目标树生长空间释放对云南松生长及其器官和土壤养分的影响1)
2022-05-16李俞鑫李莲芳合金鑫姜若超黄孝杰王姝茜刘畅
李俞鑫 李莲芳 合金鑫 姜若超 黄孝杰 王姝茜 刘畅
(西南林业大学,昆明,650224)
目标树是指林木特征满足经营目标的要求,响应于适宜的营林措施,并能在各种环境干扰中长期保持竞争力的优质林木[1-2]。目标树经营是一种通过降低相邻木的林冠竞争和释放目标树生长空间,从而提高单株林木生长和质量的营林技术,核心为“选择目标树、伐除干扰木或降低干扰木的影响、释放生长空间”,不同于常规的对全林采用相同的抚育措施,是一种围绕有益于目标树生长和质量提升的抚育经营方式[3-4]。Ward[5]研究发现,对美国红橡(Quercusrubra)、红枫(Acerrubrum)、黑橡(Quercusssp.)和黑桦(Betuladahurica)幼龄林等开展目标树经营4 a后,其胸径生长量分别较对照提高86%、56%、65%和52%;Ward[6]进一步研究指出,橡树人工林目标树经营25 a后,促进橡树胸径和上层林冠生长,提高大径阶优质林木占比。王懿祥等[7]研究表明,马尾松(Pinusmassoniana)人工林开展干扰树间伐试验3 a后,目标树的胸径和材积年均生长量分别高于对照的80%和55%。已有研究结果共同指出,目标树经营显著或极显著地影响单木胸径和材积生长,对树高生长的影响则尚无相似的结论[4,7]。
目标树经营以伐除干扰树来降低林木间的竞争,促进林内资源空间的合理配置,适宜的目标树经营措施显著促进林木生长,同时有效改善林下土壤理化性质[8]。柏木(Cupressusfunebris)[8]、马尾松[9]和杉木(Cunninghamialanceolata)[10]等树种开展目标树经营后,结果指出目标树抚育初期即促进土壤与林木间的物质循环和养分周转,为优质大径阶人工林培育提供促进林木生长的有利条件。目标树经营改善土壤理化性质,也改变植物器官对养分元素的吸收和利用,植物生长和发育过程与其不同器官的养分元素含量状况密切相关[11]。其中,叶片和根系内营养元素含量能反映目标树经营后的土壤养分变化;碳(C)是构成植物生物量的主要元素,氮(N)、磷(P)和钾(K)则是其生长必需的营养元素[12],这4种元素在植物体内的含量和生态化学计量特征是揭示林木对土壤养分利用状况的重要依据[13]。已有研究表明,目标树经营对林分土壤、叶片和根系的C、N、P和K具有显著影响,不同经营措施下土壤和林木器官养分呈现差异变化[14-17]。合理的目标树经营措施,有效改善林内光照和水热条件,从而增加土壤中的元素含量,林木根系大量吸收养分并输送至叶片,转化为有机物加速目标树生长,最大化的提高林分质量和产量。植物叶片和根系中C、N、P和K含量及其化学计量比的变化可以表征植物养分含量和养分间的限制性关系,以及不同植物对养分的需求和利用状况[12,18]。在森林培育中,应用生态化学计量学的方法,分析林木叶和根的生态化学计量关系,及其随森林经营措施导致的林木生长变化,可有效选择最佳抚育经营措施,更好的服务于目标树抚育经营实践[19]。
云南松(Pinusyunnanensis)是中国西南地区主要的用材树种之一,其分布区以山地为主,传统的全林抚育成本极高,在大径级木材培育中,目标树抚育经营技术,极大降低抚育成本的同时,促进林木生长,提升木材质量,是较为有效的抚育经营措施。目前,已有云南松林分抚育措施与林分生长[20]、化学计量比和针叶性状特征[21-23]、土壤理化性质及凋落物养分[24-26]等方面的研究,但将目标树经营措施对云南松生长、土壤和器官养分相结合的综合分析文献较少查及。基于此,本研究以16年生云南松人工林为研究对象,采用目标树经营的方法,运用目标树不同方向干扰树间伐的抚育方式释放生长空间,研究云南松人工林生长及器官与土壤养分对目标树空间释放的响应,分析最优的目标树生长空间释放措施,以期为滇中云南松目标树空间释放的抚育经营提供参考。
1 研究区概况
研究地位于云南省昆明市宜良县禄丰村国有林场尖山林区(24°33′30″~24°34′N,103°9′30″~103°10′E,图1),海拔1 820~1 970 m;属中亚热带季风气候,年平均、绝对最低和最高气温分别为17.5、-5.0和31.5 ℃,年降水量995.3 mm,蒸发量1 710.9 mm,年相对湿度68%,年平均霜期和日照时间67 d和2 032.8 h,偶有3~7 d的降雪期;土壤为酸性红壤,风化程度高[20,27]。林分为云南松纯林,林中少许混交马尾松、旱冬瓜(Alnusnepalensis)和华山松(Pinusarmandii)等树种;林下灌木主要是滇石栎(Lithocarpusdealbatusrehd)、棠梨刺(Pyruspashia)和南烛(Vacciniumbracteatum)等,草本主要有芒萁(Dicranopterisdichotoma)和黄茅(Heteropogoncontortus)等,地表为大量半分解或未分解针叶凋落物,腐殖质层约3~5 cm。
云南松人工林于2004年采用百日苗造林,林龄16 a,为人工纯林,造林面积约8 hm2。试验地位于东北坡的中上部,坡度5°~15°;林分初植密度3 300株·hm-2,经疏伐,目标树抚育时保留密度为1 945株·hm-2,郁闭度0.6~0.7。试验地面积1.0 hm2,林木冠形生长良好,干形通直。
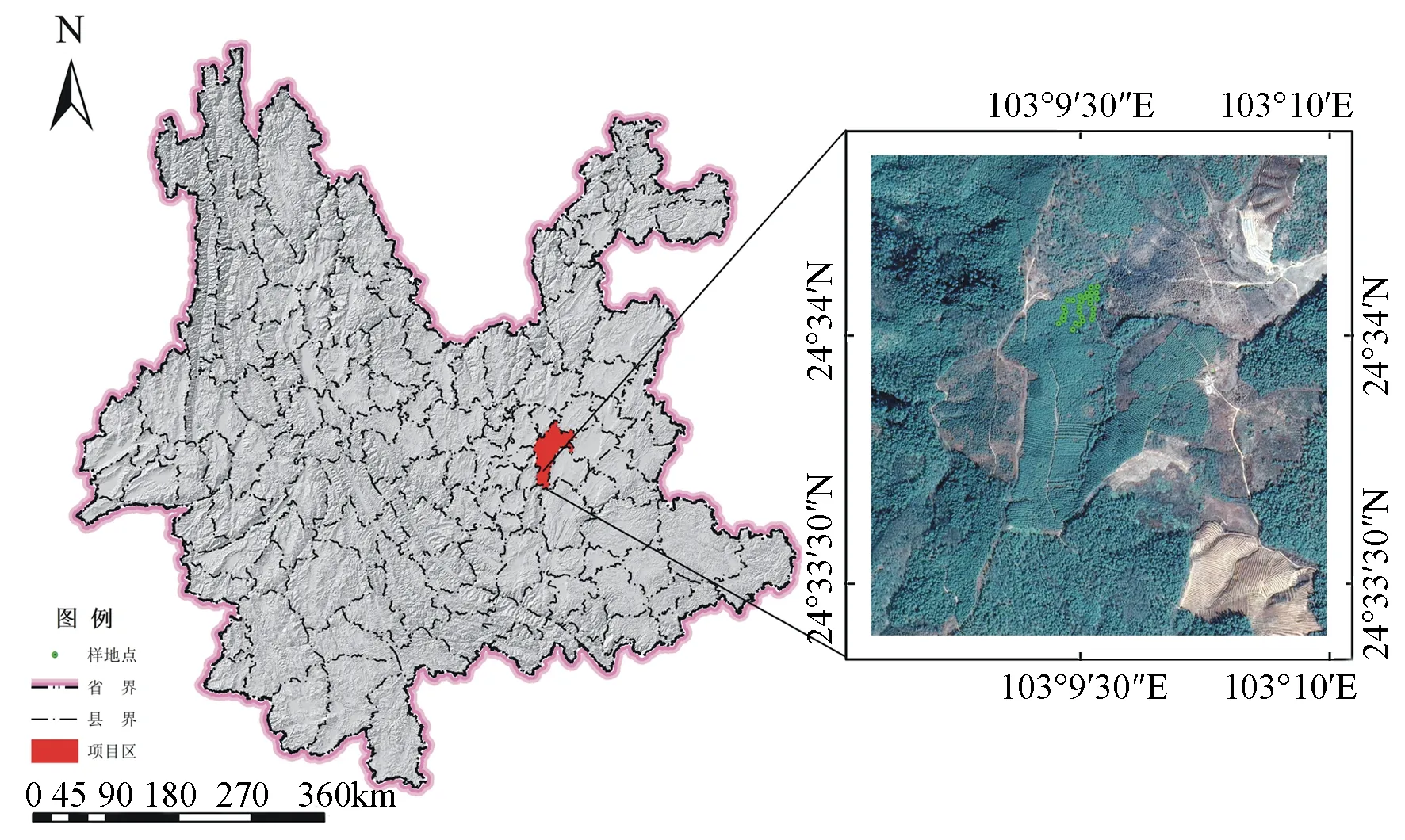
图1 研究地位置图
2 研究方法
于2020年5月在试验地内选择立地条件和林木生长情况基本一致的地段设置样地,共5块面积均为0.1 hm2(50 m×20 m)的样地,其中4块为目标树空间释放样地,1块为对照样地。目标树选择方法参考王懿祥、吕倩和杨历雨等[7-9,20]的方法,综合文献的基础上,在样地内选择生长旺盛、树干通直圆满、健康无病虫害、树高高于平均高的林木作为目标树,开展空间释放的抚育。为防止目标树间相互干扰,相邻目标树间的距离大于5 m,用红色油漆和铝牌进行永久性标记。
目标树生长空间释放方法参考Lamson et al.[27]的“树冠重叠”释放作业技术,该技术以目标树为中心,用虚拟的2条相互垂直的线段将目标树的树冠划分为4个象限,形成一个平面坐标系,试验中如果伐除释放生长空间N个象限周围的相邻木,表示目标树经过N个方向的释放,并获得N个方向的自由生长空间(FTG)[3-4],记为FTG-N(N=0、1、2、3、4)。FTG-0表示目标树4个象限的相邻木均未伐除,即为对照(CK);FTG-l、FTG-2、FTG-3和FTG-4分别表示4个方向的林冠释放(图2)。
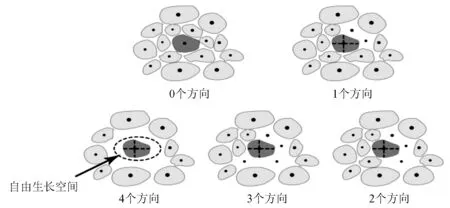
图2 林冠重叠释放后目标树树冠投影图
采用单因素设计(包括对照)进行目标树生长空间释放。在编号为1~5的样地中,1~4号样地分别进行4个方向的干扰树间伐,5号样地只选出目标树,不做处理,为对照样地,试验构成5个处理,单株小区,10次重复,共50株林木。按照目标树要求分别于相应的样地内选择目标树后,根据目标树生长空间释放设计,对目标树周边的干扰木进行伐除,充分释放目标树的生长空间,为其生长提供条件。
2.1 生长调查与样品采集
试验地内,2021年6月于抚育后1 a时,对试验所有目标树进行一一对应的每木检尺(胸径和树高);同时,每处理随机选择3株目标树,在目标树冠幅下,以及目标树周边林窗下各设置3个土壤采样点,清理枯落物及黑土层(消除腐殖质土对养分的影响)后,开挖土壤剖面,分别在0~20和20~40 cm土壤层,采用环刀取土样称质量测定密度,3次重复,相同土层土壤组成混合土样,用四分法取混合土样装袋,共计10个土样[8-11,24-28]。采集同样的目标树,每株林木从顶梢往下的第5轮枝条,在林木南面(阳面)中上部采集3条小枝,获取针叶,混合为一个样品,每一处理3个样品,5个处理共计15个针叶样品;同时,与针叶对应的方向挖取目标树侧根,采集细根作为根系样品,与针叶样品对应按编号分装到不同自封袋中,带回实验室测定相关指标。
2.2 样品测定与统计分析
采集的土壤样品经剔除杂质后分为两份,一份称质量后于烘箱中80 ℃烘至恒质量,计算土壤含水率。另一份自然风干,研磨后过0.25 mm孔径筛,用于pH值、有机碳和养分测定。针叶样品于105 ℃温度杀青15 min后,在65 ℃条件下烘干,粉碎过100目筛,装袋封存。根系样品用蒸馏水洗净后,于烘箱中65 ℃烘至恒质量,磨碎过100目筛备用。土壤和植物样品均以浓H2SO4-H2O2消煮后测定相关指标。土壤密度采用环刀法测定;烘干法测定土壤含水率;pH计测定土壤pH值;重铬酸钾氧化—外加热法测定土壤、针叶和根系有机碳(SOC);全氮(TN)、全磷(TP)和全钾(TK)分别采用凯氏定氮法、钼锑抗比色法和火焰光度法测定;土壤有效氮(AN)采用碱解扩散法,有效磷(AP)采用酸溶—钼锑抗比色法,土壤有效钾(AK)采用火焰光度法[8,25,28]。
云南松人工林单株立木材积计算公式:V=0.000 087 151 050 0D1.954 479 3H0.755 839 523(式中:V为单株立木材积;D为胸径;H为树高)[20]。土壤和针叶相关指标采用国标《森林土壤分析方法》计算为标准数据[29];应用Excel2010和SPSS20.0统计分析软件对数据进行处理、统计分析和绘图,单因素方差分析。
3 结果与分析
3.1 生长空间释放对目标树生长的影响
试验前,云南松目标树的胸径、树高和单株材积分别为16.6~17.3 cm、8.6~9.3 m和0.111~0.121 m3·株-1,对照为16.9 cm、9.6 m和0.120 m3·株-1;试验1 a后,试验4种处理的以上3项指标提高至18.1~18.4 cm、8.9~9.6 m和0.136~0.145 m3·株-1,对照则为17.4 cm、10.4 m和0.135 m3·株-1;其胸径、树高及材积增长率分别达5.5%~10.4%、2.7%~7.7%和17.4%~23.9%(对照为3.0%、8.2%和12.5%)。试验处理的胸径和材积增长率极显著高于对照的,同时,FTG-3和FTG-4的此两项指标极显著高于FTG-1和FTG-2(P=3.79×10-20~3.33×10-12<0.01),即生长空间释放方向越多,越有益于胸径生长;树高增长率则相反,FTG-1、FTG-2和对照的增长率高于FTG-3和FTG-4(P=2.10×10-9<0.01),即林冠受压的目标树,其树高生长高于被释放的(表1)。
3.2 生长空间释放对林下土壤密度和含水率的影响
试验实施1 a后,土壤0~20和20~40 cm的密度分别为1.70~1.86和1.80~1.90 g·cm-3,对照的为1.60和1.76 g·cm-3,试验处理0~20 cm土壤层的密度显著大于对照的;FTG-1和FTG-2的显著大于FTG-3和FTG-4(P=0.049<0.05),随着生长空间释放方向的增加,土壤密度随之降低;与0~20 cm土壤层的不同,处理间20~40 cm土壤层的密度无显著差异(P=0.282>0.05),即两个土壤层次的密度受空间释放影响的结果不同(表2)。

表2 目标树林下土壤密度和含水率对空间释放的响应
表2显示,0~20和20~40 cm的土壤含水率分别为6.73%~8.23%和11.29%~15.75%,对照的则为12.61%和19.06%;试验的所有处理0~20 cm土壤层含水率极显著地低于对照的(P=0.002<0.01);20~40 cm土壤层含水率则是FTG-1、FTG-2和FTG-3的极显著低于FTG-4和对照的(P=3.65×10-4<0.01),结合土壤密度,也许FTG-4的植被丰富度较高,土壤通透性较好,降水部分深入下层提高其含水率,对照含水率较高的原因则与0~20 cm土壤层的相同;试验样地0~20 cm土壤层含水率低于20~40 cm的,主要是20~40 cm土壤层受光照的影响较0~20 cm土壤层小造成。
3.3 生长空间释放对林下土壤化学性质的影响
表3显示,试验实施1 a后,试验处理的0~20 cm土壤层pH值极显著地低于对照的(P=2.11×10-5<0.01),FTG-4的土壤层pH值极显著低于其余处理(pH为3.83),土壤表层pH值随生长空间释放方向的增加而降低,即土壤趋于酸化,这也许与林下水热条件的改善后,微生物加速分解枯落物产生有机酸有关,有待进一步试验研究;0~20 cm土壤层有效磷和土壤有效钾随生长空间释放方向的变化规律与pH值一致,不同的是,FTG-3和FTG-4的有效磷极显著地低于其余处理和对照的(P=0.001<0.01),土壤中有效磷和土壤有效钾可被植物直接吸收利用导致的,目标树的生长证实此结论;0~20 cm土壤层有效钾无显著差异(P=0.115>0.05),这与土壤中K质量分数较高有关。试验处理的0~20 cm土壤层根系有机碳、全氮、全磷和全钾质量分数均极显著地高于对照样(P=(7.06×10-7~0.008)<0.01),随生长空间释放方向的增加,以上指标呈现逐渐增加的变化规律;处理间,FTG-1的根系有机碳质量分数极显著地低于其他的;全磷随着空间释放方向的增加而增加,FTG-3和FTG-4的土壤层全磷质量分数极显著地高于FTG-1和FTG-2;0~20 cm土壤层有效氮为127.65~171.07 mg·kg-1,FTG-1极显著地低于其他的处理(表3);试验实施后,林下土壤凋落物增多,光热条件改善,微生物活跃,其分解作用导致根系有机碳、全氮、全磷和全钾质量分数增加。
在20~40 cm土层中,FTG-3和FTG-4的土壤层pH值极显著低于FTG-1、FTG-2和对照(P=1.75×10-4<0.01);对照的有效磷质量分数极显著地高于试验处理且在FTG-4达到最小值,FTG-3和FTG-4极显著小于FTG-1和FTG-2(P=4.61×10-7<0.01),有效钾质量分数与0~20 cm土壤层的一致;随生长空间释放方向的增加,试验处理0~40 cm土壤层的pH、有效磷和有效钾均呈下降趋势,这与0~20 cm土壤层的变化规律相同;处理间,FTG-4的土壤根系有机碳(17.26 g·kg-1)、全氮(1.47 g·kg-1)、全磷(0.54 g·kg-1)、全钾(25.75 g·kg-1)和有效氮(81.14 mg·kg-1)均为最高,其次是FTG-3,除有效氮外,以上4个指标均极显著地高于对照的(P=(2.70×10-7~0.002)<0.01);FTG-3和FTG-4的土壤层根系有机碳和全磷极显著地高于FTG-1和FTG-2;FTG-4的全钾和有效氮极显著地高于FTG-1和FTG-2;与0~20 cm土层相同,20~40 cm土层的上述5个指标均随生长空间释放方向的增加而升高。不同土层间,除土壤pH值外,0~20 cm土壤层的根系有机碳、全氮、全磷、全钾、有效氮、有效磷和有效钾均高于20~40 cm土层的各指标(表3),这与干扰树间伐后枯落物集中于地表,地表土壤营养物质积累增加有关。
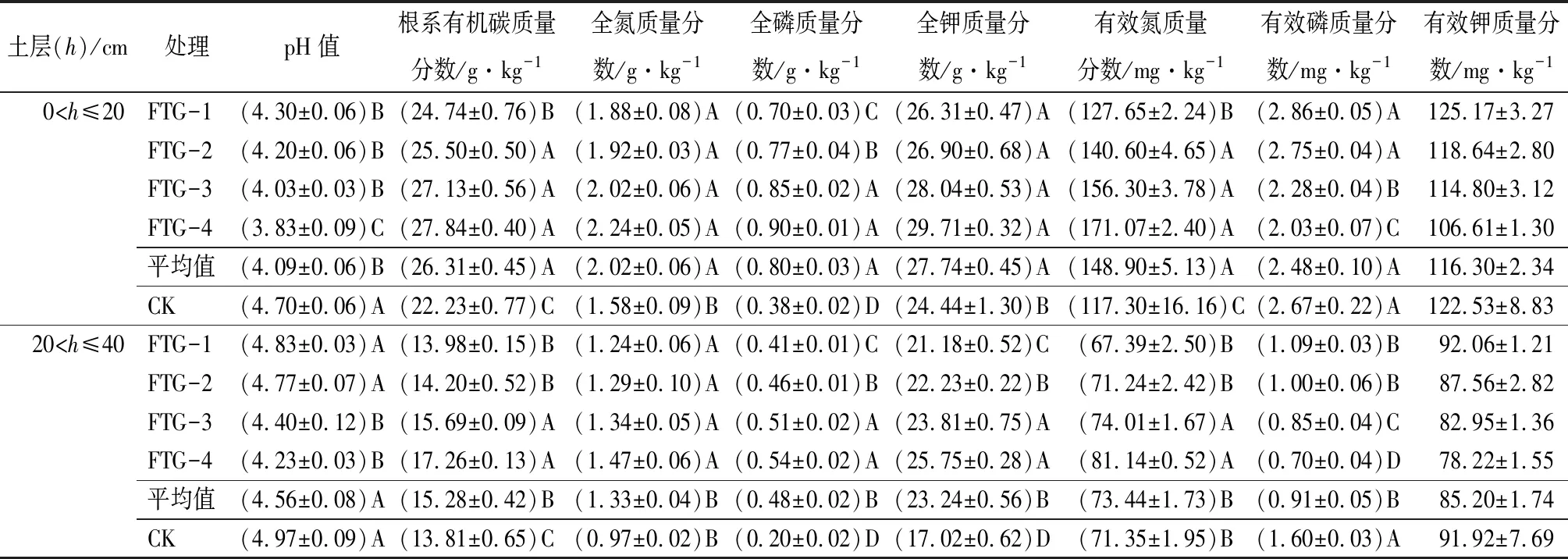
表3 自由生长空间对云南松土壤化学性质的影响
3.4 目标树叶和根养分质量分数对生长空间释放的响应
试验实施1 a后,云南松针叶中C、N、P和K质量分数分别为434.15~462.87、14.72~17.25、2.64~3.42和8.62~9.33 g·kg-1(对照的431.71、13.83、2.58和7.62 g·kg-1);根的以上4个指标分别为428.39~449.74、8.25~9.46、0.74~1.16和3.09~3.60 g·kg-1(对照的435.73、8.10、0.84和3.01 g·kg-1);与土壤的一致,FTG-3和FTG-4的针叶C质量分数极显著地高于FTG-1、FTG-2和对照(P=0.004<0.01),随生长空间释放方向的增加,针叶和根系中C质量分数逐渐增大,试验处理的针叶N质量分数显著高于对照(P=0.015<0.05);针叶和根系N和P质量分数随生长空间释放方向的关系与土壤的相同;试验处理针叶K质量分数极显著地高于对照(P=0.002<0.01),并随生长空间的增加而上升,根则是FTG-4的极显著高于其余处理和对照(P=0.004<0.01);生长空间释放方向的增加,叶和根中C、N、P和K质量分数上升,且针叶的C、N、P和K质量分数均高于根系(表4)。

表4 不同自由生长空间云南松器官C、N、P和K质量分数 g·kg-1
3.5 生长空间释放的目标树针叶和根中C、N、P、K的生态化学计量特征
试验实施1 a后,云南松针叶w(C)∶w(N)、w(C)∶w(P)、w(C)∶w(K)和w(N)∶w(P)分别为26.89~29.51、135.54~164.90、49.37~50.42和5.06~5.69,对照的为31.25、167.70、56.85和5.37;根的以上4个指标分别为47.60~51.93、389.41~582.05、125.16~138.87和8.19~11.20,对照的则为53.84、522.98、144.87和9.69;FTG-4的针叶w(C)∶w(P)极显著低于其他3项(P=0.003<0.01;比对照低24%),根的w(C)∶w(P)显著低于其他处理和对照(P=0.03<0.05;低于对照34%);试验处理的针叶w(C)∶w(K)极显著低于对照(P=0.002<0.01);FTG-3和FTG-4的根w(C)∶w(K)显著低于FTG-1、FTG-2和对照(P=0.024<0.05);针叶和根系的w(C)∶w(N)、w(C)∶w(P)、w(C)∶w(K)和w(N)∶w(P)随生长空间释放方向的增加而降低,针叶的以上4个指标均低于根系(表5),器官w(C)∶w(N)、w(C)∶w(P)和w(C)∶w(K)比值越低,表明其N、P和K质量分数较高,w(N)∶w(P)则与林木生长速率有关。

表5 不同自由生长空间云南松器官C、N、P和K生态化学计量特征
4 讨论
4.1 云南松目标树生长对生长空间释放的响应
合理的抚育间伐增加林木的生长空间,有效促进保留木的快速生长[30]。在云南松人工林中开展目标树空间释放抚育1 a的较短时间内,目标树胸径和材积增长率极显著地大于对照,表明间伐干扰树后,短期内即反应于目标树生长,这与有关干扰树间伐对人工林影响的研究结果基本一致[1-7,30-31]。试验实施1 a后,4个方向生长空间释放的树高增长率仅为2.7%,为对照的1/3,释放生长空间的林木树高增长率低于未释放的,说明自由生长空间越大,冠幅受压程度越小,在短期内主要以促进胸径生长为主,这与橡树[27]和黑樱桃(Cerasusmaximowiczii)[32]人工林的目标树抚育生长效应相一致;16年生云南松处于林分速生时期,释放生长空间后,目标树周围出现较大林隙,林冠由垂直扩张转为横向生长,这也许与有益于胸径生长有关。然而,有研究指出,目标树抚育经营提高林分树高生长量[33]或无显著影响[7],目前关于目标树抚育后,树高生长对干扰树间伐消除竞争的响应还存在争议,还需要大量的试验研究获取规律性的结论。本研究中,目标树胸径和材积增长率随空间释放方向的增加而增大,FTG-3和FTG-4的目标树胸径和材积增长率极显著高于FTG-1、FTG-2和对照,这与马尾松[7]、蒙古栎(Quercusmongolica)和水曲柳(Fraxinusm)[34]等的研究结果相一致,也表明中龄林云南松消除4个方向的干扰树影响对林木生长效果最优,揭示不同树种目标树对自由生长空间释放的响应具有共性,即干扰树竞争的消除,增加目标树的自由生长空间,为其生长提供更多资源,促进有机物的积累,加速林木生长。
4.2 生长空间释放对云南松林下土壤理化性质的影响
目标树通过抚育释放自由生长空间,促进林木生长的同时,也引起林下土壤理化性质的变化[9]。本研究中,试验实施1 a后,0~20 cm土壤层的密度随生长空间释放方向的增加而降低,对照的则低于目标树抚育的,0~20和20~40 cm土壤层密度对不同方向空间释放的响应不同。生长空间释放方向的增加,有效改善林内光照和水热条件,林下植被丰富度提高,微生物活跃,分布于土壤0~20 cm层次的植物根系较多,改善土壤结构,形成土层密度在不同土层和不同空间释放方向下的差异;与对照样地相比,也许是目标树抚育样地采伐过程中的人为活动造成的,导致试验样地0~20 cm土壤层密度较高。与土壤密度不同,释放生长空间的土壤层含水率小于未释放的,消除干扰树的竞争降低林分郁闭度,地表光照的增强加快水分的蒸发,而且研究区域处于雨季初期,降雨量较少,遮荫度较高的对照样地,蒸发较少,有利于土壤表层水分保持,故其含水率高于目标树空间释放的,此结果与吕倩等[8]对柏木目标树的研究结果相一致。试验样地0~20和20~40 cm土层的含水率随生长空间释放方向的响应不同,一方面,与20~40 cm土层位于土壤下层,光照对其影响较小,水分蒸发量较低有关;另一方面,4个方向的空间释放条件下,林下植被较为丰富,其生理活动导致土壤通透性提高,水分通过土壤孔隙进入下层,引起FTG-4的20~40 cm土壤层含水率升高。由于试验开展目标树经营时间较短,土壤密度和含水率随干扰树竞争消除出现不同变化,土壤含水率变化与马尾松[9]和杉木[10]的研究结果相一致,而土壤密度与之相反,也许是抚育方式不同造成,具体原因有待进一步试验研究。
试验实施1 a后,目标树空间释放的土壤pH值极显著低于未抚育的,吕倩等[9]对马尾松目标树抚育的结果认为,消除干扰树竞争后,林地采伐剩余物和枯落物增多,其分解时产生的有机酸导致根际土壤pH降低,极大促进林木生长的同时土壤酸碱性也有所变化,本研究结果与其相一致,形成原因也相同。土壤的有效磷和有效钾随空间释放方向的增加而降低,与李旭等[35]对落叶松(Larixolgensis)、云杉(Piceagezoensis)和冷杉(Abiesnephrolepis)林间伐后土壤有效磷和有效钾的变化相一致,主要是林木生长对此2类养分有效吸收引起的。本研究中,不同方向的空间释放对土壤根系有机碳、全氮、有效氮、全磷和全钾质量分数均有影响,并随空间释放方向的增加而提高,土壤根系有机碳和N主要来源于土壤有机质,而土壤有机质、全磷和全钾与采伐剩余物和枯落物分解有关,空间释放有效缓解植物对资源空间的激烈竞争,林下植被丰富度提高,土壤微生物活动活跃,加速枯落物及采伐剩余物的分解与有机质的矿化作用,导致土壤以上养分质量分数的提高。此外,云南松林下土壤表现为“缺P少N富K”的特征[24],不同生长空间释放的目标树抚育引起局部生境及林分特性的变化,导致土壤养分多样性的变化。
4.3 生长空间释放对云南松器官养分质量分数及生态化学计量特征的影响
生长空间释放改变林分的立地环境,加速林地凋落物和采伐剩余物的分解,土壤养分质量分数提高,林木从土壤中吸收养分,不同器官的养分元素分配格局产生变化[15,18]。本研究中,随生长空间的增加,针叶和根系中C、N、P和K质量分数积累逐渐升高,16年生云南松人工林处于生长旺盛时期,干扰树间伐或修枝释放生长空间,林木生长加速,干物质和含碳的结构性物质积累增加,促进针叶和根系C质量分数提高;刘万德等[36]和曾德慧等[37]指出,为支撑林木快速生长,核糖体需要快速地合成蛋白质和核酸,蛋白质的合成则需要较多的rRNA,从而导致N质量分数升高,rRNA又是植物的一个主要的P库,生长速率的加快导致分配到rRNA中P质量分数的增加,因此,导致植物器官C、N和P质量分数升高,本研究与此规律相一致。盘金文等[38]指出,植物体内K元素主要与植物抗性有关,试验实施1 a后,小生境的变化引起目标树适当增加K元素的吸收,以适应环境的变化,即K元素在器官中的变化,也许是目标树适应抚育措施导致环境变化的应急响应。FTG-4的目标树生长量最大,针叶和根系C、N、P和K质量分数均为最高,揭示4个方向的完整林冠释放对林木生长极为有效。本研究中,目标树针叶的C、N、P和K质量分数均高于根系,这主要是叶片作为植物进行光合作用和利用营养元素的主要器官,需要大量营养元素合成有机物以支撑林木快速生长导致的,这与盘金文等[38]的研究结果相一致,揭示林木抚育后,C、N、P和K的变化具有共同特性;同时,间伐目标树周围干扰树有效改善林地环境,土壤养分增加,林木加速吸收利用养分,针叶和根系中养分明显提高;针叶与根系养分差异是其结构和功能不同导致的,也表明不同器官对营养元素的吸收利用具有特异性。
贺金生等[39]和刘万德等[36]指出,C、N、P和K生态化学计量比是生态系统过程及其功能的重要表征,植物不同器官w(C)∶w(N)、w(C)∶w(P)和w(C)∶w(K)表征植物吸收营养过程对碳的同化能力,在一定程度上反映植物的营养利用效率及植物的生长速率。本研究中,不同生长空间释放方向针叶和根系的w(C)∶w(N)无显著差异,说明在相同的N元素条件下,生长空间释放对云南松器官碳固定效率影响不显著,这与马尾松[13]和侧柏(Platycladusorientalis)[38]人工林的研究结果类似。崔宁洁等[18]研究指出,植物w(C)∶w(N)、w(C)∶w(P)和w(C)∶w(K)比值较高时,养分供应条件受限,林木生长缓慢,为提高养分的利用率,需要保持较高的C与N、P和K质量分数比值以增强对养分胁迫环境的适应,即林木的生长速率较高时,其w(C)∶w(N)和w(C)∶w(P)较低,本研究FTG-4的以上4个指标均为最低,林木生长速率最快的结果与此结论相一致,揭示4个方向的空间释放对目标树生长最为有利。封焕英等[40]研究指出,N、P和K质量分数高低决定w(C)∶w(N)、w(C)∶w(P)和w(C)∶w(K)的高低,本研究试验实施1 a后,快速生长的林木从土壤中吸收利用的N和P积累增加,导致云南松针叶和根系的w(C)∶w(N)、w(C)∶w(P)和w(C)∶w(K)随生长空间释放方向增加而降低的变化规律,与前述文献相一致,揭示森林抚育中林木器官养分变化具有共同特性。本研究云南松根系的C与N、P和K、N与P质量分数比值均高于针叶,原因是针叶为光合作用和营养元素利用的主要器官,根则主要负责营养物质的吸收和运输,结构和功能的不同造成云南松针叶和根系的生态化学计量比的差异。
刘俊雁等[21]和杨卫等[25]研究指出,植物叶片的w(N)∶w(P)是判断环境对植物生长养分供应状况的重要指标,反映土壤养分供给能力的大小,当叶片w(N)∶w(P)低于14时,林木生长主要受N限制;本研究中,生长空间释放和未释放的云南松针叶和根系的w(N)∶w(P)均低于14,表明研究的目标树生长主要受到N元素的限制,这与上述文献对不同地区云南松的研究结果相同,揭示云南松林分受N养分限制的共性,也表明在云南松不同器官间N和P质量分数及w(N)∶w(P)具有一致关联性。本研究试验实施1 a后,处理间云南松针叶和根系的w(N)∶w(P)虽无显著差异,但其化学计量比随生长空间释放方向增加而降低。Elser et al.[41]和Virginia et al.[42]在生长速率假说理论中指出,生物体必须改变w(C)∶w(N)∶w(P)比值适应生长速率的改变,高生长速率对应较低的w(C)∶w(N)、w(C)∶w(P)和w(N)∶w(P)比值[43];本研究的目标树生长与器官生态化学计量比随生长空间释放的变化与上述理论相一致,表明目标树开展生长空间释放的抚育措施后,有效改善林木器官养分质量分数及其化学计量比,林木生长速率加快,即云南松人工林中龄林阶段开展目标树的生长空间释放,可有效促进林木生长,其中4个方向生长空间全面释放对林木生长最为有效的结论,通过生长速率假说和生态化学计量学得到充分的理论解释。
5 结论
云南松中龄林目标树的生长空间释放抚育,在抚育实施后1 a的短期内极显著地促进其胸径和材积生长(P<0.01),并且随生长空间释放方向的增加,云南松林下土壤理化性质、器官(针叶和根系)养分质量分数及其生态化学计量比均有明显改善。云南松人工中林龄的土壤N和P元素质量分数较低,目标树生长主要受N养分限制。综合目标树生长空间释放抚育1a后的林木生长、林下土壤理化性质、目标树器官养分质量分数及其生态化学计量比随生长空间释放方向的变化,开展4个方向的全面生长空间释放,是云南松中龄林目标树空间释放抚育中最有效的措施,可应用于生产实践。
