钙离子对不同水稻品种苗期镉吸收转运的影响
2022-05-16付天岭普天赟高珍冉罗德强何腾兵
滕 浪, 付天岭, 郑 锋, 普天赟, 高珍冉, 秦 松, 罗德强, 何腾兵,
(1.铜仁市农业农村局, 贵州 铜仁 554300; 2.贵州大学农学院, 贵阳 550025;3.贵州大学新农村发展研究院, 贵阳 550025; 4.贵州省农业科学院土壤肥料研究所, 贵阳 550025;5.贵州省农业科学院水稻研究所, 贵阳 550025)
镉(Cd)是稻田土壤主要的重金属污染元素,水稻对Cd具有很强的吸收能力,农田土壤中主要是以Ca2+的形态通过水稻根系进入水稻体内积累,对水稻产生毒害作用[1]。一般来说,水稻对Cd的解毒与耐受机制有2个基本途径,一是解毒排斥机制,即避免过量Cd进入水稻组织细胞,或者阻碍Cd在水稻体内的运输;另一途径是内部耐受机制,即Cd结合到细胞壁上、主动运输进入液泡或与某有机酸和蛋白质结合等[2]。水稻各品种对Cd吸附能力有显著差异[3-4],呈现出籼型杂交稻>粳型杂交稻>常规籼稻>常规粳稻的规律,各部位Cd积累量由高到低顺序为:根系、茎叶、穗轴、籽粒[5-6],水稻对Cd的吸收能力差异性取决于各水稻品种的基因型差异。钙是植物生长发育必需的营养元素之一,参与细胞壁的结构合成,并且还能与磷脂分子联结起来,稳定细胞膜结构[7]。Ca2+与Cb2+具有半径相近电荷相同的特点,能够相互形成竞争离子,共同竞争植物根表面的吸附位点与离子通道,从而降低植物对Cd的吸收[8]。植物抗胁迫能力与植物体内的抗氧化酶活性息息相关,研究表明,添加外源Ca2+能够使水稻植物体内抗氧化酶及抗氧化物质的活性提高,减少丙二醛与自由基含量,但是对植物耐Cd性机理尚不明确[9]。已有大量研究表明,在稻田土壤Cd污染治理中,施用含Ca材料的调理剂与肥料,能够降低水稻根系对Cd的吸收,可有效地降低水稻籽粒中Cd的含量,保障粮食安全[10-11],但是未明确指出是材料中Ca降低了水稻对Cd的吸收。
贵州属于Cd的地球化学背景异常区,土壤中Cd平均含量为0.659 mg/kg[12],同时,贵州也是西南典型的喀斯特地区,土壤与水中富含Ca2+、Mg2+、CO32-/HCO3-、SO42-等,其中Ca2+含量占阳离子总量的40%~80%[13]。已有研究表明,在喀斯特地区中性土壤Cd的有效态占比较低,从而降低水稻对Cd的吸收[14]。田茂苑[15]研究表明,在贵州典型喀斯特地区石灰性水稻土中Cd风险程度较高,而稻米风险较低,在黄壤性与红壤性水稻土中土壤与稻米风险格局呈现一致性。但上述研究均未指明喀斯特地区土壤某种物质对水稻Cd积累有影响,因此本研究根据喀斯特地区土壤与稻田灌溉水中Ca2+含量,探讨Ca2+能否降低水稻苗期对Cd的吸收。采用本地水稻品种红优2号、黑糯72与杂交品种C两优华占、宜香优2115相对比,通过营养液短期培养的方法,研究不同Cd污染程度下添加不同浓度Ca2+对水稻苗期根系Cd吸收及转运的影响,同时为非喀斯特地区施用含Ca材料修复治理不同程度Cd污染稻田土壤提供理论依据,对筛选低富集水稻品种具有重要意义。
1 材料与方法
1.1 供试材料
供试水稻品种选择籼稻:红优2号、宜香优2115、C两优华占;粳稻:黑糯72,其中红优2号与黑糯72为贵州地方品种,C两优华占(C 815 S×华占)为两系杂交品种,宜香优2115(宜香1 A×雅恢2115)为三系杂交品种。水稻种子与幼苗均由贵州省农业科学院水稻研究所提供。
1.2 试验方法
试验于2019年6月12—19日进行,采用1/4水培营养液培养,营养液由硝酸铵、二水磷酸氢钠、硫酸钾等物质混合配置而成[16]。处理所用试剂为分析纯氯化镉(CdCl2)及氯化钙(CaCl2)。水培所用容器为1 000 mL棕色广口瓶,瓶口用定植棉固定。选择长势一致的3叶1心的水稻秧苗,用蒸馏水清洗根系后移栽至1 L营养液中,4株/瓶,每2 d用NaOH与HCl调节pH值保持在5.75,设置2个Cd2+浓度水平:1.0、2.0 mg/L(简称为:Cd 1、Cd 2),4个Ca2+浓度0、0.5、1.0、2.0 g/L(简称为:Ca 0、Ca 1、Ca 2、Ca 3),在2个Cd2+水平下添加4种不同浓度Ca2+培养,1个空白对照(Cd 0 Ca 0),共计9个处理,每个处理重复3次。每60 h更换一次营养液,每次更换营养液添加相应Cd2+与Ca2+处理,共计换3次营养液,培养7 d。
1.3 样品处理及测定
将水稻根系用0.02 mol/L的Na2EDTA溶液浸泡15 min,除去表面Cd2+与Ca2+,用超纯水冲洗水稻根系3次,吸干水分,然后将水稻根系、叶鞘、叶片分开,称取鲜重后烘干。植物中Cd与Ca含量:用HNO3-H2O2消解,用原子吸收分光光度计(ICE-3500)测定,每个样品3个重复,同时用芹菜标准物质(GBW 10048)进行质量控制,试验过程中均使用优级纯药品与超纯水。
2 结果与分析
2.1 Ca2+对水稻根系、叶鞘及叶片Cd与Ca吸收的影响
由图1可知,在Cd 1与Cd 2胁迫下,水稻根系对Cd的吸收能力地方品种<杂交品种,添加Ca2+对红优2号根系Cd吸收呈先增加后降低的趋势,其他水稻品种根系对Cd的吸收均呈降低趋势。在Cd 1与Cd 2浓度胁迫下,添加Ca 2以上浓度处理水稻根系对Cd的吸收差异均达显著水平(p<0.05),在Cd 1 Ca 3处理时各品种水稻根系Cd含量依次为:红优2号>C两优华占>宜香优2115>黑糯72,与Cd 1 Ca 0相比分别下降33.63%、52.24%、68.79%、72.60%;在Cd 2 Ca 3处理时各品种水稻根系Cd含量依次为:C两优华占>红优2号>黑糯72>宜香优2115,与Cd 2 Ca 0相比分别下降51.71%、58.39%、71.38%、80.21%。综上所述,Cd胁迫下添加Ca2+能够降低根系对Cd的吸收。
由图2可知,在Cd 1与Cd 2胁迫下,水稻叶鞘中Cd的含量地方品种低于杂交品种,红优2号叶鞘中Cd的含量随着Ca2+浓度的增加呈先增加后降低的趋势,在Cd 1与Cd 2浓度胁迫下,添加Ca 2、Ca 3水平水稻叶鞘Cd含量均显著降低,且在Ca 3水平下,水稻叶鞘对Cd的吸收量最小,在Cd 1 Ca 3处理水稻叶鞘Cd含量依次为:红优2号>C两优华占>宜香优2115>黑糯72,与Cd 1 Ca 0相比分别降低47.85%、58.50%、67.86%、65.60%,在Cd 2 Ca 3处理各水稻叶鞘Cd含量依次为:C两优华占=红优2号>黑糯72>宜香优2115,与Cd 2 Ca 0相比分别降低67.77%、39.03%、60.54%、73.59%。综上所述,Cd胁迫下添加Ca2+能够降低水稻叶鞘中Cd的含量。

注:不同小写字母表示在同Cd胁迫下不同施Ca量处理对水稻根系Cd吸收间差异性显著(p<0.05)。下同。图1 Ca2+对Cd胁迫下水稻根系Cd吸收的影响Fig.1 Effect of Ca2+ on cadmium uptake by rice roots under cadmium stress

图2 Ca2+对Cd胁迫下水稻叶鞘Cd吸收的影响Fig.2 Effect of Ca2+ on cadmium uptake by rice sheath under cadmium stress
由图3可知,在Cd 1与Cd 2胁迫下,水稻叶片中Cd含量本地品种低于杂交品种。在添加Ca2+后,各处理水稻叶片中Cd的含量与Cd 1 Ca 0、Cd 2 Ca 0相比均显著下降,且随着Ca2+浓度的增加而降低,在Cd 1 Ca 3与Cd 2 Ca 3时,降低幅度最大。在Cd 1 Ca 3处理下水稻叶片Cd含量依次为:宜香优2115>C两优华占>红优2号>黑糯72,与Cd 1 Ca 0相比分别下降60.03%、75.04%、58.19%、64.66%;在Cd 2 Ca 3处理下水稻叶片Cd含量依次为:宜香优2115>黑糯72>红优2号>C两优华占,与Cd 2 Ca 0相比分别下降59.35%、56.77%、46.39%、71.73%。综上所述,在Cd胁迫下Ca2+能够降低水稻叶片中Cd含量。
由表1可知,4个水稻品种各器官Ca含量分布规律为:茎鞘>叶片>根系,4个品种Ca含量分布规律为:C两优华占>红优2号>宜香优2115>黑糯72。在Cd胁迫下红优2号、C两优华占及宜香优2115均降低了水稻根系中的Ca含量,在Cd 1与Cd 2胁迫下添加Ca2+,水稻各器官中的Cd含量均显著提高,4个水稻品种在两种Cd浓度胁迫下各器官Ca含量Ca 3均显著高于Ca 1。综上所述,在Cd胁迫下添加Ca2+,能够促进水稻根系对Ca的吸收,吸收量随着Ca的增加而增加。
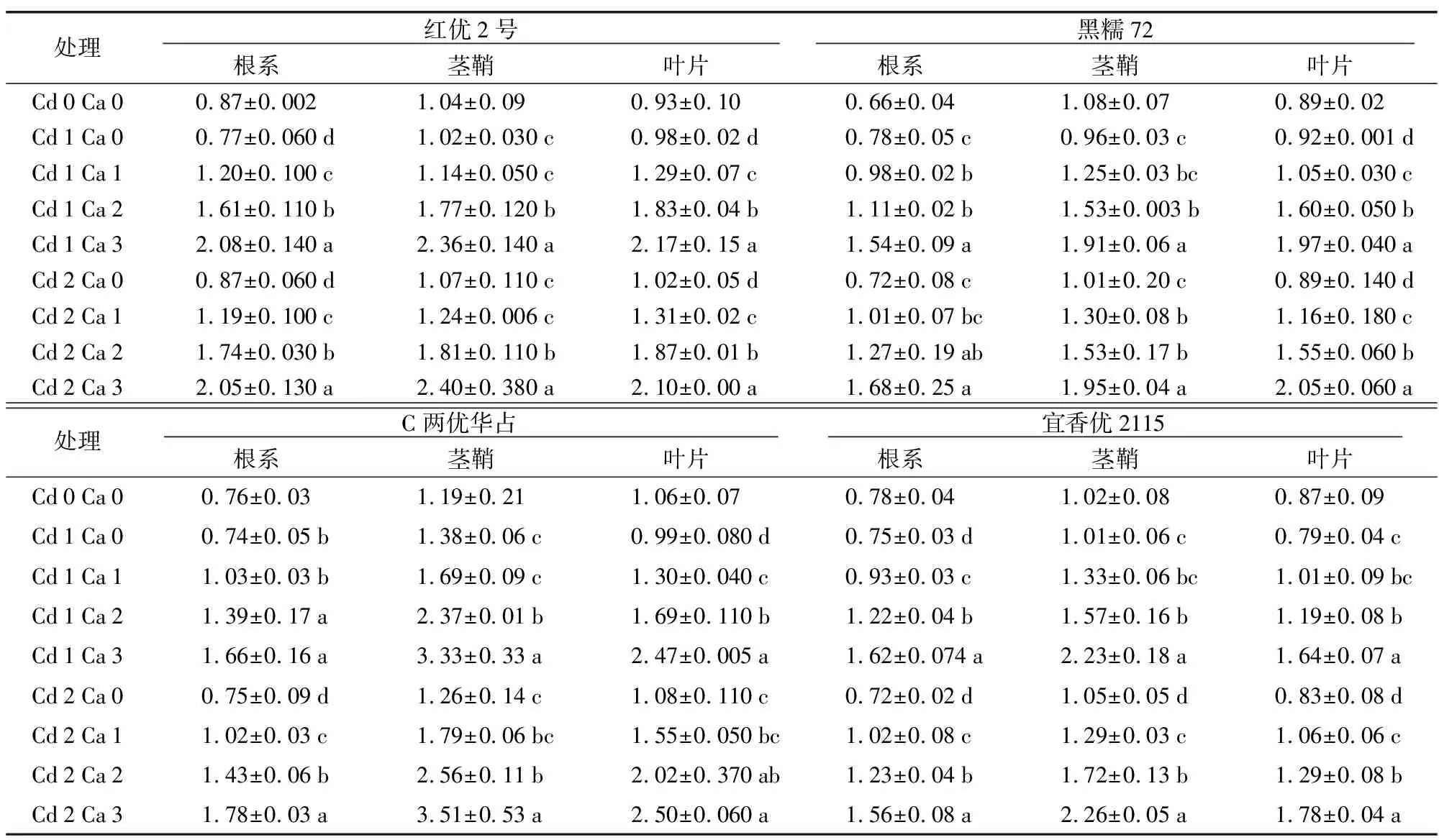
表1 4个水稻品种各器官Ca含量 Table 1 Calcium content in each organ of 4 rice varieties 单位:g/kg

图3 Ca2+对Cd胁迫下水稻叶片Cd吸收的影响Fig.3 Effect of Ca2+ on cadmium uptake in rice leaves under cadmium stress
2.2 Ca2+对Cd在水稻各器官迁移系数的影响
由图4可知,各水稻品种在不同浓度Cd胁迫下Ca2+对叶鞘/根系转移系数的影响有差异。贵州地方品种红优2号与黑糯72在不同浓度Cd胁迫下Ca2+对叶鞘/根系转移系数的影响没有差异,杂交品种C两优华占与宜香优2115在高浓度Cd胁迫下添加Ca2+有显著差异,C两优华占在Cd胁迫下添加Ca2+转移系数均降低,且有显著差异,添加Ca2+处理与Cd 2 Ca 0相比,转移系数降低33.33%~35.80%,由此说明在Cd胁迫下添加Ca2+能够降低C两优华占中Cd由根系向叶鞘与叶片运输。宜香优2115在高浓度Cd胁迫下Ca2+对叶鞘/根系转移系数均升高,且有显著差异,添加Ca2+处理与Cd 2 Ca 0相比,转移系数升高18.52%~48.15%,由此说明在Cd胁迫下添加Ca2+能加快宜香优2115中Cd由根系向叶鞘与叶片运输。
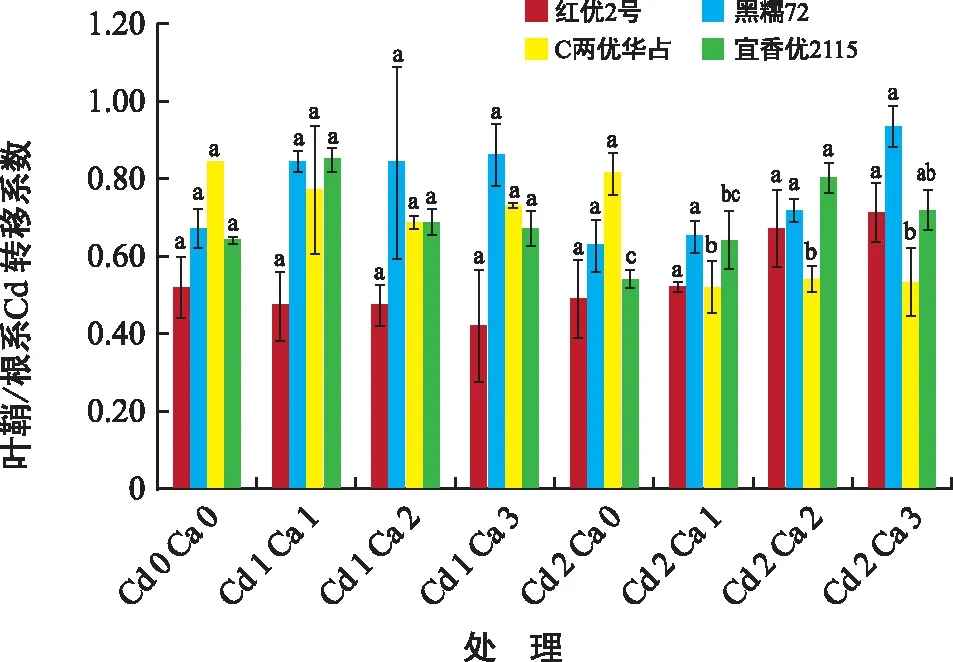
图4 Ca2+对Cd胁迫下水稻叶鞘/根系Cd转移系数的影响Fig.4 Effect of Ca2+ on leaf sheath/root transfer coefficient of rice under cadmium stress
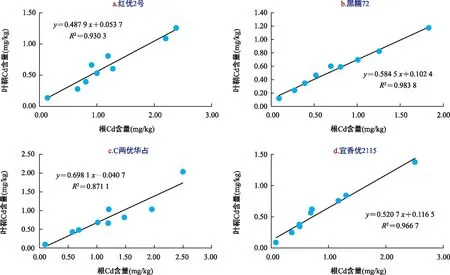
图6 不同水稻品种根部与叶鞘中Cd含量相关性分析Fig.6 Correlation analysis of cadmium content in roots and sheaths of different rice cultivars
由图5可知,各水稻品种在不同浓度Cd胁迫下Ca2+对Cd在叶片/叶鞘的转移系数有差异。红优2号在Cd 1 Ca 1处理下与同浓度Cd胁迫下有显著差异,Cd 1 Ca 1与Cd 1 Ca 0相比Cd在叶片/叶鞘间的转移系数降低39.78%;黑糯72各处理间没有显著差异;C两优华占在Cd 1水平胁迫下Ca2+对Cd在叶片/叶鞘的转移系数降低12.97%~39.77 %,在Cd 2水平胁迫下添加Ca2+处理,Cd 2 Ca 1与Cd 2 Ca 0相比转移系数升高39.16%,存在显著差异;宜香优2115在同Cd胁迫水平下各处理间无显著差异。
2.3 Ca2+对不同水稻品种各器官中Cd含量相关性的影响
由图6可知,在不同浓度Ca2+处理下,水稻根系与叶鞘中Cd积累量呈显著正相关,不同水稻品种间相关性强弱关系为:黑糯72>宜香优2115>红优2号>C两优华占,说明添加Ca2+对黑糯72根系中的Cd向叶鞘中迁移的促进作用最强,对C两优华占根系中Cd向叶鞘迁移促进作用最弱。由此得出,添加Ca2+可加速黑糯72中Cd由地下部分向地上部分转移,增加地上部分的风险。
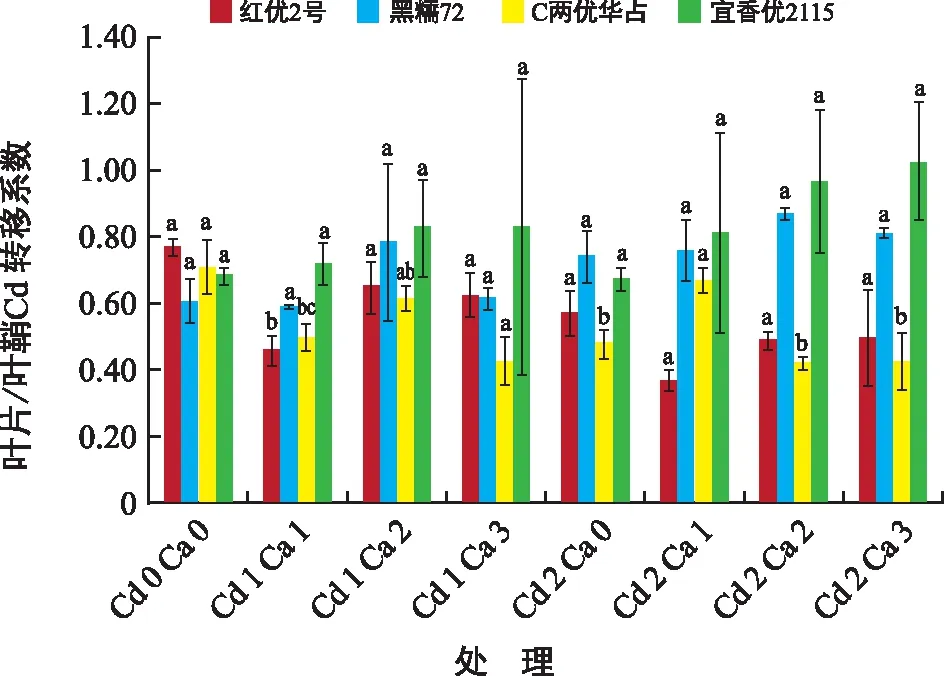
图5 Ca2+对Cd胁迫下水稻叶片/叶鞘Cd转移系数的影响Fig.5 Effect of Ca2+ on leaf/leaf sheath transfer coefficient of rice under cadmium stress
由图7可知,在不同Ca2+浓度处理下,水稻叶鞘与叶片中的Cd含量呈显著正相关,不同水稻品种间相关性强弱关系为宜香优2115>黑糯72>C两优华占>红优2号,说明Ca2+对宜香优2115叶鞘中Cd向叶片中迁移的促进作用最强,对C两优华占的促进作用较弱,由此得出,Ca2+能够增加宜香优2115叶鞘中Cd的移动性,增加水稻叶片中Cd的含量。由此可知,Ca2+能够促进宜香优2115与黑糯72中Cd的移动性,能够增加水稻叶鞘与叶片中Cd风险,Ca2+能够降低C两优华占与红优2号中Cd的移动性,让大部分Cd储存在根系中,降低了叶鞘与叶片中Cd的风险。
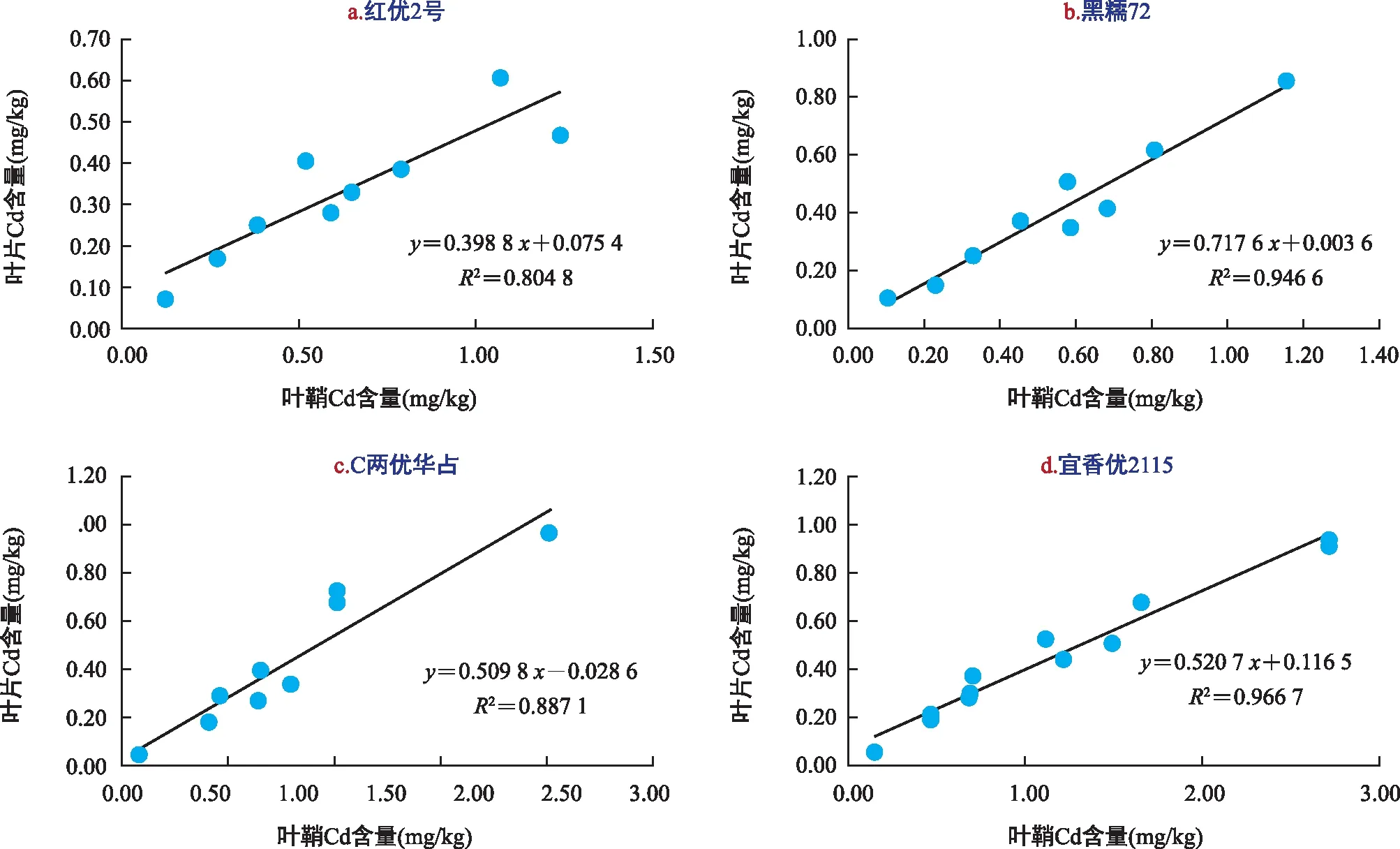
图7 水稻品种叶鞘与叶片中Cd含量相关性分析Fig.7 Correlation analysis for cadmium content between leaf sheath and leaves of rice varieties
3 讨 论
3.1 Ca2+对水稻Cd积累的机理研究
Ca是植物生长发育所必需的营养元素,参与细胞壁的结构合成,并且还能与磷脂分子联结起来,稳定细胞膜,对膜结合蛋白的稳定性、参与细胞内稳态也有重要作用[7]。水稻对Cd的吸收主要通过外质体与共质体途径进入根系维管柱中,外质体途径是水稻植株内外Cd浓度差使Cd2+扩散进入细胞,然后通过质外空间进入根系维管柱中,属于被动运输,随着Cd浓度差异的增加,被动吸收越多;共质体途径是利用代谢能量使Cd载体蛋白进入根系细胞,通过胞间连丝转移至根系维管柱中,属于主动运输[2]。水稻对Ca的吸收主要以共质体途径,是通过能量代谢使Ca载体蛋白进入植物根系,最后通过胞间连丝进入根系维管柱中,属于主动运输[17],因此在Cd胁迫下Ca2+导致水稻对Cd的吸收能力降低有3个可能,第1是Cd与Ca相互竞争能量,第2是Cd与Ca竞争载体蛋白,第3是Cd与Ca载体蛋白竞争通道,此外,Cd2+与Ca2+因半径相近电性相同,共同竞争水稻根系离子吸附位点,也会降低水稻对Cd的吸收[8]。本研究在营养液中添加Cd2+与Ca2+,不同浓度Ca2+能够降低水稻苗期对Cd的吸收,且随着Ca2+浓度的增加,各部位Cd含量差异越明显,水稻各器官Cd分布遵循根系>叶鞘>叶片的规律。由表3~表6可知,4个水稻品种各器官中Cd和Ca含量均呈极显著相关性,相关性强弱为:杂交品种>本地品种,原因是杂交品种代谢能力强,Cd与Ca在根系、叶鞘及叶片间的移动速率高[2-3]。水稻根系中Cd与Ca均呈负相关,相关性强弱顺序为:宜香优2115>黑糯72>红优2号>C两优华占,C两优华占是杂交品种,水稻生长比红优2号旺盛,根系对Cd的吸收能力较强,根系中Cd与Ca相关性较差,可能是由于在低浓度Ca2+处理下,根系对Ca的吸收能力较低,对Cd的吸收作用增强,导致根系中Cd大量富集[8],在Cd 1 Ca 1与Cd 2 Ca 1处理下,黑糯72、C两优华占、宜香优2115根系含量均高于红优2号,可能是由于在低Ca浓度下,水稻根系向叶鞘转移速率Ca大于Cd,Cd大量在根系聚集,导致根系Cd含量增加[9]。4个水稻品种苗期杂交品种对Cd的吸收能力较强,添加Ca2+均显著提高根系中Ca含量,两个地方品种水稻品种有差异,红优2号是籼稻、黑糯72是粳稻,有研究表明,籼稻比粳稻对Cd的吸收能力强[7],且红优2号根系Ca含量比黑糯72低。水稻杂交品种根系比本地品种长势强,本地品种水稻根系、叶鞘及叶片中Cd含量占比范围分别为44.70%~55.26%、29.47%~32.08%、15.27%~23.23%,杂交品种根系、叶鞘及叶片中Cd含量占比范围分别为46.28%~49.35%、30.24%~32.77%、17.88%~23.48%,可知Ca2+对根系中Cd的转移速率杂交品种比本地品种强,杂交品种根系中的Cd易向地上部位转移,本地品种中的Cd更多的是储存在根系中,可能是因为水稻细胞形态维持受有丝分裂的过程受Ca调蛋白(Ca2+-CaM)的调控,杂交品种有丝分裂强于本地品种,CaM蛋白调控能力本地品种比杂交品种强,从而阻碍根系对Cd的吸收,在低浓度Cd胁迫下,Cd2+通过Ca2+通道进入根系,Cd含量在体内升高会抑制Ca的吸收,在高浓度Cd胁迫下,Cd2+占用Ca2+通道需要消耗大量的能量,CaM蛋白的活性受到影响,导致水稻根系不良生长,从而减少水稻对Cd的吸收。Wei-Jie Xue等[18]研究得出,Ca/Mn条件下水稻根系影响Cd的化学形态,使得Cd的有效性降低,从而减少了Cd对水稻的毒害作用,与本研究结果一致,可能是因为Ca与Cd的半径相近电价相同,Ca2+也会通过影响根系的生长,从而减少Cd的毒害作用。Johnson P T等[19]研究得出,Cd胁迫下Ca2+能够降低过氧化氢与丙二醛的含量,减少自由基的合成,增强酶抗氧化性的活性减缓水稻对Cd的吸收,可能是Ca参与水稻抗氧化酶的调节,降低了水稻体内自由基的合成,减少了Cd的毒害作用。徐胜光等[20]认为,Ca能够刺激水稻根系产生有利于提升pH值的物质增加土壤pH值,使得Cd的有效性含量降低,减少水稻根系对Cd的吸收。本研究中可能是Ca2+促进水稻根系分泌中多种有机酸能起到螯合Cd的作用,降低营养液中Cd的活性[21]。周金林等[22]研究表明,Cd胁迫能够抑制叶绿素a与叶绿素b的合成,使气孔增加,光合作用减弱,同时,还导致丙二醛含量升高,过氧化氢酶、过氧化物酶、超氧化物歧化酶的活性增加,Cd积累能力不同的品种间活性氧差异显著。朱智伟等[23]研究表明,Ca能够减少活性氧与丙二醛对植物的毒害,调控植物叶绿素含量与气孔导度,增强光合作用。因此,在Cd胁迫下施Ca能够降低Cd的毒害作用,导致水稻对Cd的吸收降低。何俊瑜等[24]研究发现,Cd胁迫能够使根尖细胞有丝阻滞,并使染色体畸变率增加;张思思[25]研究表明,Ca能够调控根毛极性生长,且Ca是构成细胞壁与细胞膜等结构的必须元素,在维护正常结构与功能、减缓与减少细胞膜的毒害与损伤等方面发挥重要作用,本研究结果与之一致,可能是因为Ca通过调控水稻 根系增强有丝分裂,促进水稻细胞壁的合成与强化,导致水稻根系对Cd的吸收能力减弱。

表2 红优2号各器官Cd与Ca含量相关性分析Table 2 Correlation analysis of Cd and Ca content in various organs of Hongyou 2

表3 黑糯72各器官Cd与Ca含量相关性分析Table 3 Correlation analysis of Cd and Ca contents in organs of Heinuo 72
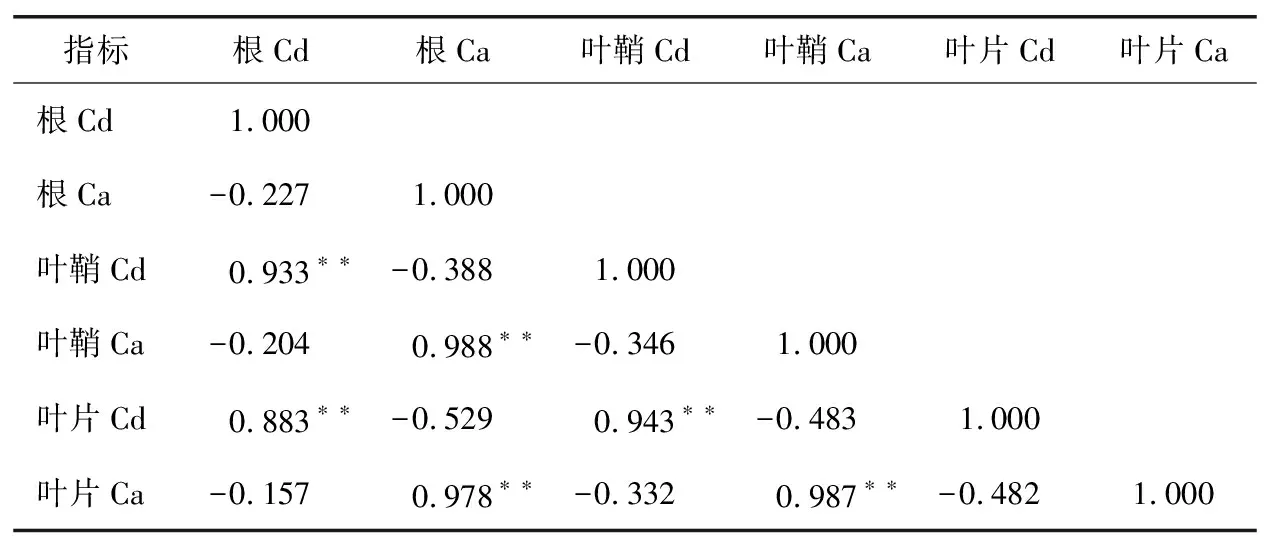
表4 C两优华占各器官Cd与Ca含量相关性分析Table 4 Correlation analysis of Cd and Ca contents in variousorgans of C Liangyouhuazhan
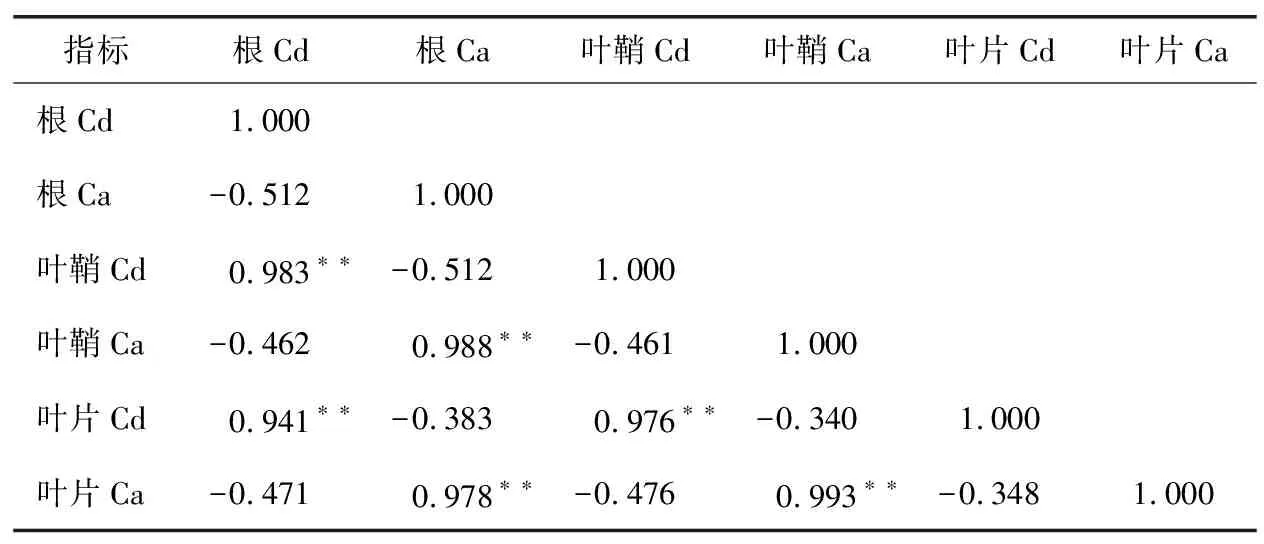
表5 宜香优2115各器官Cd与Ca含量相关性分析Table 5 Correlation analysis of Cd and Ca contents in various organs of Yixiangyou 2115
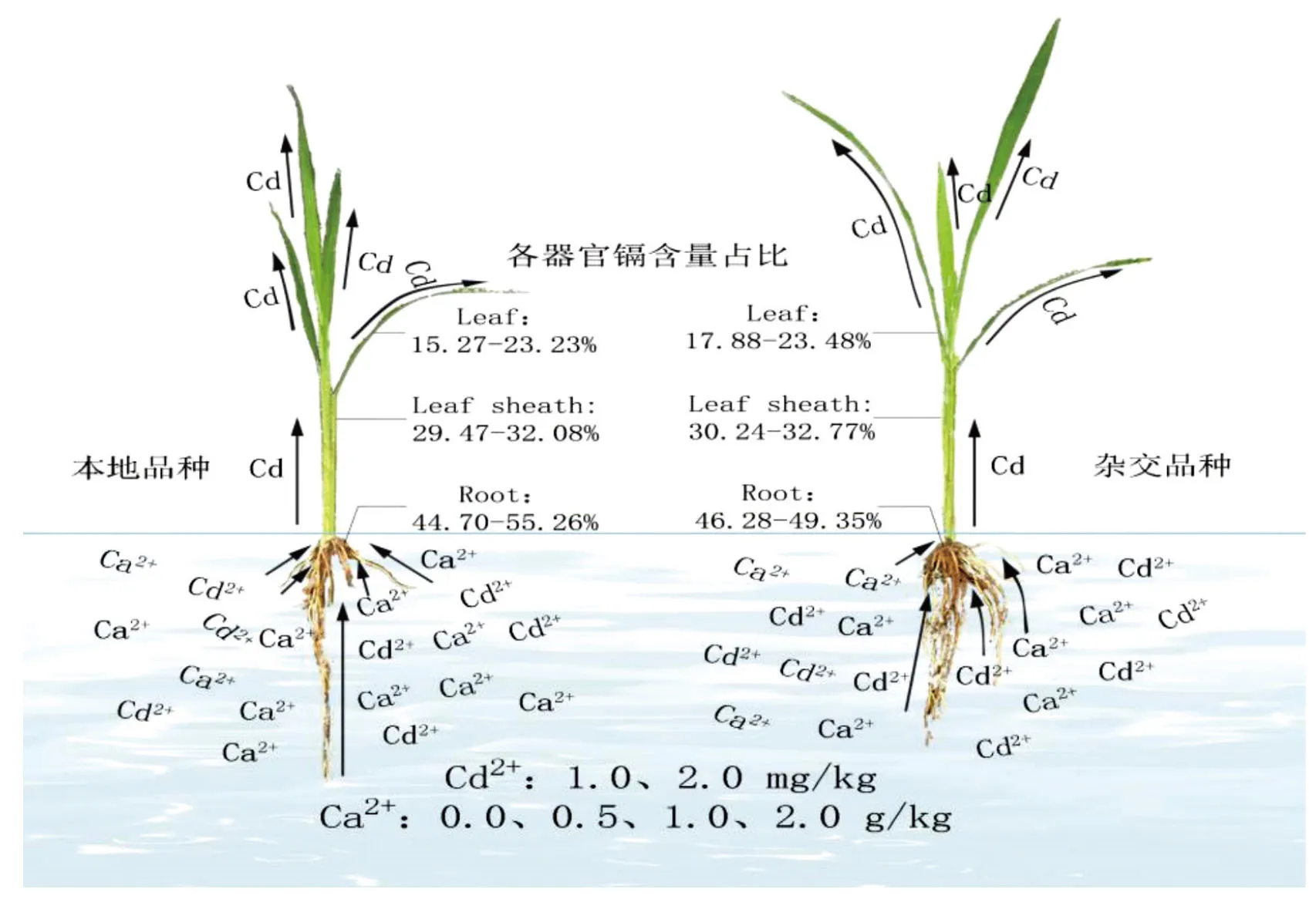
图8 Ca2+离子对水稻Cd吸收的示意图 Fig.8 Schematic diagram of Ca2+ uptake by cadmium in rice
3.2 Ca2+对不同水稻品种Cd吸收差异性研究
由图8可知,杂交水稻对Cd的吸收转运能力比地方品种强,已有研究表明,不同水稻品种对重金属Cd的吸收能力不同,各器官中含量也有显著差异[3-4];杨祥田等[27]研究表明,水稻根系、茎、叶、穗、糙米中Cd含量与水稻类型有显著相关,其中糙米中Cd含量呈籼型杂交稻>粳型杂交稻>常规籼稻>常规粳稻,说明水稻对Cd的吸收能力为杂交稻>常规稻,籼稻>粳稻,本研究结果与之基本一致,可能是因为杂交稻生长力较强,能够更加充分地利用养分,从而促进水稻对Cd的吸收。本研究各水稻品种中Cd含量依次为根系>叶鞘>叶片,这与鸿凯等[28]的研究结果一致。不同水稻品种对Cd吸收和运转机制不同,因此造成品种之间各器官Cd含量有巨大差异[29]。本研究表明,贵州本地品种比杂交品种各器官Cd含量低,本地水稻根系Cd含量表现为:红优2号>黑糯72,叶鞘与叶片Cd含量均为:黑糯72>红优2号,说明在黑糯72根系中Cd易向地上部转移,杂交品种根系、叶鞘及叶片Cd含量均显示出C两优华占>宜香优2115,这说明各水稻品种之间Cd的吸收分配存在差异[30]。
由图9可知,杂交品种(C两优华占、宜香优2115)比本地品种(红优2号、黑糯72)水稻根系侧根发达,须根系较多,水稻株高长势杂交品种比本地品种强。LIU J G等[31]研究表明,水稻对Cd的吸收主要受根系生长的影响,根系较发达的品种对Cd的吸收能力较强,本研究结果与之一致。Hui Yu等[32]研究表明,杂交品种比本地品种各器官Cd的含量高,可能是因为本地品种已经适应当地的温度、湿度及光照情况;因长期生长在Cd地质高背景地区[12],根系对Cd的感应能力较差,而杂交品种选育环境与推广环境有差异,特别是土壤中Cd2+与Ca2+含量差异较大,因此杂交品种对营养液中Cd2+与Ca2+感应能力较强,杂交品种对Cd的吸收速率大于本地品种。Li Kun等[33]研究表明,在轻微Cd污染区,低富集Cd水稻品种稻米中Cd含量超标,可能是筛选研究环境因子不同引起的,即各水稻品种对Cd积累受地力条件、农艺性状及温度湿度等环境因子的影响,故水稻对Cd的吸收可能还受温度、湿度、光照及土壤中Ca2+浓度等因素的影响。

图9 不同水稻品种苗期长势情况Fig.9 Growth situation of different rice varieties in seedling stage
4 结 论
通过水培试验,探究Cd胁迫下添加Ca2+对水稻苗期Cd积累的影响,初步得到以下结论:
1) 在Cd胁迫下Ca能够显著降低水稻根系、叶鞘及叶片中Cd含量,且在2 g/kg处理下降Cd效果最为显著;
2) 在Cd胁迫下各水稻品种对Cd的吸收能力有明显差异,强弱关系为:宜香优2115>C两优华占>红优2号>黑糯72,杂交品种比本地品种Cd吸收能力强;
3) 水稻根系、叶鞘及叶片中Cd的含量关系为:根系>叶鞘>叶片,且根系对叶鞘及叶片中Cd与Ca含量均有显著影响。
