铜藻中岩藻黄质对H2O2 诱导RAW264.7 细胞氧化损伤的保护作用
2022-05-18杨凌杰胡永东张雷芳闻正顺
杨凌杰,胡永东,张雷芳,闻正顺
(浙江海洋大学食品与药学学院,浙江舟山 316022)
活性氧包括过氧化氢、超氧阴离子自由基、羟基自由基、单线态氧等,自由基过多便引起细胞大分子(DNA、蛋白质和脂质)的氧化损伤[1]。细胞内活性氧的过量产生,并超过内源性抗氧化防御系统的状态被称为氧化应激[2]。在多种人类疾病的发病机制中氧化应激可能起着重要作用,包括动脉粥样硬化、糖尿病、高血压、衰老、肥胖、肾病和癌症[3]。越来越多的证据表明,抗氧化剂可以预防或抑制活性氧引发的细胞凋亡,并可能在治疗因活性氧诱导的疾病中发挥重大作用[4]。由于消费者对天然成分的偏好和对合成抗氧化剂的毒性作用的担忧,用天然来源的抗氧化剂受到了人们的关注。近年来,具有清除过量自由基能力、防止细胞氧化损伤的天然活性物质受到了人们的重视,并开展了各项研究[5-7]。
铜藻Sargassum horneri 又称柱囊马尾藻、海柳麦、草茜,是热带和亚热带水域最显著的褐藻,产于太平洋沿岸及邻近台湾、中国、韩国和日本海域[8]。在东亚,铜藻被用作矿物质、膳食纤维和维生素的丰富来源,成为了受人们关注的多功能保健食品[9]。研究表明铜藻含苯萜类化合物、褐藻糖苷类化合物、甾醇类化合物和糖脂类化合物等,铜藻提取物或分离的纯成分具有广泛的药理活性[10]。岩藻黄质是褐藻等中含有的天然色素,是一种对人类十分有利的活性物质,包括抗炎、抗肿瘤、抗氧化等,日本等国家早已实际应用它,我国对岩藻黄质的研究还不够深入,可以再深入地验证其活性及其机理。NOMURA,et al[11]发现铜藻岩藻黄质含量存在季节性变化,在10 月和11 月呈上升趋势,到1 月达到最大值,之后慢慢下降。结果表明,在冬至春季采收的铜藻中岩藻黄质含量较高。HEO,et al[12]从马尾藻中分离得到岩藻黄质,发现其对H2O2诱导的细胞氧化损伤有较强的抑制作用,且具有剂量依赖性,并通过Hoechst 染色证实这一结果。这些结果表明,马尾藻岩藻黄质可能成为一种治疗氧化应激相关疾病的药物。目前,铜藻岩藻黄质的研究较多集中于含量分析、提取、纯化等阶段,而细胞水平的研究较少。
巨噬细胞对宿主防御系统中微生物病原体的识别和消灭至关重要,是促氧化剂作用的主要目标[13]。因此,RAW264.7 细胞系是筛选抗氧化药物的理想模型[14]。本实验通过建立双氧水刺激RAW264.7 巨噬细胞氧化损伤模型,以评价铜藻岩藻黄质对RAW264.7 巨噬细胞氧化损伤的保护作用,为开发铜藻岩藻黄质预防氧化损伤相关药物和功能性食品提供理论依据。
1 材料和方法
1.1 材料和仪器
铜藻:于舟山嵊泗岛采得,由浙江海洋大学食品与药学学院鉴定,洗净外部泥沙,在阴暗处晾干后粉碎,经80 目过筛,避光储存;RAW264.7 巨噬细胞;DMEM 培养基、胰蛋白酶、胎牛血清:美国Gibco 公司;双抗:北京方程生物科技有限公司;超氧化物歧化酶(SOD)、乳酸脱氢酶(LDH)、谷胱甘肽过氧化物酶(GSHPx)、一氧化氮(NO)检测试剂盒:南京建成生物工程研究有限公司;噻唑蓝(MTT):美国Sigma 公司;DMSO、PBS 缓冲溶液:杭州吉诺生物医药技术有限公司。
旋转蒸发仪:巩义市宇华仪器有限责任公司;冷冻干燥机LGJ-10D:北京四环科学仪器厂;902 型超低温冰箱、311 型细胞培养箱:赛默飞世尔科技有限公司;多功能酶标仪:美国伯腾仪器有限公司;电子分析天平:梅特勒-托利多国际贸易有限公司;超净工作台SW-CJ-2F:苏州安泰空气技术有限公司;高速冷冻离心机:湖南湘仪实验室仪器开发有限公司;Leica 倒置荧光显微镜:美国Leica Microsystems。
1.2 方法
1.2.1 铜藻岩藻黄质溶液的制备
准确称取20 g 铜藻粉末,按液料比1:50 的比例加入70%体积分数的乙醇,在温度70 ℃,超声功率为180 W、超声频率为40 kHz 的条件下避光提取50 min。在上清液中加入1 L 的石油醚,充分振摇后静止分层,将下层有机试剂弃去。最后加入1 L 的甲醇溶液,振摇后静止分层,将下层甲醇溶液收集起来。用旋转蒸发仪浓缩蒸去甲醇溶液后,置于-20 ℃冰箱保存备用。将9.9 mg 岩藻黄质溶于3 mL 二甲基亚砜,密封放置于4 ℃环境备用,配置成5 μmol·mL-1岩藻黄质溶液。
1.2.2 细胞培养
配制含质量分数10%胎牛血清和质量分数1%双抗的DMEM 完全培养基后,在含DMEM 培养基的无菌培养皿中接种复苏后的RAW264.7 巨噬细胞,放置于无菌培养箱中,培养箱中温度为37 ℃、CO2含量为5%。对细胞的生长情况进行观察记录,当细胞在培养皿中长到80%左右时,以1:4 的比例传代,并根据细胞的生长速率判断培养皿中培养液的营养成分是否充足,及时更换培养液。
1.2.3 建立双氧水诱导细胞氧化损伤模型
将RAW264.7 巨噬细胞以104个·mL-1接种于5 块96 孔板,各孔均加入100 μL,放入培养温度为37 ℃、CO2含量为5%的培养箱中培养4 h。观察细胞是否处于贴壁状态,贴壁后将上清液弃去,用PBS 缓冲液洗涤细胞,加入100 μL 由DMEM 完全培养基配制的不同浓度的H2O2溶液,H2O2终浓度为0、0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9 mmol·mL-1,每个浓度设6 个平行。培养时间分别为0、4、8、12、24 h。加20 μL 质量浓度为5 mg·mL-1的MTT 溶液于每个孔,培养4 h 后,1 000 r·min-1离心5 min,弃掉孔板液体,最后每孔加入150 μL DMSO,放于恒温振板仪上振荡15 min,用酶标仪测定570 nm 波长下的OD 值,细胞存活率计算参照公式(1)。

式中:CSR 为细胞存活率,%;A1为空白组的平均吸光度,nm;A2为样品组的平均吸光度,nm。
1.2.4 铜藻岩藻黄质浓度的筛选
细胞培养过程均和1.2.3 节一致,将1.2.3 节所加入的DMEM 完全培养基配制的不同浓度的H2O2换成同培养基配制的不同浓度铜藻岩藻黄质溶液,铜藻岩藻黄质终浓度分别为0、5、10、15、20、25 mmol·L-1。根据公式(1)计算细胞存活率。
1.2.5 氧化损伤模型细胞下铜藻岩藻黄质浓度的筛选
将RAW264.7 细胞以104个·mL-1接种于1 块96 孔板,各孔均加入100 μL,放入培养温度为37 ℃、CO2含量为5%的培养箱中培养4 h。观察细胞是否处于贴壁状态,贴壁后将上清液弃去,用PBS 缓冲液洗涤细胞,加入100 μL 由DMEM 完全培养基配制的不同浓度的铜藻岩藻黄质溶液,铜藻岩藻黄质终浓度为0(空白组)、0(双氧水对照组)、1、5、10、12.5、15、20、25 μmol·L-1,每个浓度设6 个平行,培养24 h。去掉培养液,用PBS 缓冲液洗涤细胞,加入100 μL 由DMEM 完全培养基配制的0.5 mmol·mL-1的H2O2溶液(第一个浓度不加)。再培养12 h。加20 μL 浓度为5 mg·mL-1的MTT 溶液于每个孔,培养4 h,1 000 r·min-1离心5 min,弃掉孔板液体,最后每孔加入150 μL DMSO,放于恒温震板仪上震荡15 min,记录各组570 nm 处吸光度,根据公式(1)计算细胞存活率。
1.2.6 岩藻黄质对双氧水诱导RAW264.7 细胞损伤的保护作用
将RAW264.7 细胞以4×105个·mL-1接种于3 块96 孔板,各孔均加入106个细胞,放入培养温度为37 ℃、CO2含量为5 %的培养箱中培养4 h。观察细胞是否处于贴壁状态,贴壁后将上清液弃去,用PBS 缓冲液洗涤细胞,空白组和双氧水阴性对照组加入完全培养基,铜藻岩藻黄质处理组加入DMEM 完全培养基配制的不同浓度的铜藻岩藻黄质溶液,VC 组加入由DMEM 完全培养基配制的20 μmol·L-1抗坏血酸溶液。每个浓度设置3 个平行,培养24 h,去掉培养液,PBS 洗涤,加入终浓度0.5 mmol·L-1的双氧水溶液。培养12 h,LDH、NO 释放量通过吸取细胞上部培养液测定。SOD、GSH-Px 的含量通过裂解细胞后测定,操作方法都按照试剂盒说明书执行。
1.3 数据分析
实验数据均使用Prism 8 绘图统计分析软件进行画图以及统计分析,分析时使用单因素方差分析以及Tukey 检验进行多重比较,数据结果均以平均值±标准差表示,P<0.05 表示差异具有统计学意义。
2 结果与分析
2.1 RAW264.7 巨噬细胞氧化损伤模型
双氧水可以迅速穿透细胞膜,引起氧化应激。在研究巨噬细胞的凋亡或氧化应激介导的细胞损伤时通常会使用双氧水[15-16]。细胞存活率是反应外部环境对细胞氧化损伤最直接的指标[17],通过MTT 法测定细胞存活率。由图1,图2 可知,与空白组相比,随着双氧水浓度的提高和双氧水作用时间的增长,细胞存活率下降。故双氧水细胞氧化损伤模型的建立采用具有显著差异的0.5 mmol·L-1浓度和12 h 作用时间。
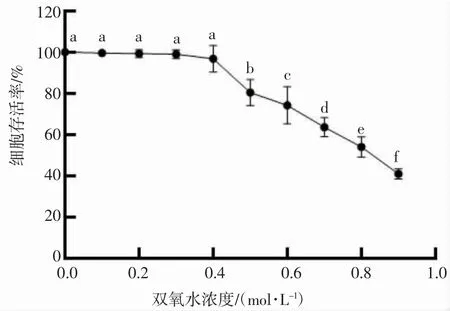
图1 不同浓度H2O2 对RAW264.7 细胞增殖的影响Fig.1 Effects of different concentrations of hydrogen peroxide on the proliferation of RAW264.7 cell

图2 H2O2 时间对RAW264.7 细胞增殖的影响Fig2 Effect of the different time of hydrogen peroxide on the proliferation of RAW264.7 cells
2.2 铜藻岩藻黄质浓度的筛选结果
评估不同浓度的铜藻岩藻黄质对RAW264.7 巨噬细胞的毒性,将铜藻岩藻黄质(5~25 μmol·L-1)预处理RAW264.7 巨噬细胞24 h,计算存活率。由图3 可知,与空白组对比,铜藻岩藻黄质浓度在5~25 μmol·L-1范围时,对细胞增殖均无显著影响(P>0.05),说明该浓度铜藻岩藻黄质对RAW264.7 巨噬细胞无杀伤力,可以进行下一步实验。
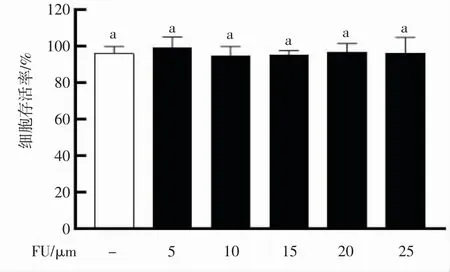
图3 不同浓度岩藻黄质对RAW264.7 细胞增殖的影响Fig.3 Effects of different concentrations of fucoxanthin on the proliferation of RAW264.7 cells
2.3 铜藻岩藻黄质对氧化损伤RAW264.7 细胞增殖的影响
之前实验已确定H2O2为0.5 mmol·L-1浓度和12 h作用时间可建立RAW264.7 巨噬细胞氧化损伤模型。因此,本实验中选用不同浓度的铜藻岩藻黄质探究其对氧化损伤RAW264.7 巨噬细胞的保护作用。如图4可知,与空白组相比,(1~15 μmol·L-1)浓度的铜藻岩藻黄质预处理组和H2O2阴性对照组的细胞存活率显著下降(P<0.05),说明氧化损伤模型建立成功。高浓度的铜藻岩藻黄质预处理后,RAW264.7 巨噬细胞的存活率随着铜藻岩藻黄质浓度的增加而提高。结果表示,25 μmol·L-1浓度的铜藻岩藻黄质组细胞存活率与空白组无显著差异(P>0.05),说明铜藻岩藻黄质对H2O2造成的细胞氧化损伤有一定的保护作用。后续实验将选用15、20、25 μmol·L-13 个浓度。

图4 岩藻黄-质对H2O2 诱导的RAW264.7 细胞增殖的影响Fig.4 Effects of fucoxanthin treatment on H2O2-induced RAW264.7 cell viability
2.4 铜藻岩藻黄质对氧化损伤RAW264.7 细胞SOD 活力的影响
抗氧化酶SOD 在宿主细胞因超氧阴离子引发的氧化应激中发挥至关重要的作用,机理是通过歧化反应催化超氧阴离子分解[18-19]。SOD 活力的上调与抵抗各种非生物胁迫引起的氧化应激有关[20]。由图5 可知,和空白组相比,H2O2损伤组细胞内SOD 活力显著降低(P<0.05)。用不同浓度铜藻岩藻黄质预培养24 h 后,细胞的SOD 活力显著升高(P<0.05)。铜藻岩藻黄质浓度为20、25 μmol·L-1时SOD 活力和VC 阳性对照组无显著差异(P>0.05)。
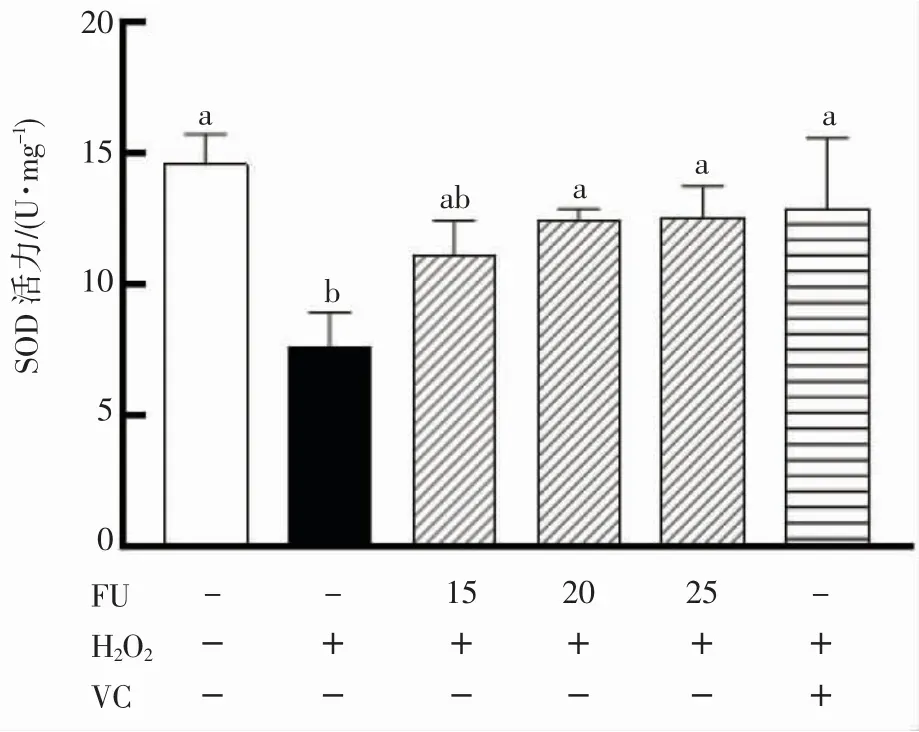
图5 岩藻黄质对RAW264.7 细胞中SOD 活性的影响Fig.5 Effects of fucoxanthin on SOD activity in RAW264.7 cells
2.5 铜藻岩藻黄质对氧化损伤RAW264.7 细胞GSH-Px 含量的影响
GSH-Px 是机体中广泛存在的过氧化酶[21],将H2O2分解成H2O 和氧分子[22]。图6 显示了铜藻岩藻黄质对RAW264.7 细胞GSH-Px 含量的影响。结果表明,H2O2组显著降低细胞中GSH-Px 活力。而20、25 μmol·L-1浓度的铜藻岩藻黄质预培养24 h 后,均可以显著提高GSH-Px 活力(P<0.05),在15 μmol·L-1浓度时没有显著提高。
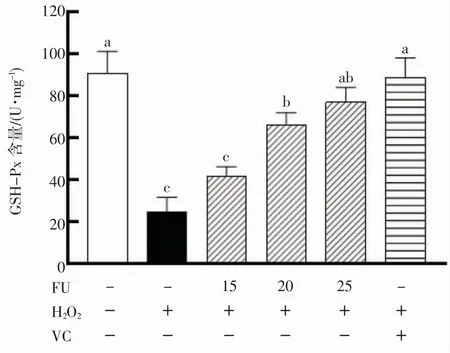
图6 岩藻黄质对RAW264.7 细胞中GSH-Px 活性的影响Fig.6 Effects of fucoxanthin on GSH-Px activity in RAW264.7 cells
2.6 铜藻岩藻黄质对氧化损伤RAW264.7 细胞LDH 活力的影响
通过测定氧化损伤细胞的释放的LDH 活力,用来评价H2O2诱导的细胞毒性和质膜损伤[23]。由图7 可知,与空白组相比,H2O2组LDH 释放量显著升高(P<0.05)。然而,铜藻岩藻黄质预培养24 h 后,使LDH 释放量呈浓度依赖性降低。并且和VC 阳性对照组无显著差异(P>0.05)。

图7 岩藻黄质对RAW264.7 细胞中LDH 活力的影响Fig.7 Effects of fucoxanthin on LDH activity in RAW264.7 cells
2.7 铜藻岩藻黄质对氧化损伤RAW264.7 细胞NO 活力的影响
NO 是具有多种生物学功能的重要调控和效应分子,过多的NO 生成会深刻影响机体的细胞[24]。由图8可知,H2O2诱导RAW264.7 细胞损伤后,NO 释放量显著升高(P<0.05)。而铜藻岩藻黄质预培养24 h 后,可抑制H2O2诱导引起的NO 释放量增高,并呈剂量依赖性。铜藻岩藻黄质浓度为25 μmol·L-1时,NO 释放量与VC 阳性对照组无显著差异(P>0.05)。

图8 岩藻黄质对RAW264.7 细胞中NO 活力的影响Fig.8 Effects of fucoxanthin on NO activity in RAW264.7 cells
3 讨论和总结
巨噬细胞对宿主防御系统中微生物病原体的识别和消灭至关重要,是促氧化剂作用的主要目标。因此,RAW264.7 细胞系是筛选抗氧化药物的理想模型。H2O2是一种常见的氧化应激诱导剂,可通过直接氧化生物分子(脂类、蛋白质、DNA)而损伤细胞,或作为信号分子触发细胞内通路导致细胞死亡[25]。本实验证实了0.5 mmol·L-1H2O2和12 h 作用时间可使RAW264.7 细胞氧化损伤。采用MTT 法检测FU 对RAW264.7 细胞的毒性,并研究在氧化损伤前铜藻FU 是否可以起到细胞保护作用。结果显示,铜藻岩藻黄质(5~25 μmol·L-1)在RAW264.7 细胞无毒性,并且以剂量依赖的方式提高了H2O2诱导的RAW264.7 细胞的存活率。结果说明铜藻岩藻黄质对H2O2诱导的RAW264.7 细胞氧化损伤具有保护作用。
H2O2引起的细胞氧化与H2O2攻击抗氧化系统有关,并导致脂质过氧化物的增加,降低酶活性和细胞膜的通透性[26-28]。SOD 和GSH-Px 是主要的抗氧化酶,可以清除自由基,维持氧化剂和抗氧化剂之间的平衡,是抗氧化损伤的第一道防线[28]。通常来说,当这些抗氧化酶的活性占优势时,ROS-抗氧化平衡和膜的完整性得以维持[29]。因此,我们通过测定抗氧化酶的活性来评估铜藻岩藻黄质对H2O2诱导的细胞氧化损伤的保护作用。本研究发现,H2O2诱导RAW264.7 细胞氧化损伤后,SOD 和GSH-Px 显著降低,说明抗氧化酶防御系统受损。然而,当铜藻岩藻黄质预培养RAW264.7 细胞24 h 后,阻止了H2O2诱导的细胞内抗氧化酶活力降低。并且25 μmol·L-1铜藻岩藻黄质和20 μmol·L-1VC 对H2O2诱导的RAW264.7 细胞氧化损伤具有相似的保护作用。这些结果表明铜藻岩藻黄质可能是通过提高H2O2诱导的RAW264.7 细胞的抗氧化酶活性来减轻氧化损伤。
LDH 释放量通常被用作衡量细胞膜完整性的指标[30]。NO 在调节细胞功能中发挥重大作用[31]。NO 本身是一种自由基,能与O2-反应生成氧化亚硝基阴离子(ONOO-),并在OH-、NO2上分解,对细胞产生急性毒性损伤[32]。本实验发现,铜藻岩藻黄质可以显著降低H2O2诱导的RAW264.7 细胞的LDH 释放量和NO 含量。说明铜藻岩藻黄质可能是通过降低LDH 和NO 的释放量来缓解H2O2对RAW264.7 细胞的损伤。
综上所述,铜藻岩藻黄质可以通过提高细胞中SOD 和GSH-Px 的活性,降低细胞内LDH 和NO 的释放,从而抑制H2O2诱导的RAW264.7 细胞氧化损伤。
