轻度人为干扰下贵州道地药用植物艾纳香群落特征研究
2022-05-17田凡李鹏廖小锋
田凡 李鹏 廖小锋

摘要:为探究艾纳香群落基本特征及群落内的生存状况,采用样方调查法对贵州省罗甸县红水河地区艾纳香植物群落进行调查,并从植物区系、生活型、重要值以及物种多样性等角度进行了分析。结果表明:艾纳香群落共有64种植物,隶属于24科57属,以禾本科、菊科、豆科的植物为主。群落内以高位芽和地面芽植物为主,分别占群落植物种数的46.87%和31.25%。群落无乔木层,艾纳香在灌木层和草本层中重要值分别达到了83.23%和26.68%。艾纳香群落的物种丰富度指数为10~27,Shannon-Wiener多样性指数为1.416~2.120,Pielou均匀度指数为0.523~0.688,Simpson优势度指数为0.260~0.433。总体而言,因为人为干预的影响,艾纳香处于群落建群种和单优势种地位,群落物种多样性水平较低,并且取决于草本层的物种丰富程度;艾纳香的生存发展状况对群落物种组成、外貌特征、稳定性和演替进程具有很大影响。
关键词:贵州;艾纳香;群落特征;物种多样性;重要值
中图分类号:Q145文献标识码:A
文章编号:1008-0457(2022)03-0057-07国际DOI编码:10.15958/j.cnki.sdnyswxb.2022.03.008
艾纳香[Blumea balsamifera (Linn.) DC.]为菊科艾纳香属(Blumea)多年生木质大草本,是获取天然冰片的重要原料植物,也是贵州省十大苗药之一[1]。艾纳香在贵州主要分布于南北盘江及红水河流域的罗甸、望谟、册亨、兴义、安龙、贞丰等县份低海拔河谷地区[2],其中罗甸县是艾纳香最为典型的分布区,“罗甸艾纳香”也被批准为国家地理标志保护产品。由于广西龙滩水电站于2009年建成蓄水,艾纳香最适分布区罗甸县红水河河谷地带被大范围淹没[3],并且因市场大幅波动,贵州省艾纳香集中种植地罗甸县的人工种植规模已由前几年10 000 hm2高位骤降至目前仅有333.333 hm2,种苗基地也已全部转产退出[4],这势必极大影响该地区艾纳香野生资源及家系资源状况,并有可能对艾纳香产业后续恢复发展造成种质资源障碍。作为少数民族习用大宗药材以及民族中药现代化的重要品种,艾纳香的化学成分及其药理活性、提取工艺等是研究的热点领域[5],其次也涉及资源调查与遗传多样性[2,6]、种苗繁育栽培[7-8]、逆境生理[9-10]以及基因组和转录组等[11-12]。但有关艾纳香群落物种组成及其群落地位等特征信息一直未见报道,而这些信息对于通过群落特征掌握艾纳香自然生存状况、评估演替趋势具有极为重要的参考价值[13]。鉴于此,笔者于2020年8-9月期间,以罗甸县红水河地區为代表,对艾纳香群落进行了样地调查,并从植物区系、生活型、重要值及多样性等角度加以分析,以明确艾纳香现存典型植物群落基本特征,进而了解艾纳香在群落中的现状和演替趋势,以期为区域艾纳香资源的保护利用提供参考。
1材料与方法
1.1研究地概况
研究地位于罗甸县红水河地区(E 106.44°~106.75°,N 25.06°~25.28°),该地区位于云贵高原向广西丘陵过渡的边缘台阶处,属中低山河谷侵蚀地貌,河流切割深,地形破碎;地带性植被为湿润性常绿阔叶林,但由于二十世纪60~80年代持续地乱砍乱伐,大部分原生植被遭受破坏,现已演变为云南松、马尾松、桦、栎等为主的次生针阔混交林[3]。该地区在贵州气候区划上属南亚热带季风湿润气候区[14],年均气温20 ℃左右,年降雨量1300~1400 mm。
1.2样地设置
调查区域位于罗甸县红水河沿岸的红水河镇及云里村周边区域。通过社区访谈和实地初查发现,由于艾纳香种植业在该地区长期发展,当地群众有采集野生艾纳香无性繁殖幼苗作为种苗以及适度抚育野生群落的习惯,导致目前几乎已不存在纯自然状态的艾纳香群落。因此,调查样地设置于艾纳香野外逸生后自然繁殖发展基础上,近几年几乎处于荒废状态而仅受到轻度干扰的半人工、半自然状态群落。
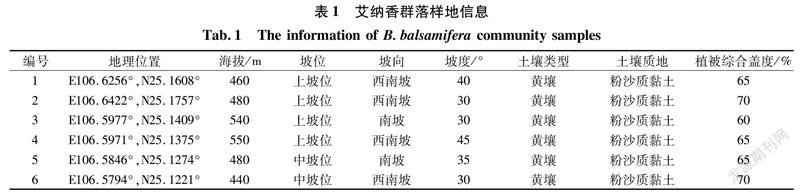
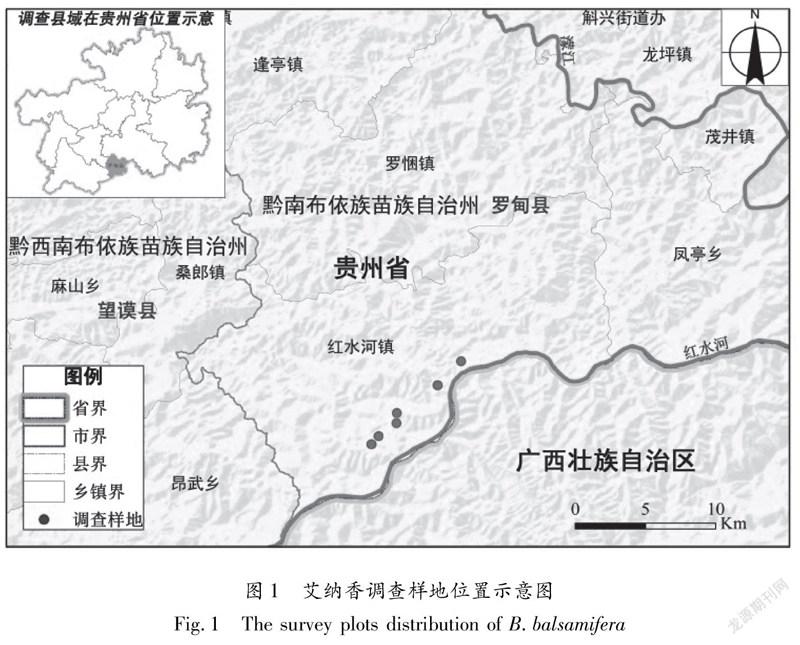
鉴于艾纳香现存典型群落不存在乔木层,因而在艾纳香种群集中分布且保存相对良好的地段内共设置6个20 m×20 m代表性样地(图1),记录样地海拔、坡度、坡向、土壤等情况(表1)。艾纳香群落各样地立地条件较为一致;海拔为440~550 m,这与牟珍等[2]2017年报道贵州艾纳香资源集中分布海拔情况基本一致。
1.3群落调查
将样地进一步划分为4个10 m×10 m样方,于样方四角和中央各设置一个2 m×2 m小样方和1 m×1 m小样方,总计2 m×2 m小样方和1 m×1 m小样方各120个。在2 m×2 m小样方中调查灌木层,1 m×1 m小样方中调查草本层,记录灌木层和草本层植物名称、高度、株数、盖度。由于艾纳香为多年生半灌木状大草本,本研究中调查时将高度大于2 m的艾纳香植株归入灌木层。
1.4数据处理与分析
植物区系及生活型谱:统计6个艾纳香群落样地植物科、属、种的组成,根据吴征镒[15]1991年提出的方案开展植物区系分析,确定群落组成植物属的分布区类型;根据Raunkiaer生活型分类系统明确群落植物生活型,建立群落生活型谱[13,16]。物种重要值:针对灌木层、草本层分别计算各植物重要值IV(%),其公式为:
IV=相对密度+相对频度+相对优势度/3(1)
式(1)中:相对密度=100×某种植物的密度/所有植物的密度之和,相对频度=100×某植物的频度/所有植物的频度之和,相对优势度=100×某植物的优势度/所有植物的优势度之和,其中:密度=样方内某种植物的个体数/样方面积,频度=某种植物出现的样方数/全部样方数,优势度=样方内某种植物的盖度[16]。
物种多样性指数:参照伊力塔等[17]、谢伟东等[18]的方法,运用物种丰富度指数(SA)、Shannon-Wiener多样性指数(H)、Pielou均匀度指数(J)以及Simpson优势度指数(D)测定群落物种多样性,其计算公式分别为:
SA=S(2)
H=-∑PilnPi(3)
J=(-∑PilnPi)/lnS(4)
D=∑Xi=1Pi2(5)
式(2)~式(5)中:S为群落植物物种数;Pi为物种i的相对重要值,即Pi=Ni/N,Ni为第i个物种的重要值,N为群落样地中所有植物重要值之和。
2结果与分析
2.1艾纳香群落物种组成及区系特征
群落物种组成是植物群落最基本特征,也是群落形成的基础[19]。统计表明,艾纳香群落共有64种植物,隶属于24科57属,群落植物灌草属性突出,无乔木层次。禾本科、菊科和豆科是艾纳香群落的优势植物科,植物种分别达到了14种、13种、8种。
植物区系分析有助于了解群落植物成分的特征与性质[20],而属比科更能反映植物的系统演化和地理特征[20]。由表2可知,艾纳香群落植物属涉及11个分布区类型,其中泛热带成分占首位,共14个属,占总属数的24.56%;其次为世界分布成分,共12个属,占总属数的21.05%;再次为旧世界热带、热带亚洲至热带非洲、旧世界温带成分,共占总属数的31.59%;其余分布区类型总计涉及13个属,共占总属数的22.81%。进一步统计表明,艾纳香群落共有30个属为偏热带性质的分布区类型,占全部植物属的比例约为53%。
2.2艾纳香群落物种生活型特征
生活型是植物对环境条件特别是气候条件长期适应而在生理、结构、尤其是外部形态上的一种具体反映[22]。由表3可知,艾纳香群落植物不存在地下芽生活型,群落生活型谱中高位芽植物总计30种,占群落植物的46.87%,优势较为突出,并且乔木性质突出的大高位芽、中高位芽、小高位芽植物达到了全部高位芽植物的46.67%;地上芽、地面芽和1年生植物总计34种,占群落植物的53.13%,其中地面芽植物相对居多。
2.3艾纳香群落物种重要值
物种重要值是群落的重要特征之一,在一定程度上反映群落物种动态及群落演替趋势[23]。群落灌木层中,艾纳香的重要值最大,达到了83.23%,其余植物重要值均低于2.00%(表4)。与灌木层情况类似,艾纳香在草本层中的重要值位列第2(26.68%),与位列第1的禾本科植物矛叶荩草(Arthraxon lanceolatus)重要值相當(27.37%),而位列第3的禾本科植物芒(Miscanthus sinensis)的重要值仅为5.99%(表5)。因此,艾纳香在群落中的地位极其重要,并占据绝对优势,为所在群落当前的建群种和单优势种。
2.4艾纳香群落物种多样性
物种多样性是生境中物种丰富情况及分布情况的一个量度[24],反映环境要素与群落植物生长的相互关系[25]。由表6可知,艾纳香群落1和群落4的物种丰富度指数SA较高,分别为25、27,而群落3最低(仅为10),群落间SA差异明显,变异系数达到了33.24%。群落1和群落4坡度相对更陡并且都处于上坡位(表1),进出相对困难,近两年人为干扰相对更少,这是该群落物种丰富度更高的重要因素。艾纳香各群落间Shannon-Wiener多样性指数H介于1.416~2.120,Pielou均匀度指数J为0.523~0.688,Simpson优势度指数D为0.260~0.433。总体上,H相对较高,J和D较低,H、J和D在群落间差异不大(变异系数10.76%~17.46%),并且均未同群落SA形成较好对应关系,表明由于受到建群种艾纳香的影响和控制,其他种群的分布总体不均、优势也不突出,这与调查的实际情况相符。
灌木层和草本层对比而言(表6),艾纳香群落灌木层H、J普遍小于草本层,而D却普遍大于草本层,主要由于灌木层物种数量(SA)明显少于草本层,单优势种艾纳香种群优势过于突出,导致整个灌木层分别反映多样性、均匀度为主的H、J低于草本层,而反映种群优势状况的D却偏高。此外,灌木层各多样性指数和群落整体水平差异较大,但草本层SA、H、J、D均较为接近群落整体水平,表明艾纳香群落物种多样性水平主要取决于草本层。
3结论与讨论
艾纳香群落由64种植物组成,隶属于24科57属,灌草属性突出,群落物种数量与同属亚热带的广西环江典型喀斯特峰丛洼地灌丛群落相当[26],但不到桂西地区灌丛群落的一半[27],也明显少于浙江天童地区灌丛群落[21],因此艾纳香群落物种组成并不丰富。同时,艾纳香调查群落分布集中,本底条件差异也较小,但单个群落物种数占全部群落物种数的比例较低(最高42.19%、最低仅15.63%),群落间物种组成差异大。此外,艾纳香群落组成有乔木树种,但不具备乔木层次。推测艾纳香群落表现出的上述情况同前几年艾纳香种植高峰期的人为干扰以及近两年逐渐荒废进而发生了一定程度自然恢复直接相关,这也与该地区存在适度抚育野生艾纳香群落的习惯相印证。
艾纳香群落植物以偏热带性质的区系成分为主(约占群落植物总属数53%),与本研究区邻近的红水河沟谷残存季雨林、上游地区北盘江石漠化灌草群落植物区系特征相似[14,28]。艾纳香群落植物生活型以高位芽和地面芽植物为主(占比78%),这与我国亚热带灌丛和地带性常绿阔叶林的植物生活型组成情况基本一致[21,29],并且明显区别于贵州典型暖温带地区草海保护区以地面芽为主的群落植物生活型谱[13]。研究认为,植物区系是植物与自然地理环境长期相互作用、相互影响的结果[20],我国亚热带地区高位芽植物和地面芽植物分布受温度和降水的共同影响[29]。因此,本研究中红水河河谷地区独特的南亚热带季风湿润气候正是艾纳香群落植物区系成分、生活型谱表现特征的气候背景。
艾纳香在群落中的重要值非常突出,在灌木层和草本层分别达到了83.23、26.68,占据群落绝对优势地位,为群落建群种和单优势种。研究表明,人为干扰可导致群落结构趋于简单,优势种重要值提高[26];而某种植物在群落中的重要值越大,其对群落稳定性的影响就越大,一旦遭到破坏,其群落也将不复存在或向另一类型群落演替[23]。因此,艾纳香的重要值表现与群落早期受到以收获艾纳香枝叶为导向的不同程度人为干扰密切相关,这也导致其群落外貌与研究区常绿性湿润阔叶林地带性植被不吻合[3],同时,持续的有利于艾纳香生存发展的人为干扰对于维持艾纳香在群落中的地位和群落现状稳定性至关重要。
作为灌草型单优势种群落,艾纳香各调查群落Shannon-Wiener多样性指数H、Pielou均匀度指数J、Simpson优势度指数D分别不超过2.120、0.688、0.433,其H、J水平与同处亚热带的广西地区及本研究区相邻上游地区的典型次生灌丛或灌草群落大致相当,但D水平明显偏低[26-28],表明艾纳香群落物种多样性水平总体较低。Simpson优势度指数侧重物种多度,其数值越大代表群落优势种越不明显[17],可以认为正是艾纳香作为单优势种的突出存在导致了群落物种多样性尤其是D水平偏低。因此,艾纳香群落物种多样性水平在现阶段虽然取决于草本层丰富程度,但人为干扰支撑的艾纳香种群发展对其又具有控制作用。
艾纳香属在植物区系上为旧世界热带分布(亚洲、非洲和大洋洲的热带地区及其邻近岛屿)[15],而艾纳香在我国仅见于长江以南的贵州、云南、广西、广东、福建、台湾[30],其热带属性较为突出;自然状态下通常分布于山坡草地、路边或灌木丛、疏林中[2,30],作为伴生种出现。本研究中艾纳香群落无乔木层次,但群落植物中存在较高比例的乔木类高位芽植物,如山槐(Albizia kalkora)、余甘子(Phyllanthus emblica)、珊瑚朴(Celtis julianae)、齿叶黄杞(Engelhardia serrata)和盐肤木(Rhus chinensis)等;同时,群落中还分布有入侵植物紫茎泽兰(Ageratina adenophora)。此外,前人研究还表明,艾纳香为典型阳性植物[31],极不耐阴[32]。因此,从群落演替角度而言,艾纳香群落具备向该地区地带性植被常绿阔叶林逐渐演替的气候、生境和物种库条件,自然状态下艾纳香在群落中的地位极有可能随着群落演替进程而逐渐被现有乔木类高位芽植物替代。研究表明,通过人为经营的持续干预,可以使得植物群落朝着预期的物种组成目标发展[33],进而影响植物群落演替进程[34]。因而,基于保护研究区艾纳香种质资源的角度,在今后对当地现存艾纳香群落实施有计划、科学合理的定向抚育管理,同时关注入侵植物紫茎泽兰的及时防除,有利于维持艾纳香在群落中的优势地位,从而达到保存更多艾纳香种质资源的目的。(责任编辑:胡吉凤)
参考文献:
[1]胡蕖,周家维.贵州省艾纳香植物资源现状及适生区区划初步研究[J].贵州林业科技,1999,27(1):44-48.
[2]牟珍,杨贤友,吴红梅,等.北盘江流域等地艾纳香植物资源调查研究[J].时珍国医国药,2017,28(4):981-983.
[3]江维克,周涛,何平,等.贵州红水河地区艾纳香植物资源调查及其保护策略[J].贵州农业科学,2010,38(8):1-4.
[4]韦政理.罗甸县艾纳香产业发展存在问题及对策[J].农技服务,2020,37(8):111-112.
[5]马海霞,杨广安,谭琪明,等.艾纳香化学成分及药理活性研究进展[J].化工管理,2021,4:69-70,72.
[6]肖永锋,黄梅,于福来,等.艾纳香种质资源表型性状遗传多样性分析[J].福建农业学报,2021,36(2):157-167.
[7]顾岑,王华磊,赵致,等.种植密度及采收期对苗药艾纳香产量和品质的影响[J].中药材,2016,39(2):235-239.
[8]黄梅,陈振夏,于福来,等.不同因素对艾纳香扦插生根的影响[J].贵州农业科学,2020,48(3):122-125.
[9]王俊丽,梁娟,任建国,等.土壤镉污染对艾纳香生长及体内镉积累和分布特征的影响[J].西南师范大学学报(自然科学版),2017,42(9):84-89.
[10]刘济明,邓明明,李丽霞,等.干旱胁迫对艾纳香盆栽幼苗生长及叶片结构的影响[J].云南农业大学学报(自然科学),2019,34(1):138-144.
[11]官玲亮,查英,白琳,等.艾纳香MYB转录因子家族生物信息学分析[J].热带作物学报,2020,41(7):1305-1312.
[12]譚永佳,陈静,张林,等.艾纳香BbCHS基因的克隆及其原核表达载体的构建[J].分子植物育种,2020,18(11):3598-3603.
[13]何斌,李青,冯图,等.草海国家级自然保护区森林群落特征研究[J].河南农业大学学报,2019,53(5):783-790.
[14]高焕晔,熊源新,苟光前.贵州红水河沟谷残存季雨林蕨类植物区系初步研究[J].浙江林学院学报,2008,25(1):48-54.
[15]吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991(2):1-139.
[16]宋永昌.植被生态学[M].上海:华东师范大学出版社,2002:99-167.
[17]伊力塔,韩海荣,豪树奇,等.灵空山辽东栋群落特征及其物种多样性[J].林业科学,2010,46(9):164-171.
[18]谢伟东,招礼军,邓荣艳,等.干扰对红鳞蒲桃群落植物多样性及重要值的影响[J].西南大学学报(自然科学版),2011,33(6):58-63.
[19]韓天丰,程积民,万惠娥.人工柠条灌丛林下草地植物群落特征研究[J].草地学报,2009,17(2):245-249.
[20]吴征镒,周浙昆,李德株,等.世界种子植物科的分布区类型系统[J].云南植物研究,2003,25(3):245-257.
[21]张光富,宋永昌.浙江天童灌丛群落的种类组成、结构及外貌特征[J].广西植物,2001,21(3):201-207.
[22]高贤明,陈灵芝.植物生活型分类系统的修订及中国暖温带森林植物生活型谱分析[J].植物学报,1998,40(6):553-559.
[23]谢春平,方彦,方炎明.乌冈栎群落垂直结构与重要值分析[J].安徽农业大学学报,2011,38(2):176-184.
[24]贺静雯,刘颖,李松阳,等.蒋家沟流域不同海拔灌草层群落特征与土壤关系的研究[J].广西植物,2020,40(8):1092-1100.
[25]张世雄,秦瑞敏,杨晓艳,等. 山西吕梁山草本群落物种多样性的海拔梯度格局及与环境因子的关系[J].广西植物,2020,40(12):1860-1868.
[26]张芳,杜虎,曾馥平,等.西南喀斯特峰丛洼地木本植物群落结构与多样性变化[J].生态学报,2020,40(12):4094-4104.
[27]申文辉,何琴飞,彭玉华,等.桂西不同灌丛植物群落物种组成及其多样性[J].广西植物,2016,36(10):1165-1171.
[28]陈龙,安明态,任娇娇,等.贵州北盘江石漠化区植物群落物种多样性及主要乔木种间联结性[J].西南大学学报(自然科学版),2020,42(5):80-94.
[29]李家湘,熊高明,徐文婷,等.中国亚热带灌丛植物生活型组成及其与水热因子的相关性[J].植物生态学报,2017,41(1):147-156.
[30]李永康,黄威廉,王兴国,等.贵州植物志(第九卷):种子植物[M].成都:四川民族出版社,1989:79-86.
[31]蔡荣,廖小锋,刘济明,等.药用植物艾纳香的光合日变化研究[J].贵州农业科学,2013,41(8):61-64.
[32]李丽霞,刘济明,池馨,等.艾纳香幼苗对夏季遮阴胁迫的生理响应[J].中药材,2016,39(8):1706-1709.
[33]臧丽鹏,张广奇,何跃军,等.近自然经营对黔中地区马尾松灌草群落多样性及物种共存的影响[J].山地农业生物学报,2021,40(3):21-28.
[34]盛茂银,熊康宁,崔高仰,等.贵州喀斯特石漠化地区植物多样性与土壤理化性质[J].生态学报,2015,35(2):434-448.
Study on the Community Characteristics of Medicinal Plant Blumea balsamifera in Guizhou Province Under Mild Human Disturbance
Tian Fan Li Peng Liao Xiaofeng
(1.Guizhou Forestry Academy of Sciences,Guiyang,Guizhou 550005,China;2.Guizhou Academy of Sciences,Guiyang,Guizhou 550001,China)
Abstract:In order to explore the basic characteristics and living conditions of Blumea balsamifera in community,these typical plant communities of B.balsamifera in the Hongshuihe area of Luodian county in Guizhou province were investigated by a sample method,and the community characteristics were analyzed from plant flora,life form,important value and species diversity.The results showed that there were 64 plant species in B.balsamifera community,belonging to 57 genera,24 families,mainly of Gramineae,Compositae and Legume.Among them,high bud and ground bud plants were the main species,and accounting respectively for 46.87% and 31.25% of the number of plant species.There was no tree layer in the community,the important value of B.balsamifera reached respectively 83.23% and 26.68% in shrub layer and herb layer.In the community,the species richness index of B.balsamifera was from 10 to 27,and the Shannon-Wiener diversity index was between 1.416 and 2.120,the Pielou evenness index was between 0.523 and 0.688,and the Simpson ecological dominance index ranged from 0.260 to 0.433.In general, due to the influence of human disturbance,B.balsamifera was the dominant specie and the only constructive specie in the community,and the level of species diversity which depended on the abundance of the herb layer was low.At the same time,the species composition,appearance characteristics,stability and succession syndynamic of the community were influenced greatly by the survival and development of B.balsamifera population.
Keywords:Guizhou;Blumea balsamifera;community characteristic;species diversity;important value
