太湖长孢藻属(Dolichospermum)生物量长期动态变化及驱动因子*
2022-05-17朱梦圆邓建明
屈 宁,李 明,张 玮,刘 霞,朱梦圆,邓建明
(1:中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,南京210008) (2:西北农林科技大学资源环境学院,杨凌712100) (3:上海海洋大学水产与生命学院,农业农村部鱼类营养与环境生态研究中心,上海201306)
太湖是我国第三大淡水湖,其水质关系到太湖周边六千多万人口的饮用水安全问题[1]. 自1950s以来,随着太湖地区经济的快速发展和城市化进程加快,水体中的营养盐浓度持续增高. 1960年在夏季开始逐渐出现蓝藻水华,进入1980s以后蓝藻水华事件频繁暴发,尤其是从2006年开始,太湖的蓝藻水华暴发时间提前、结束时间推迟、强度加大、水华面积明显增大[2]. 在2007年5月,太湖蓝藻水华提前大面积暴发,发生了震惊中外的水危机事件,严重影响无锡城市供水,带来了极大的负面影响[3]. 2007年后太湖经历了前所未有的高强度大规模治理[4],有效削减了太湖入湖的氮、磷负荷,富营养化程度得到一定的控制. 但近几年蓝藻水华有所反弹,对太湖水质和生态系统安全造成巨大的威胁[5].
太湖蓝藻种类多样,包括微囊藻属(Microcystis)、长孢藻属(Dolichospermum,曾用名鱼腥藻Anabaena)、束丝藻属(Aphanizomenon)、浮丝藻属(Planktothrix)等. 太湖的蓝藻水华通常以微囊藻属为绝对优势[6]. 然而近年来,不少报道指出长孢藻的比例有增加趋势,尤其是在水华形成初期的春季. 杨柳燕等通过对太湖11年的调查发现,冬季和春季长孢藻属是东太湖和南部沿岸区的优势蓝藻属[7],太湖梅梁湾的长孢藻含量占总蓝藻的8.4%,而在湖心区的占比更高,是太湖蓝藻第二大优势类群[8]. 张民等通过对太湖1993-2015年野外数据调查分析,发现梅梁湾至湖心区的长孢藻生物量呈现较大波动变化,并升高明显[9].
微囊藻与长孢藻的演替规律及驱动因子一直是水华相关研究关注的焦点. 在全球范围内,长孢藻水华出现频率仅次于微囊藻水华[10]. 长孢藻的形态、生理和有害产物具有更高的多样性. 长孢藻是一类含有异形胞的丝状蓝藻,具有固氮作用. 当水体中氮元素缺乏时,长孢藻等固氮蓝藻能通过固氮作用以满足自身的需求[11]. 相比于微囊藻,长孢藻通常被认为更适应低温环境,最适温度在25℃左右,温度进一步升高导致其比增长速率明显下降[12]. 同时,部分微囊藻能产生藻毒素抑制其他藻类的生长[13],进一步诱导藻类群落结构发生演替[14]. 在我国富营养化湖泊巢湖中,每年水华时期微囊藻和长孢藻都会出现演替[15-16]. 在巢湖中,这种蓝藻水华优势物种的演替时间很短,通常持续不到1个月[17]. 野外监测数据显示,巢湖春季湖水中氮浓度急剧下降[17-18],而这一限制,可能会提高固氮蓝藻如长孢藻在浮游植物群落中的竞争优势. 通过对五里湖的长期监测结果表明,由于潜在的氮限制,浮游植物的优势种群可能由微囊藻转向固氮蓝藻长孢藻[19]. 但目前还不清楚,太湖地区微囊藻与长孢藻之间演替的内在规律,以及长孢藻丰度的动态变化除了与氮元素相关外是否与其他因子存在密切关联.
本文以太湖为研究对象,探究太湖不同湖区春季长孢藻的长期变化趋势及驱动因子. 根据以前的研究,太湖近年来水体中营养盐浓度和局地的气候条件都发生了显著变化[20]. 经过高强度的治理和修复后,太湖水体中氮的浓度显著下降,可能引起氮限制,从而增加长孢藻的竞争优势[19];此外,太湖地区冬春季增温明显,同时风速显著下降,而且气候变化对太湖春季浮游植物的影响存在跨季节效应,即冬季的气候状况会显著影响来年春季的浮游植物生物量及群落组成[21]. 我们推测太湖地区春季长孢藻的变化趋势可能也与冬春季气候变化和营养盐浓度有关. 因此提出假设:营养盐浓度,尤其是氮浓度下降和冬季气象条件改变可能是太湖春季长孢藻优势上升的两个主要驱动因子. 为检验上述假设,本文选取太湖2005-2019年期间太湖冬、春季气象条件和营养盐浓度以及太湖春季长孢藻生物量进行分析和讨论,为亚热带浅水湖泊冬季条件与春季浮游植物群落演替的关系提供新的视角.
1 材料与方法
1.1 研究区域
太湖(30°55′40″~31°32′58″N,119°52′32″~120°36′10″E)属于大型浅水湖泊,位于长江三角洲地区,是中国工业化和人口最密集的地区,面积约2238 km2,多年平均水深1.9 m. 由于快速的经济增长和强烈的人类活动,太湖目前处于富营养化状态,蓝藻水华在春季发生并持续整个夏秋季[22].
1.2 气象数据获取
为研究太湖地区气象条件的长期变化趋势,本文选取太湖周边的无锡、东山、湖州3个气象观测基本站2005年1月1日-2019年12月31日的逐日气象数据,数据来自中国气象数据共享服务系统(http://cdc.cma.gov.cn/,站点号分别为58354、58358、58450). 这3个气象站点分别位于太湖的3个方位,能较好地表征太湖地区气象条件的平均变化趋势. 气象指标包括风速(m/s)、气温(℃)、日照时长(h)和降雨量(mm). 通过逐日观测数据分别计算季节平均气温、季节平均风速、季节累计日照时长、季节累计降水量等. 四季划分如下:冬季(12月-次年2月)、春季(3-5月)、夏季(6-8月)和秋季(9-11月).
1.3 太湖分区采样
由于湖泊面积大,太湖生境多样,各个区域水体营养水平、沉积物性质和大型植物分布存在显著的空间差异,发生蓝藻水华的时间和强度都有所区别[23-24]. 自1980s以来,太湖流域由于大量污水的排入,太湖北部和中部水质快速恶化,富营养化加速,水华暴发已成为常态现象[25]. 梅梁湾位于太湖北部,是太湖富营养化程度最高的区域之一,其水华强度高于其他区域[26],属于藻型湖区. 太湖湖心区营养水平较低,处于中营养水平. 竺山湖是太湖水质污染最严重的湖区之一[27],且一直以来都是太湖蓝藻水华暴发较为严重的区域[28]. 上述湖区一直是太湖典型的藻型生态系统. 太湖西部地区属于河口区,河口区作为河流和湖泊之间的过渡区域,受到陆地污染源沿河汇入以及河流和湖泊的双重影响[29-30],藻类群落结构变化具有特殊性,不适合分析太湖藻类长期变化趋势以及影响因素. 贡湖湾和东太湖是太湖的草型生态系统,在雨季水生植物覆盖度较高,也是长江中下游典型的草型浅水湖泊生态类型[31];水生植物对藻类有一定抑制作用,在滇池和洱海湖滨带也有相似的结果[32-33]. 研究期间,贡湖湾和东太湖长孢藻生物量很小,因此本文只选择竺山湖、梅梁湾、湖心区以及南部湖区等典型的藻型生态系统进一步开展研究.
按照不同生境,太湖共布设了14个样点,涵盖了太湖的梅梁湾(THL1、THL3~THL5、THL32)、竺山湖(THL16、THL17)、南部湖区(THL21~THL23)和湖心区(THL7、THL8、THL18、THL19)共4个区域(图1). 在各个监测点位于2005-2019年逐月采集水样测定水质指标并且采集浮游植物样品来确定浮游植物群落结构.

图1 太湖采样点分布Fig.1 Distribution of sampling sites in Lake Taihu
混合水样采用长2 m、直径为10 cm的塑料管进行采集. 采用国家标准方法进行理化指标的测定,包括水深(depth)、pH、悬浮物(SS)、电导率(Cond)和营养物质浓度. 总氮(TN)浓度采用碱性过硫酸钾消解紫外分光光度法(GB 11894-1989)测定,总磷(TP)浓度采用钼酸铵分光光度法(GB 11893-1989)测定.
每个采样点共采集1 L混合水样,用于浮游植物的鉴定和计数. 浮游植物样品用鲁哥试剂固定,沉淀72 h后采用虹吸法去除上清液并浓缩至30 mL,参照文献[34]对浮游植物种类进行鉴定. 计数时取0.1 mL在显微镜下放大400倍计数,浮游植物鉴定到种或属. 由于浮游植物的比重接近1,故可直接将浮游植物的体积换算成生物量(湿重,单位为mg/L),单细胞的生物量主要根据浮游植物个体形状测量而得[35].
1.4 统计方法
综合考虑理化和时空因素的影响,计算太湖4个区域各采样点的季节平均值,采用广义可加模型(generalized additive models,GAM)比较了2005-2019年不同湖区的长孢藻长期变化趋势. 广义可加模型能更好地处理此类非正态分布、非线性关系的数据[36]. 本文主要使用mgcv软件包中gam函数实现[37]. 首先进行方差膨胀因子(VIF)检验,排除共线性因素[38]. 使用矫正后的r2和P值评估模型的拟合度和显著性[36]. 在进入GAM模型拟合之前,对数据进行预处理,气象因素计算出3个气象站点的季节平均值,营养条件和长孢藻生物量计算出不同湖区的季节平均值,代入模型进行长期变化趋势预测.
采用偏最小二乘路径建模(partial least squares path modeling, PLS-PM)方法,以春季长孢藻生物量为研究对象,评估了冬季气候因子(风速、气温、日照时长、降雨量)、冬季营养水平(TN、TN/TP)、春季气候条件(风速、气温、日照时长、降雨量)和春季营养水平(TN、TN/TP)与春季长孢藻生物量的关系,最后量化各气候条件和营养盐对长孢藻生物量变化的贡献大小. 这种方法被称为结构方程模型的偏最小二乘法(SEM),可以评估带有潜在变量的复杂因果关系模型[39]. PLS-PM一般采用GOF指数(goodness of fit,GOF)判断模型整体拟合程度,GOF值越大,模型拟合程度越高:GOF=0.10,模型拟合程度低;GOF=0.25,模型拟合程度中等;GOF=0.36,模型拟合程度高[40]. 在R中使用plspm运行PLS-PM[39].
2 结果
2.1 太湖2005-2019年气象因子长期变化趋势
太湖冬季平均温度为(5.6±0.87)℃,2005-2019年冬季气温无明显变化规律(GAM,下同,P>0.05). 冬季最高温度出现在2017年,为7.12℃,在2012年温度出现最低值(3.7℃). 冬季平均日照时长为333.58 h,2005-2010年基本保持不变,但2011-2019年略有下降. 2007-2009年太湖周边冬季风速有所上升,2009-2019年显著下降(r2=0.72,P<0.01). 冬季降雨量从2005年的260 mm降至2011年的105 mm左右,随后又上升至200 mm左右(图2).
太湖春季平均温度为(16.405±0.75)℃,2005-2019年春季气温无明显变化规律(P>0.05),在2010年出现最低值(14.28℃). 春季平均日照时长为516.61 h,近15年无明显变化规律(P>0.05),且2016年最低(273.83 h). 太湖春季平均风速逐年显著下降(r2=0.74,P<0.01),春季风速大都小于3 m/s. 春季平均降雨量为279.77 mm,2005-2019年降雨量无明显规律(P>0.05)(图3).
2.2 太湖不同湖区水质长期变化趋势及区域差异
太湖4个湖区冬、春季TN、TP浓度之间存在差异. 冬、春季TN、TP浓度最高的是竺山湖,冬、春季TN浓度最低的是南部湖区,冬季梅梁湾TP浓度最低,春季南部湖区TP浓度最低(附图Ⅰ). 太湖这4个湖区中,竺山湖是富营养化程度最高的湖区.
太湖不同湖区冬季营养盐浓度的长期变化趋势也存在差异. 2005-2019年冬季,梅梁湾TN浓度呈显著下降趋势(r2=0.96,P<0.01),从5.8 mg/L下降至约2 mg/L;湖心区TN浓度呈显著下降趋势(r2=0.59,P<0.01)并且2015-2019年保持在平均值为3 mg/L左右;竺山湖TN浓度偏高,这15年间TN浓度都在5 mg/L以上,从2010年之后有明显下降趋势;南部湖区TN浓度无明显变化趋势(P>0.05)(图4).
2005-2019年冬季,太湖4个湖区TP浓度的长期变化趋势存在空间差异. 梅梁湾冬季TP浓度总体呈显著下降趋势(r2=0.68,P<0.01),从2006年最高的0.135 mg/L降至0.07 mg/L左右;湖心区冬季TP浓度除了2012年达到最高(0.18 mg/L),其余年份基本平稳在0.12 mg/L左右;竺山湖冬季TP浓度在4个湖区中最高,平均为0.188 mg/L,2005-2008年TP浓度持续增高,2008年以后呈下降趋势;南部湖区冬季平均TP浓度为0.114 mg/L,2005-2019年无明显变化趋势(P>0.05)(图4).

图2 2005-2019年冬季太湖气温、日照时长、风速和降雨量的长期变化趋势(实线代表广义可加模型 (GAM)估计的长期趋势,阴影区域是估计的标准误差.校正后的r2;**: P<0.01; *: P<0.05)Fig.2 Long-term trends of meteorological factors (air temperature, sunshine duration, wind speed and rainfall) in Lake Taihu in winter from 2005 to 2019 (The solid line represents the long-term trends of annual mean values estimated by generalized additive models (GAM), and the shaded area represents the standard error of the estimate, r square of GAM;**: P<0.01; *: P<0.05)
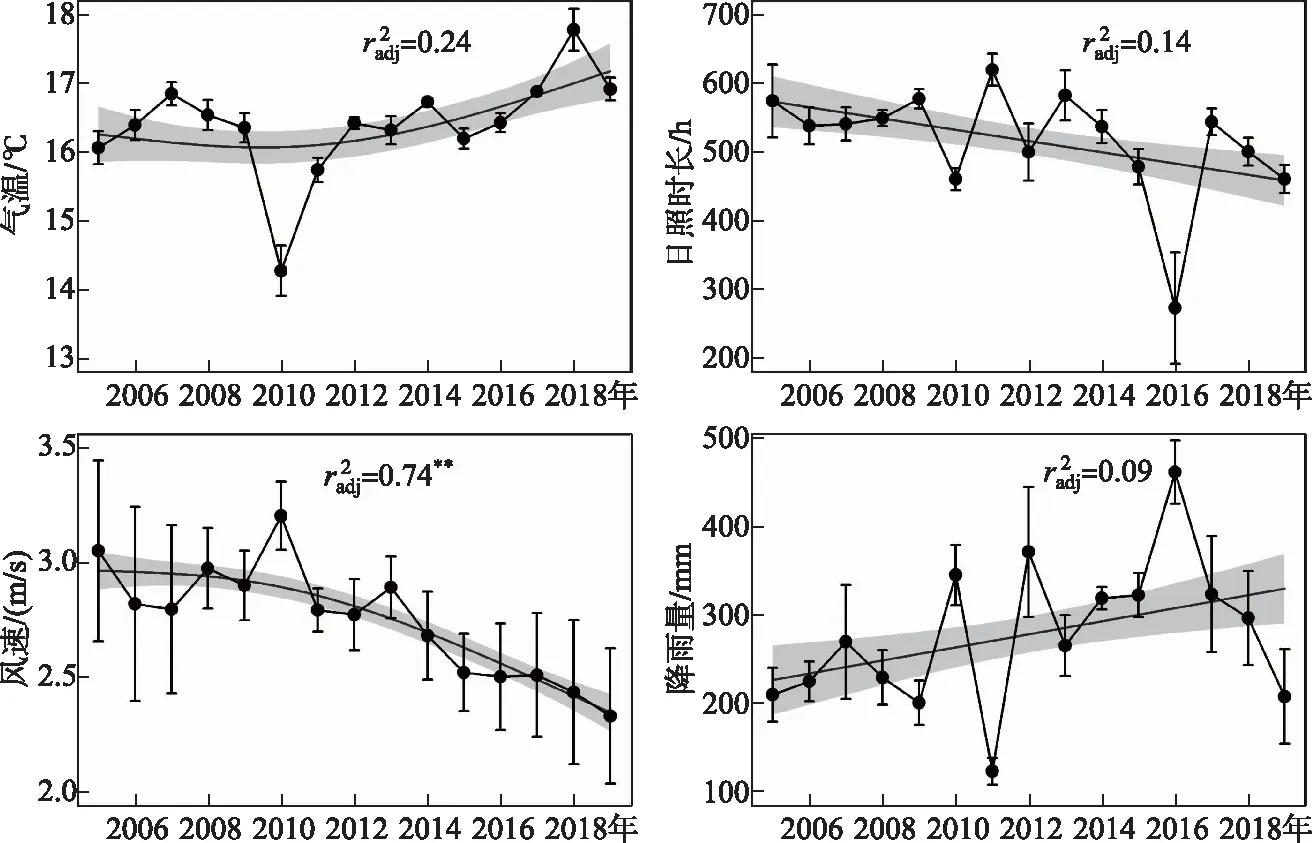
图3 2005-2019年春季太湖气温、日照时长、风速和降雨量的长期变化趋势(实线代表广义可加模型 (GAM)估计的长期趋势,阴影区域是估计的标准误差.校正后的r2;**: P<0.01; *: P<0.05)Fig.3 Long-term trends of meteorological factors (air temperature, sunshine duration, wind speed and rainfall) in Lake Taihu in spring from 2005 to 2019 (The solid line represents the long-term trends of annual mean values estimated by generalized additive models (GAM), and the shaded area represents the standard error of the estimate, r square of GAM;**: P<0.01; *: P<0.05)

图4 2005-2019年冬季太湖不同湖区总氮(TN)、总磷(TP)浓度的长期变化趋势(实线代表广义可加 模型估计的长期趋势,阴影区域是估计的标准误差.校正后的r2;**:P<0.01;*:P<0.05)Fig.4 Long-term trends of total nitrogen and total phosphorus concentrations in different areas of Lake Taihu in winter from 2005 to 2019 (The solid line represents the long-term trends of annual mean values estimated by generalized additive models (GAM), and the shaded area is the standard error of the estimate, r square of GAM;**: P<0.01; *: P<0.05)
太湖不同湖区春季营养盐浓度长期变化趋势也存在差异. 2005-2019年春季,梅梁湾、湖心区、竺山湖以及南部湖区TN浓度均呈显著下降趋势(P<0.01). 梅梁湾春季TN浓度从6.8 mg/L下降至约2.3 mg/L;湖心区春季TN浓度从5.5 mg/L降低至2.5 mg/L左右;竺山湖春季TN浓度最高,这15年间TN平均浓度为6.1 mg/L;南部湖区春季TN浓度总体呈下降趋势,但在2010年出现最大值(图5).
太湖4个湖区春季TP浓度存在空间差异. 2005-2019年春季4个区域TP浓度的长期变化趋势均无显著变化规律,竺山湖春季TP浓度总体呈下降趋势(图5).

图5 2005-2019年春季太湖不同湖区总氮(TN)、总磷(TP)长期变化趋势(实线代表广义可加模型 估计的长期趋势,阴影区域是估计的标准误差.校正后的r2;**: P<0.01; *: P<0.05)Fig.5 Long-term trends of total nitrogen and total phosphorus concentrations in different areas of Lake Taihu in spring from 2005 to 2019 (The solid line represents the long-term trends of annual mean values estimated by generalized additive models (GAM), and the shaded area is the standard error of the estimate, r square of GAM;**: P<0.01; *: P<0.05)
2.3 太湖不同湖区长孢藻组成比例变化趋势
总体而言,太湖各个区域春季长孢藻生物量过去15年来都有增加的现象(图6). 本研究发现竺山湖春季长孢藻生物量最多,其次是湖心区. 2005-2019年梅梁湾区域长孢藻生物量总体有增加的现象(r2=0.31,P<0.05),平均值为0.09 mg/L,2016年长孢藻生物量达到最大值0.47 mg/L(图6),并且占优势蓝藻的比例为80%(附图Ⅱ);湖心区春季长孢藻生物量在2011-2018年持续增加,2005-2019年春季长孢藻平均生物量为0.11 mg/L,占优势蓝藻的平均比例约为32%(附图Ⅱ);春季竺山湖长孢藻总体有上升趋势,2005-2019年平均生物量为0.15 mg/L,并且在2016年达到最大值,生物量为1 mg/L左右,占优势蓝藻的比例大于60%(附图Ⅱ). 南部湖区2005-2019年春季长孢藻平均生物量最低,约为0.05 mg/L.
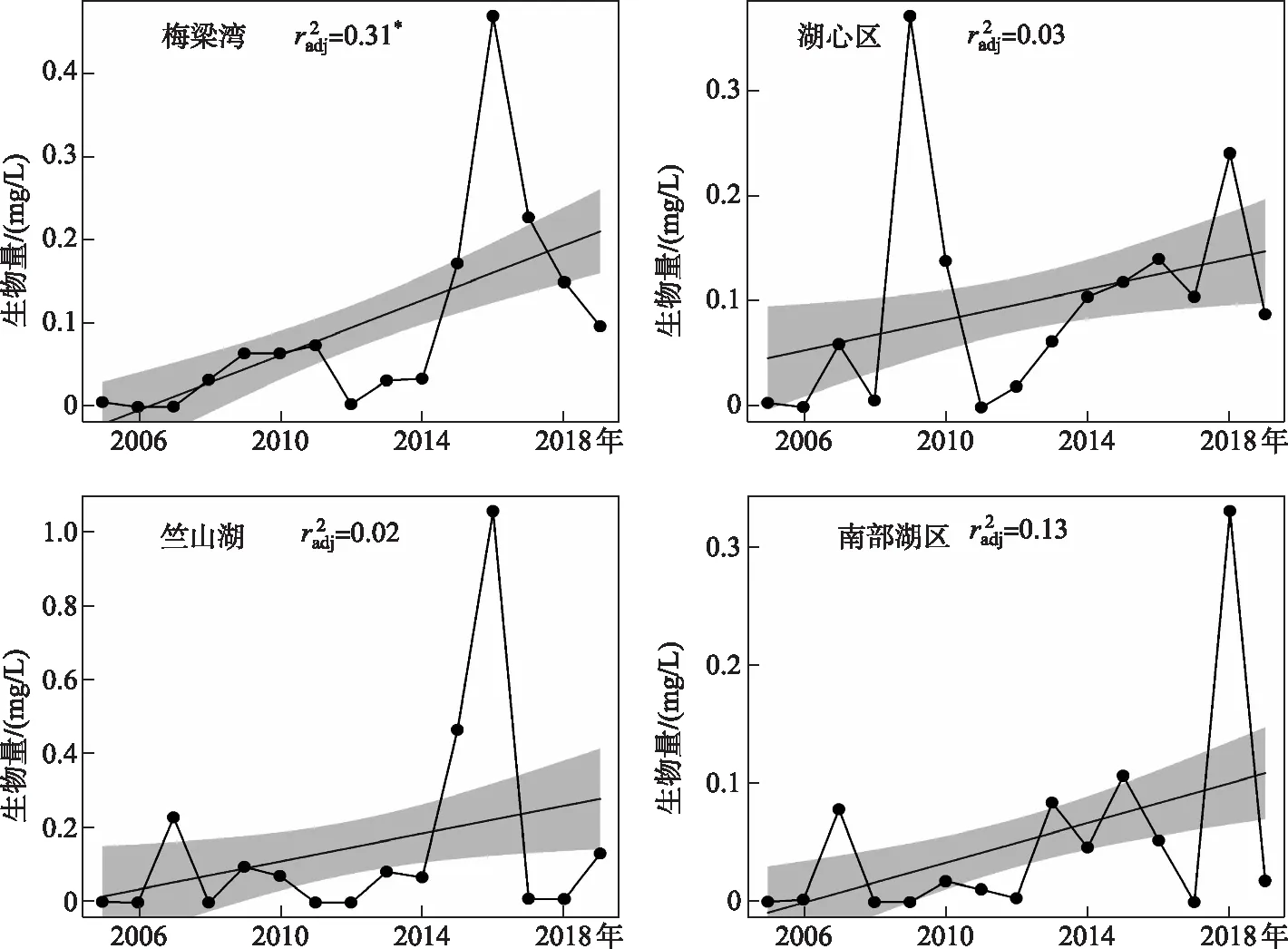
图6 2005-2019年春季太湖不同湖区长孢藻生物量的长期变化趋势(实线代表广义可加模型 估计的长期趋势,阴影区域是估计的标准误差.校正后的r2;**:P<0.01, *:P<0.05)Fig.6 Long-term trends of Dolichospermum biomass in different areas of Lake Taihu in spring from 2005 to 2019 (The solid line represents the long-term trends of annual mean values estimated by generalized additive models (GAM), and the shaded area represents the standard error of the estimate, r square of GAM;**: P<0.01; *: P<0.05)
2.4 环境因子对长孢藻生物量变化的贡献
在太湖,春季长孢藻生物量受到冬、春季营养物质以及冬、春季气候条件不同程度的直接或间接影响. 梅梁湾、湖心区、竺山湖和南部湖区模型拟合GOF值分别为0.53、0.40、0.46和0.36(图7),模型拟合程度较好,结果可信. 梅梁湾春季长孢藻生物量受春季气候条件的显著影响,其次是春季营养盐(图7a);湖心区春季长孢藻生物量主要受冬季气候条件的显著影响,其次是春季气候条件(图7b);竺山湖和南部湖区都受到春季气候条件的直接显著影响,并受冬季气候条件的间接显著影响(图7c,d).
在梅梁湾湖区,春季气候条件和春季营养盐(日照时长、风速和氮磷比)在春季长孢藻生物量的长期变化中发挥着重要作用(附图Ⅲa),春季营养盐主要受冬季营养盐和春季气候条件的影响;湖心区,春季长孢藻生物量主要受冬季气候条件(冬季风速、温度、日照时长)影响,同时也受春季气候条件(春季风速、温度)影响(附图Ⅲb);竺山湖春季长孢藻生物量主要受春季气候条件(春季风速、日照时长)和冬季气候条件(冬季风速、日照时长)的影响(附图Ⅲc);南部湖区春季长孢藻生物量取决于春季气候条件(春季温度、风速)和冬季气候条件(冬季风速)(附图Ⅲd).
3 讨论与结论
本研究利用2005-2019年太湖地区气象、环境和生物要素监测数据,评估了我国亚热带大型浅水湖泊太湖冬、春季气候条件和营养水平对春季长孢藻生物量长期变化趋势以及空间差异的影响. 结果显示太湖不同湖区近些年来春季长孢藻生物量均有升高的现象,尤其是在富营养程度较高的竺山湖,其次是梅梁湾、南部湖区等区域(图6).
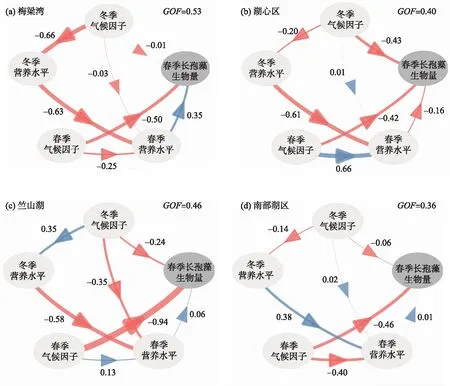
图7 偏最小二乘路径建模(PLS-PM)确定冬季气候条件、冬季营养盐水平、春季气候条件和春季营养盐水平 对春季长孢藻生物量的影响路径(蓝色箭头表示呈正相关;红色箭头表示负相关,数字代表贡献相对大小)Fig.7 The effect paths of winter meteorological factors, winter nutrients, spring meteorological factors, spring nutrients on the spring Dolichospermum as determined by partial least squares path modeling (PLS-PM) (The blue arrow indicates a positive correlation; the red arrow indicates a negative correlation, and the numbers represent the relative contribution)
不同湖区长孢藻生物量增加的主要驱动因素存在差异(图7). 太湖是典型的大型浅水湖泊,营养物质、沉水植物、浮游植物等因子的空间分布具有显著的空间异质性[23-24],这可能是导致长孢藻生物量变化的主要驱动因子存在空间差异的原因. 在湖心区,由于吹程较大,频繁受到风浪等扰动,水体长期处于浑浊状态,因此风速降低能增加水下光场,促进长孢藻形成优势;冬季出现较高温度时可能会促进冬季微囊藻的生长[41],导致以微囊藻为优势种的蓝藻水华在早春提前暴发,同时降低春季长孢藻的生物量. 湖心区TN浓度显著降低至较低水平(图5),春季营养盐与春季长孢藻生物量呈负相关(图7),导致湖心区春季长孢藻生物量增加(图6). 这与以往的控制实验和观测结果相一致[41-42]. 太湖长孢藻生物量的波动趋势与TN浓度的波动趋势相反. 湖心区也是营养盐水平偏低的区域,所以藻类生长容易受到营养盐的制约. 梅梁湾、竺山湖等湖区是承接外源污染负荷的主要区域,均属于富营养化程度较高的区域. 在太湖的研究表明,营养盐水平较高时,气候条件成为浮游植物群落演替的主要影响因素. 春季较低的风速和较短的日照时长均显著促进了长孢藻的生长(附图Ⅲa,c,d). 在梅梁湾湖区,春季TN/TP对春季营养盐水平的贡献比较大(附图Ⅲa),导致春季营养物质对春季长孢藻生物量的影响较大,与其他湖区相比,春季营养盐的贡献要大于冬季气候的贡献,这可能是因为梅梁湾湖区的营养盐,尤其是TN浓度,相比于其他湖区下降最为明显(图5). 同时春季气候条件是梅梁湾湖区春季长孢藻生物量增加的最重要影响因素. 与梅梁湾不同,竺山湖春季营养物质对春季长孢藻生物量的影响很小,是因为在营养盐水平相对较高时,长孢藻生物量与营养盐浓度没有关系[42]. 气候变化在富营养湖泊中的影响大于贫营养湖泊,特别是对冬、春季生物活性极低的蓝藻而言[43]. 长孢藻属于固氮蓝藻,在不同湖区长孢藻营养盐利用效率可能存在差异,导致不同湖区营养盐对春季长孢藻生物量的贡献不同.
基于不同湖区的分析结果,研究结论与我们前面的假设基本一致,春季气候条件主要影响春季长孢藻生物量变化,同时,值得注意的是冬季气候条件对太湖春季长孢藻生物量变化也有重要的影响,尤其是在湖心区;同时冬季气候条件还是竺山湖和南部湖区春季长孢藻的次要影响因素. 冬季气候可以通过两种途径来影响春季长孢藻生物量[21]. 首先,冬季气候对冬季湖区内浮游植物群落结构和藻类生物量产生直接影响,进而影响春季浮游植物群落结构和藻类生物量. 太湖是典型的大型浅水湖泊,受风浪影响水体通常比较浑浊,在富营养化湖区透明度仅有20 cm左右,因此光照强度是冬季和春季浮游植物演替重要的影响因素[44]. 长期气象数据结果显示,太湖15年来冬季风速明显下降(图3),较低的风速会减少沉积物再悬浮,降低水柱中悬浮物浓度,从而改善水下光场条件,引起藻类群落结构发生变化[45]. 普遍认为在温带湖泊中温度是影响浮游植物生长的重要因素[46-47],但是我们的研究发现,冬季增温并没有成为春季长孢藻生物量变化的最重要因素,这可能是因为太湖冬季平均气温(2005-2019年冬季平均气温为(5.6±0.87)℃)比较高[48],浮游植物在冬季并没有停止生长,而是以相对较低的生长速率生长,并在春夏季外界条件适宜时快速繁殖,形成水华[49]. 另一个途径是冬季气候条件可以影响冬季的营养水平,从而影响春季的营养盐浓度,最终影响长孢藻的生长[43]. 风速下降增加了水柱稳定性,从而导致底部出现缺氧/厌氧条件[50],加剧底泥中有机态营养盐矿化降解并释放到水柱中[51-52]. 本文结果显示第一个途径比后者扮演着更重要的角色,这可能是因为太湖整体上属于富营养湖泊,浮游植物类群一般受湖泊物理因素影响而对营养盐水平变化不再敏感. 太湖营养盐水平较高,气候条件成为浮游植物群落演替的主要影响因素[53]. 研究表明,当TP浓度大于0.059 mg/L 时,藻类生长速率和生物量不随磷浓度增加而增加[54]. 当氮、磷浓度均比较充足时,氮磷比对蓝藻生长速率已经没有影响[55]. 之前有关太湖、巢湖和滇池长孢藻影响因素的研究表明,长孢藻生物量表现出湖泊特异性,并且在太湖长孢藻的变化与营养盐浓度没有关系[42]. 与之类似,本文发现在营养盐水平较高的梅梁湾、竺山湖、南部湖区,春季气候条件对春季长孢藻生物量的影响更重要.
总的来说,太湖不同湖区长孢藻生物量近年来均有所增加,但不同湖区的主要影响因素不同. 在湖心区,冬季气候条件、春季营养盐水平和春季气候条件是春季长孢藻生物量变化的主要影响因子;而在梅梁湾、竺山湖和南部湖区,由于营养盐水平相对较高,气候因子是春季长孢藻生物量变化的重要影响因子. 上述结果说明气候变化对湖泊的影响受到不同湖区本身特征的调节.
致谢:感谢中国科学院南京地理与湖泊研究所太湖湖泊生态系统研究站提供太湖历史监测资料;感谢何云川和张庆吉等在数据分析中给予的帮助.
4 附录
附图Ⅰ~Ⅲ见电子版(DOI: 10.18307/2022.0303).
