PCR重复序列;(4)为保证变异的准确性,利用picard-tools v1.108软件再次删除重复序列;(5)最后用GATK v4.2工具包进行变异检测。根据上海师范大学黄学辉团队[22]对水稻重要性状基因等位信息的整理结果(http://www.xhhuanglab.cn/tool/RiceNavi.html),通过检测江79S序列变异情况判断其是否含有光温敏不育和稻瘟病抗性的等位基因。
2 结果与分析
2.1 选育过程
2012年春,种植培矮64S/粳稻H179的F1植株,抽穗期取花药进行组织培养,获得约150株再生苗。再生苗中约有60株为单倍体,表现为植株矮小、不结实。再生苗中正常生长的植株在9月5日前抽穗,其中11株表现不育但长势较好,割茬再生并在海南收获种子,于2013年同时在浙江嘉兴和江苏无锡分期播种,观察其育性和农艺性状。11个DH不育系中S79株系综合表现优异,但生育期长,属于典型晚粳类型,表现雄性完全不育的时间较短。
为培育生育期较短的光温敏不育系,2013年冬采用高能碳离子束处理S79种子,在海南种植获得M2种子。2014年,于5月上旬在浙江嘉兴直播M2种子,8月中旬在约68 000株M2中筛选到9个早抽穗单株。同年在海南陵水,将9个早穗植株割茬再生获得的种子与S79一起按株系种植,观察发现其中一个株系较S79迟抽穗5 d,综合农艺性状好,将该突变系定名为GS79。
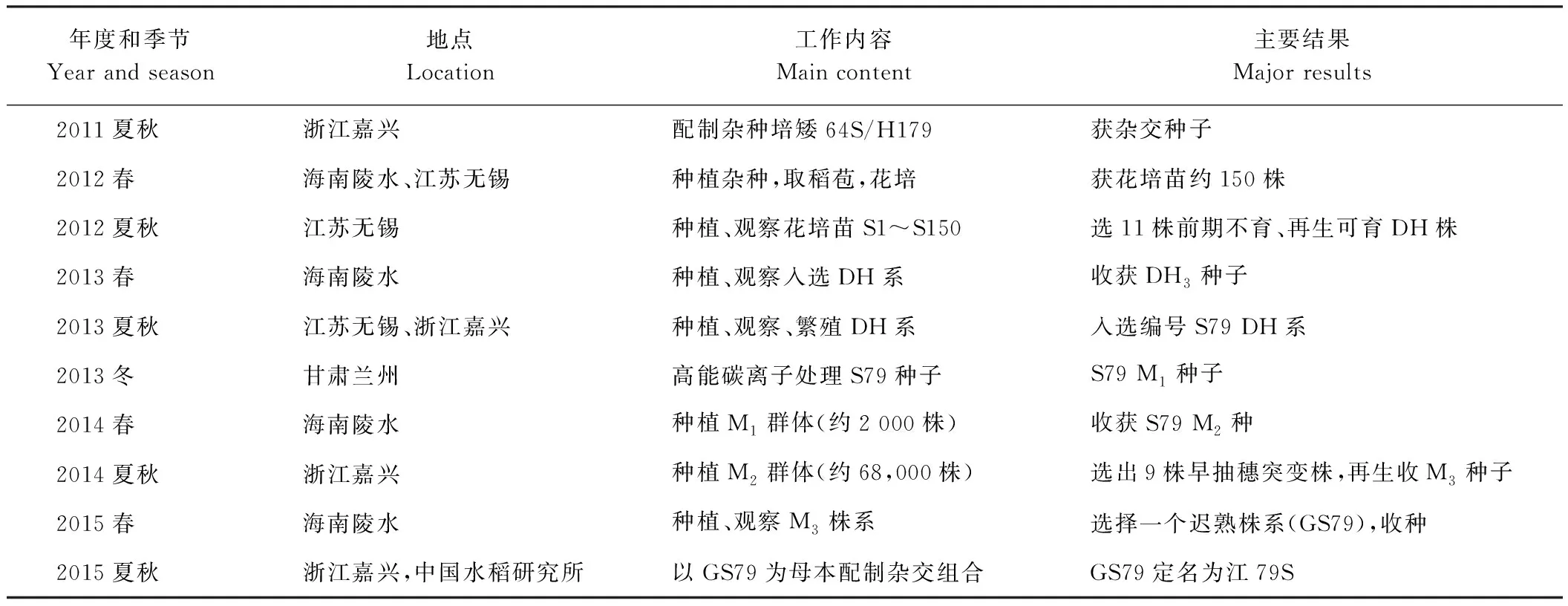
表1 粳型光温敏不育系江79S的选育流程Table 1 The breeding flow of the Japonica photoperiod/temperature sensitive male seterile (PTGM) line Jiang 79S
将突变系GS79和其亲本S79于2015年5月15日在嘉兴同期播种,前者的抽穗时间较后者早约7 d(图1)。之后经多次种植比较,嘉兴5月播种时GS79抽穗较S79早,即播种越早差异越大;海南12月播种时GS79抽穗较S79迟5 d左右。
2.2 育性转换特性
在海南陵水12月中下旬播种,江79S在3月上中旬始穗,表现完全可育,结实率达到80%以上,和常规粳稻接近。即使推迟至1月中旬播种,4月上旬抽穗,育性仍表现正常。上述结果表明该不育系在短日条件下的可繁性好。

图1 水稻花培系S79与其熟期突变体GS79田间表现Fig.1 The performance of S79 derived from anther culture and its heading date mutant GS79 in field
在浙江嘉兴,从5月上中旬开始进行6期的分期播种,8月25日开始隔日镜检,直至9月8日出现极少量黑染花粉,9月15日有大量花粉染成黑色,表明已经转为可育。套袋自交结实考查结果表明,前期结实率均为0,9月8日自交结实率为0.1%,9月15日自交结实率为19.7%,与镜检结果一致。
2015年在人工气候箱中对江79S不同光温条件对育性的影响进行了测试。结果表明,在14.5 h长日照条件下,3个温度(28、24、23℃)处理后,花粉育性和自交结实率均为0,表现完全不育。随着日照时间缩短,经24或23℃处理后,育性有所回复,但平均结实率均维持在较低水平(表2),这与短日照自然条件下的正常结实表现存在较大的差异。
2.3 农艺性状与开花习性
江79S株型适中,分蘖力较强,叶色深绿,叶片较短,植株较矮,抗倒性好。在浙江嘉兴作单季稻种植表现为不育,株高83 cm,单株有效穗数8.2个,平均每穗总粒数可达到125.9粒,包颈率1.2%,柱头白色,柱头基本不外露。在陵水春季(3月初抽穗)种植表现为可育,株高72 cm,单株有效穗数7.1个,平均每穗总粒数达到97.6粒,结实率为81.9%,不包颈,千粒重25.3 g。
江79S为典型粳稻,开花较集中,花期较短。一般秋季晴天11:00左右开花,与粳稻品种相似,但比多数籼稻品种开花时间迟0.5 h以上。
2.4 稻米品质与稻瘟病抗性
经中国水稻研究所-农业部稻米及制品质量监督检验测试中心检测,春季在海南陵水收获的江79S的稻米品质指标如下:糙米率83.5%,整精米率70.4%,垩白米率14%,垩白度2.1,碱消值6.7,胶稠度80 mm,透明度1级,直链淀粉含量15.8%。综合评价达到三级优质米标准。
对江79S开展的苗叶瘟抗性鉴定结果表明,江79S叶瘟平均级1.8级,最高级2级,抗性评价为“抗”。
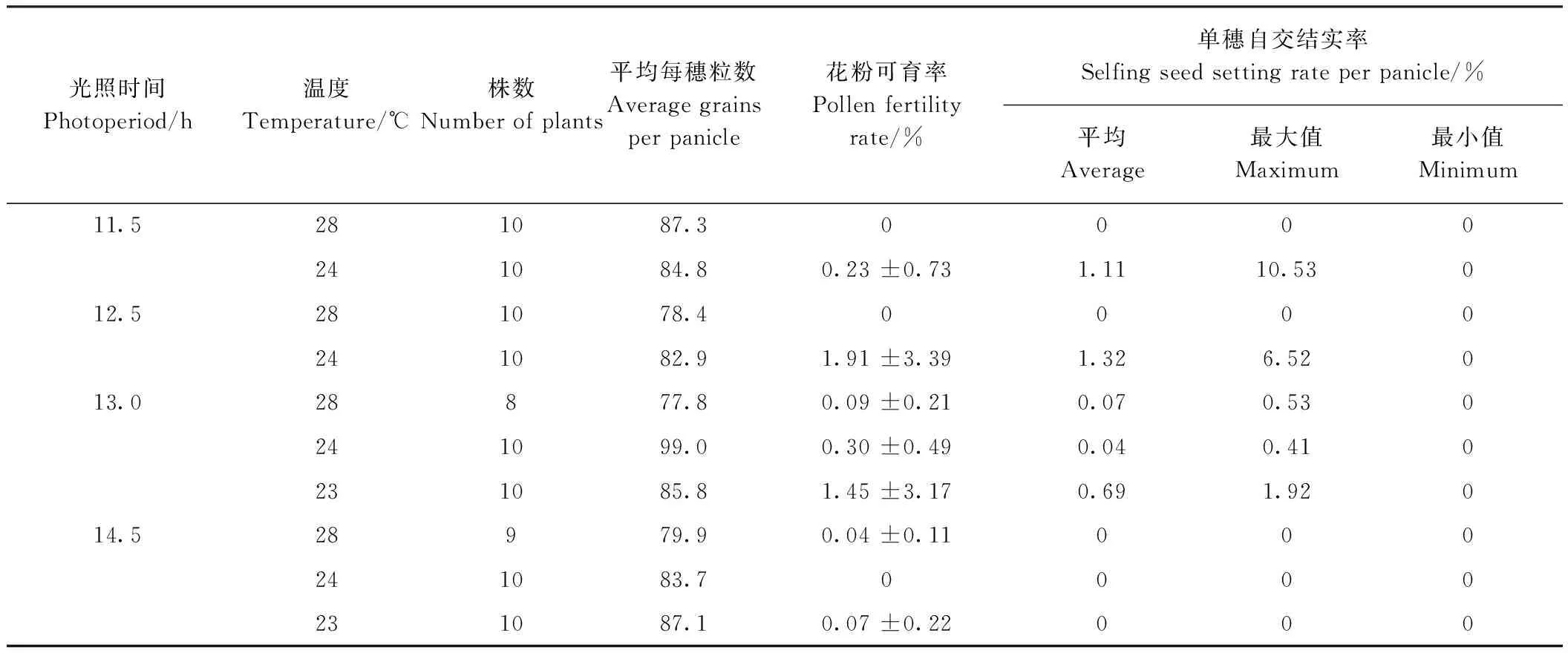
表2 江79S光温敏不育系人工气候箱鉴定结果Table 2 Photoperiod/temperature-sensitivity of Jiang 79S tested in phytotron
2.5 配组应用
以江79S为母本,与广亲和籼稻恢复系江恢1501配制杂交水稻品种江两优7901(图2),该品种于2016年开始参加长江中下游单季晚粳区域试验,两年平均亩产分别达11.4 t·hm2和10.07 t·hm2,比对照嘉优5号分别增产18.2%和15.1%;生产试验亩产平均9.9 t·hm2, 比对照增产9.1%。江两优7901全生育期160.9 d,比对照嘉优5号晚熟0.1 d,株高117.7 cm,穗长21.1 cm,有效穗数217.5万个·hm-2,每穗总粒数260.4粒,结实率80.1%,千粒重24.1 g。稻瘟病抗性综合指数两年分别为3.5、4.3,穗颈瘟损失率最高级3.0,中抗稻瘟病。江两优7901已于2018年通过国家主要农作物品种审定(国审稻20180119)。

图2 江两优7901在浙江新昌田间表现Fig.2 The performance of Jiangliangyou 7901 in Xinchang, Zhejiang
2.6 光温敏不育和抗稻瘟病基因分析
根据已有的重要基因的克隆和功能研究结果,利用全基因组测序结果,分析了江79S中控制光温敏不育和稻瘟病抗性基因情况。基因序列变化会影响表型,由如pms3基因C→G的突变(基因组第22 054 886 bp的位置)导致水稻从可育变为不育。对3个光温敏不育基因分析发现,江79S不含温敏不育基因tms5,但含有光敏不育基因pms1和pms3,与农垦58S和培矮64S一致(表3)。同样,Pi21存在不同等位基因类型,基因组第19 835 834~第19 835 848 bp或第19 836 044~第19 835 865 bp位置序列发生缺失会导致稻瘟病抗性增加。对16个抗稻瘟病基因进行分析,鉴定到江79S含有其中4个抗性基因,即pi21、Pid3、Pia/RGA4和Pi-ta(表3)。
3 讨论
不育系是杂种优势利用的基础,培育优良的粳稻不育系对推动杂交粳稻和亚种间杂交稻的选育尤为重要。与籼稻不育系育种相比,粳稻不育系的数量较少,其中具有产量、品质、抗性等方面优势的不育系更为短缺。本研究采用花培与诱变相关结合培育了一个具有良好配合力的粳稻不育系江79S,有望对杂交粳稻育种和籼粳亚种间优势利用发挥重要作用。在本研究中,采用花药培养技术快速培育不育系,并通过诱变技术对重要性状进行改良,为花培和辐照诱变育种的综合运用提供了范例。
籼粳杂种优势利用以“籼不粳恢”或“粳不籼恢”的育种模式为主。籼型两系不育系育性稳定,“籼不粳恢”模式的花时差异更加有利于制种。虽然两系籼型不育系在生产上开展了大面积的应用,但两系法籼粳亚种存在着粒型表现为中间型、株型偏高的问题[23]。“粳不籼恢”模式粳稻花时迟于籼稻,制种产量低,但可以通过选育早花时、高外露与柱头活力强的粳型不育系解决[24]。此外,粳稻两系不育由核基因控制,相比籼稻更容易通过花药培养、遗传转化、诱变等技术对其进行改良[25-26]。因此,粳稻两系不育系具有生产应用价值且容易创制,不仅能为杂种优势利用提供关键遗传材料,也能为培育强优势杂交粳稻品种提供重要亲本材料。

表3 江79S中光温敏不育和抗稻瘟病基因序列分析Table 3 The sequence analysis of PTGM genes and rice blast resistant genes in Jiang 79S
育种家通常将光敏不育系与常规材料杂交,选育获得新的不育系,但是许多不育系携带的不再是原始亲本的光敏不育基因。培矮64S含有光敏不育基因,但国内的培矮64S为亲本选育的多数新的不育系携带温敏不育基因tms5,如广占63S及其衍生系,其原因尚不明确[27]。江79S系由培矮64S与H179经花培和诱变培育而成,本研究证明其携带与培矮64S一样的光敏不育基因,说明花培和诱变过程未产生新的不育基因。
本研究中,江79S在苗期表现为抗稻瘟病,而全基因组测序结果表明江79S含有pi21、pid3、pia/RG4和Pi-ta基因,表明江79S的稻瘟病抗性表型与基因型一致。虽然在本试验中未开展江79S穗颈瘟抗性的分析,但已有报道表明pi21[28]、Pid3[29]和Pi-ta[30]同样具有穗瘟抗性,因此推测该不育系同样有较强的穗瘟抗性。综上可知,不育系江79S适合与其他材料配制组合,培育出抗稻瘟病的杂交稻新品种。
花药培养和辐照诱变技术结合常规的杂交育种方法同样适用于水稻新品种培育[31],随着基因组学的发展,基因编辑等先进技术方法也用于作物育种中,尤其是用于创制新材料[32-33]。在光敏不育系材料创制方面,林艳等[34]利用TALEN技术编辑日本晴和明辉86光敏核不育基因PMS3,但突变体不能产生光温敏不育表型,由此推测与转化品种背景和pms3位点复杂调控机制有关。农春晓[35]利用CRISPR-Cas9技术获得pms3+orf4-j突变体,初步判断T1突变植株的育性受温度影响,但未有光照时长对突变育性影响的研究。
作为两系不育系,江79S配组自由,特别是与广亲和籼稻恢复系配组,优势强,熟期适中,抗性好,具有重要应用前景。如以其为母本配制的江两优7901(江79S/江恢1501)是一典型的亚种间杂交水稻品种,比甬优12早熟10 d,且产量相仿,具有超高产的潜力。江79S的缺点主要表现在柱头不外露,与籼型恢复系配制的组合因父母本花时相差较大,制种产量偏低,需要通过农艺手段调节或选择适宜的地点,缩小父母本的花时差,从而实现高产制种。
4 结论
本研究利用花培和辐照诱变技术培育了一个早熟、优质、抗稻瘟病的光温敏两系不育系江79S,并以其为亲本,育成国审杂交水稻品种江两优7901。通过对江79S进行全基因组测序,发现其含有来源于农垦58S的2个光敏不育基因pms1和pms3以及4个抗稻瘟病基因,与本试验中观察到的光敏不育特性和抗稻瘟表型一致。
