关于植物生殖概念的思考
——植物学教材质疑(七)
2022-05-15黎维平
黎维平
(湖南师范大学生命科学学院,中国湖南 长沙 410081)
绿色植物(Viridiplantae)由绿藻(Phycophyta)和有胚植物(embryophytes)组成,其中绿藻包括绿藻门(Chlorophyta)和链形绿藻(streptophyte green algae)[1]。分子系统学研究显示,链形绿藻和有胚植物是单系,属于链型植物门(Streptophyta)[1]。为了避免陷入复杂的生殖概念泥淖,本文把所讨论的植物限定为链型植物(streptophytes)。链型植物中的链形绿藻,简称链形藻(streptophyte algae),相当于传统的轮藻纲(Charophyceae)[1~3],其中的轮藻目(Charales)是有胚植物的姊妹类群[1,3]。有胚植物又称高等植物,包括苔藓植物(bryophytes)、广义的蕨类植物(pteridophytes;含石松类lycopodiophytes)、裸子植物(gymnosperms)和被子植物(angiosperms)。
生殖(reproduction)或繁殖(propagation)是生物的基本属性之一,是产生新的个体来延续后代的过程[2,4~10]。生殖一般被定义为“以生殖细胞发育下一代新个体的方式”[8~10]。繁殖与生殖是近义词,有时二者可以通用[8,10]。但是,二者有一些差别:1)生殖包括有性生殖(sexual reproduction)和无性生殖(asexual reproduction),其过程中一定出现生殖细胞,营养繁殖(vegetative propagation)等则不包括在内,所以生殖的外延要窄于繁殖[8~10];2)繁殖含有增殖(multiplication)的意义[2,6,8];3)繁殖一词的英文propagation还含有传播的意义。
上述植物生殖概念,看似较为简单明晰,但深究起来,植物学教材的生殖概念含混不清,值得仔细推敲。
1 有性生殖
有性生殖是植物学教材概念混乱的“灾区”之一。
1.1 由合子发育成新个体,属于有性生殖的范畴吗?
有性生殖常被定义为“经过两性细胞结合,产生合子,由合子发育成新个体的生殖方式[2,6,10]”。生殖是新个体产生的过程,精、卵融合形成合子,单细胞植物新个体或多细胞植物新个体的第一个细胞诞生,生殖过程便结束;接下来则是植物体的生长发育。由合子发育成单细胞生物或多细胞生物,是个体发育而非生殖过程,所以,合子的发育不应包括在生殖概念之中。如果将生殖和新一代个体的早期发育剥离开来,下文提到的种子形成不是生殖过程的说法就比较容易接受了。
1.2 有性生殖等于受精作用加减数分裂吗?
动物无世代交替,通常减数分裂和受精作用紧密联系,形成一个连续的过程,构成较为一致的有性生殖模式。但是,链型植物的有性生殖随着进化,发生着巨大而深刻的转变,可以归为6类(表1)。
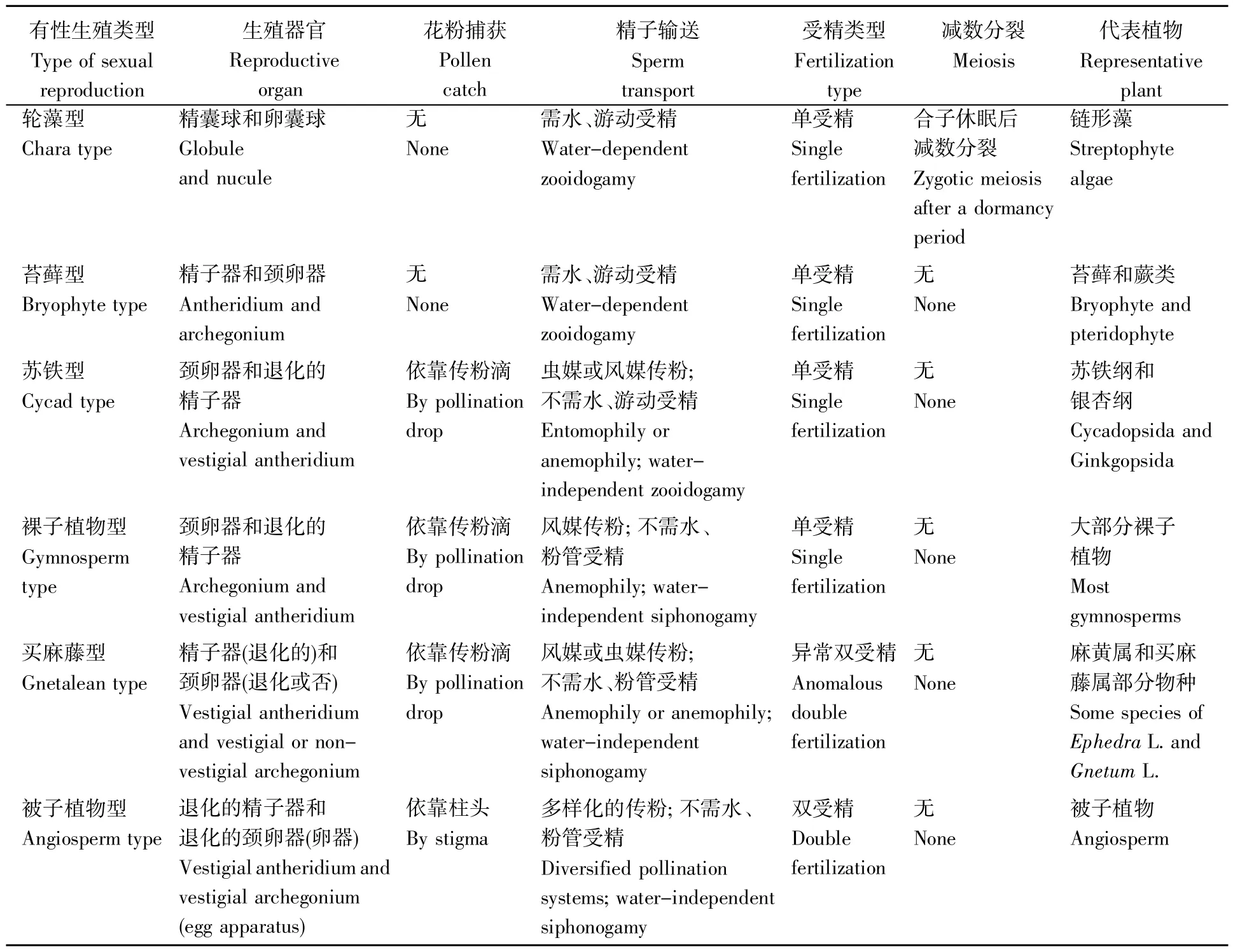
表1 链形植物有性生殖的分类Table 1 Classification of sexual reproduction in streptophytes
1.2.1 轮藻型有性生殖:需水单受精-合子休眠-合子减数分裂
有性生殖是否等于受精?卵配才受精;生殖细胞的融合(包括同配、异配和卵配)是有性生殖的核心,但通常不是其全部。轮藻门植物的有性生殖过程并非两性细胞结合之简单过程,而是需经历4个较漫长的阶段:1)雌、雄配子在卵囊和精子囊中分别通过有丝分裂形成;2)精子被释放到水中后游至颈卵器,与其内的卵细胞结合成受精卵;3)受精卵(合子)休眠;4)合子减数分裂,形成孢子,至此有性生殖完成。不能把轮藻有性生殖的终点定为受精作用完成,因为受精产生的合子并不是任何植物体的细胞,只是一个过渡性结构。合子的功能就是休眠,并通过减数分裂产生新植物体的第一个细胞——孢子(1合子减数分裂产生4个核,其中3个退化,1个发育[6]),至此新植物体诞生,有性生殖完成。轮藻的孢子发育成原丝体,进而形成植物体,这些是生殖之后的生长发育过程。轮藻型有性生殖的特征包括需水受精、合子休眠、合子减数分裂(表1)。
1.2.2 苔藓型有性生殖:需水单受精
链形藻进化为高等植物最大的变化之一是通过合子的延迟减数分裂导致孢子体的出现,演化出世代交替[1]。同时,随着孢子体世代的出现,链形藻由复杂的有性生殖转变成高等植物世代交替中配子体的有性生殖(受精作用)和孢子体的无性生殖(减数分裂)。苔藓和蕨类植物的配子体通过有丝分裂产生生殖细胞(雌、雄配子),然后精子借助水游至颈卵器,通过受精作用形成合子——孢子体的第一个细胞(可称为单细胞胚single-cell embryo),至此有性生殖完成,这是苔藓和蕨类植物共有的有性生殖方式,为简洁起见,称之为苔藓型有性生殖(表1),其中不包括减数分裂。而二者孢子体通过孢子母细胞的减数分裂产生生殖细胞——大、小孢子,孢子是配子体的第一个细胞,配子体的第一个细胞诞生,标志着无性生殖(孢子生殖sporogony)完成。
有的教材[4,11~13],将减数分裂与有性生殖捆绑在一起,将苔藓植物的受精作用到孢子形成及其传播当成有性生殖,而把藓类通过配子体碎片和苔类通过胞芽进行的营养繁殖称为无性生殖[12~14]。将减数分裂与有性生殖捆绑在一起的原因,一是将遗传变异与有性生殖相联系,减数分裂(孢子生殖)过程中有遗传重组,而亲子遗传上的一致性被说成无性生殖的特征[4,14];另一方面可能是,在苔藓植物中,孢子体寄生在配子体上,且结构相对简单、存活时间较短,其无性生殖——孢子生殖常被忽视而被看成有性生殖的一个阶段。而在蕨类植物中,孢子体和配子体各自独立生活,所以一般不会把原叶体(配子体)的有性生殖和孢子体的无性生殖(孢子生殖)合在一起,但是原叶体往往由于不易观察到而受到忽略。到了种子植物,由于配子体寄生于孢子体内,二者各自的生殖过程又被错误地捆绑起来[13]。配子体的寄生改变了其营养生长和生殖过程,但植物体还在,生殖也未消失。将苔藓和种子植物的有性生殖定义为跨世代现象(transgenerational reproduction,一个生殖过程跨越不同世代),这是不恰当的。
苔藓植物进化成蕨类植物,在孢子体和配子体的发达程度上有了巨大变化,表现在蕨类植物的孢子体发达,独立生活,具维管组织和真正的根、茎、叶;配子体为不显著的、独立生活的原叶体。但是,苔藓植物进化到蕨类植物在生殖方式上并无显著区别,其有性生殖仍是需水受精型有性生殖。
学术界在有性生殖概念上的混乱更甚于植物学教材,除了上述提到的将苔藓植物的无性生殖隐藏在有性生殖概念中[14]之外,一些论文中还有更奇特的观点,比如:“有性繁殖即植物(原叶体)产生具有繁殖和休眠作用的孢子,孢子在适宜条件下能直接发育成新个体[15]”。且不说原叶体作为蕨类植物的配子体,无法产生孢子,将受精过程排除在有性生殖之外,取而代之的是无性生殖产生孢子,该定义很是奇特,令人困惑。
1.2.3 苏铁型:传粉-不需水游动受精-单受精
在裸子植物系统发育树上,苏铁纲和银杏纲组成是一个分支,为基部类群[16]。二者的共有祖征是精子具有纤毛(不少教材称“鞭毛”),具运动精子,无真正的花粉管,其所谓的花粉管为伸向珠心获取营养的吸器(寄生根)[17]。教材一般未指出纤毛的功能[8~9],或认定其为无功能的痕迹器官(vestige)[2]。纤毛是有功能的结构,帮助精子游至颈卵器,实现受精。虽然苏铁纲和银杏纲植物没有运输精子的花粉管,但它们通过虫媒或风媒传粉、传粉滴对花粉的捕捉和转运以及精子的游动,使受精作用摆脱了水的束缚[17]。苏铁纲和银杏纲的有性生殖方式称为“苏铁型”,特征是其受精既不受外界的水束缚,也不依赖于花粉管运送精子,而靠传粉和精子游动,最后完成单受精(表1)。
1.2.4 裸子植物型:传粉-粉管受精-单受精
现存裸子植物中,除上文提到的苏铁纲和银杏纲外,大部分植物通常依靠风媒传粉、传粉滴捕捉花粉和将花粉吸入花粉室、花粉管运输精子和受精等系列过程完成有性生殖。因大部分裸子植物采用此方式,所以这种有性生殖被称作裸子植物型,其与苏铁型唯一的不同是粉管受精(表1)。
1.2.5 买麻藤型:传粉-粉管受精-异常双受精
裸子植物和被子植物均属于种子植物,二者的主要区别之一是前者单受精,后者双受精。但是,有的买麻藤纲(Gnetopsida)植物[18]有双受精现象,如麻黄属植物粗麻黄(Ephedra nevadensis)、长叶麻黄(E.trifurca)和买麻藤属植物Gnetum gnemon。准确地说,买麻藤纲植物有异常双受精(anomalous double fertilization),不是真正意义上的双受精,因为:1)与被子植物的双受精不同,买麻藤纲植物的第二受精不是精子和染色体数为2N的中央细胞结合。在麻黄属中,第二精子与1N的腹沟核融合[18];而在买麻藤属中,1个花粉管释放的两个精子与未分化的雌配子体中的两个游离核分别受精[18];2)其第二受精产物发育成胚,而不是胚乳[18]。目前,尚未发现整个买麻藤纲植物都有异常双受精,本文将这种有性生殖命名为“买麻藤型”(表1),是因为其仅见于买麻藤纲。买麻藤型有性生殖与裸子植物型的区别在于前者有单受精,后者演化出异常双受精(表1)。
1.2.6 被子植物型:传粉-粉管、双受精
与裸子植物的有性生殖过程相比,被子植物从传粉到受精,各阶段都有显著差异(表1)。被子植物的传粉多样化,并有与此相适应的各种传粉综合征;捕捉花粉的是柱头而不是传粉滴;花粉管将精子从柱头,经花柱,输送至子房中的胚珠内,最后抵达卵器(包括1卵细胞和2助细胞,由颈卵器退化而成)并释放2精子;行双受精,即小精子和1N的卵细胞受精,大精子和2N的中央细胞受精。被子植物有性生殖过程不包括减数分裂,“通过减数分裂发育出进行有性生殖的卵细胞”[19]等说法是不严谨的。
总之,链型植物的有性生殖都含有核心过程——受精作用(表1),但是轮藻没有世代交替,其有性生殖完成、产生新个体有赖于减数分裂的参与;而高等植物有世代交替,有性生殖和减数分裂发生在不同世代,由不同世代的植物体完成。高等植物有性世代的配子体行有性生殖,即通过受精作用导致新植物体的发生,无需减数分裂;减数分裂是无性世代的孢子体实行的生殖过程——无性生殖。此外,从受精需水到因传粉而摆脱水的束缚,从传粉滴捕获花粉到柱头捕获花粉,从游动受精到粉管受精,从单受精到异常双受精和双受精,高等植物的有性生殖经历了一系列的演化。
2 无性生殖
无性生殖有广义和狭义两种定义。广义上的无性生殖是把有性生殖之外的所有生殖方式均归为无性生殖[13,19];狭义的无性生殖是中文植物学教材上普遍定义的孢子生殖[2,5~10,19~20]。采用哪种定义都有道理,问题是在无性生殖定义上存在一些混乱,如将营养繁殖等同于无性生殖。
姜汉侨等[21]指出,“许多植物都具有无性生殖的能力,即营养增殖或称无性系生长,通过营养生殖体如珠芽、匍匐茎、根茎、枝条、分蘖株等形成新的植株,并与原来的植株保持一致的基因型”。另一些教材[4,12~13]也强调无性生殖中后代和亲代DNA上的一致性。显然,按照亲子遗传上相同的标准,在高等植物中无性生殖就等同于营养繁殖,还应再加上未减数胚囊和不定胚等两种形式的无融合生殖。但是,高等植物的孢子生殖是减数分裂过程,2N的孢子体产生1N的配子体的第一个细胞(大、小孢子),亲子二者在遗传上显然有较大差异,与上述无性生殖概念不同。可见,上述无性生殖定义[4,12~13,21]大有问题。
上文提到,苔藓植物的孢子生殖常被纳入有性生殖的一个阶段,例如:地钱属(Marchantia L.)胞芽进行的营养繁殖被称为无性生殖,而其孢子生殖未被当作无性生殖[11,14~15]。这是无性生殖概念另一方面的问题。
种子植物的“孢子不能脱离母体而独立生活,只能进行异养的寄生生活,所以,被子植物中不能划分出自然的无性生殖”[2],这种说法缺乏逻辑性。因为种子植物通过生殖产生的生殖细胞(孢子)可发育成新的生物体(配子体),这些新个体无论以何种方式(寄生还是自养)生存,都不能否认此生殖是其自然生命过程,不能用来确定生殖存在与否。
无融合生殖中的二倍体无融合生殖(体细胞无孢子生殖和二倍体孢子生殖,即通过未减数胚囊行无融合生殖)和不定胚,其子代与亲代遗传上一致,符合营养繁殖的特征,也归入营养繁殖。至于单倍配子体无融合生殖(单倍体孤雌生殖和单倍体无配子生殖,即通过减数胚囊行无融合生殖),其与孢子生殖有些类似,即都经历了减数分裂,可归入无性生殖。本文这一提议有些突兀,但值得考虑。
虽然各中文植物学教材[5~10,20]在对繁殖类型划分时将孢子生殖归入无性生殖,但是,其种子植物解剖学部分在叙述大、小孢子母细胞通过减数分裂形成大、小孢子的过程中,没有提到这是一个生殖过程,更没提及这是无性生殖过程。这样处理,增加了理解无性生殖的难度。
3 繁殖乎?传播乎?
有植物学教材认为,“苔藓植物无性生殖产生单倍体的孢子,以孢子进行无性生殖,故苔藓植物属于孢子植物类群之一”[9];类似的说法还有,孢子植物是“以孢子进行繁殖的植物统称”[10]、“藻类、苔藓和蕨类植物主要是通过产生大量的孢子来增加植物个体的数量,称孢子植物”[8];“孢子植物是和种子植物相对而言的,它繁殖时不产生种子,以孢子进行繁殖”[20];孢子植物“用孢子繁殖”[2]。把以孢子进行无性生殖当作孢子植物定义的核心,这很不恰当,因为种子植物的孢子体也通过产生孢子进行无性生殖,进入配子体世代。事实上,孢子植物和种子植物一样,都以孢子进行无性生殖。另一方面,也有与之相对应的说法,例如,种子植物是“以种子进行繁殖的植物统称”[10];种子植物“利用种子增殖本物种的个体数量”[6];“通过种子进行繁殖的植物,称为种子植物”[2];“种子的产生对植物的繁衍具有重要的意义”[8];“种子是植物有性生殖过程中由胚珠发育而来的结构”[19];“种子是植物有性生殖过程的最终产物”[2,7];“种子植物通过有性生殖形成种子”[8];“种子是种子植物特有的繁殖器官,也是种子植物的繁殖单位”[2,5];古蕨(Archaeopteris)“用自由孢子繁殖,而不是用种子繁殖[6]”。这些论述都将种子与繁殖、有性生殖捆绑在一起,也是不正确的。
孢子和种子分属于高等植物的不同世代,在系统演化和个体发育中都无直接关系。但二者常被联系到一起,这是因为二者在功能上存在相似性,即均为传播体(dispersal organs、dispersal units、diaspore或propagule)。孢子植物以孢子作为传播体,这是孢子植物名称的来源(并非定义为行孢子生殖的植物),而种子植物以种子作为传播体则是其名称的依据。与孢子植物的关键区别在于,种子植物适应“受精摆脱水束缚”而演化出孢子的原位发育与传粉,并发展出种子以代替孢子的传播功能[17]。
前文提到,propagation(繁殖)一词既有增殖又有传播的意思,生殖或繁殖和扩散(dispersal)是有着必然联系的两个生物学过程。由于陆生植物固着生长的习性,扩散往往在繁殖之后的某个阶段发生。但是,繁殖和传播是两个截然不同的生物学过程,不能模糊二者的区别,更不能合二为一。“通过繁殖,植物大量地增加了新一代的个体,扩大了生活范围”[6],这一说法给繁殖附加了扩散的内容——“扩大了生活范围”,这不恰当。类似地,种子植物“利用种子增殖本物种的个体数量”[6]之说,给种子的形成附加了繁殖的意义,显然不合适。
有人把传粉当成一种扩散。笔者在本系列文章的开篇[17]中指出,将传粉当作是受精摆脱水束缚的策略,是种子植物实现有性生殖的前提条件;虽然传粉可导致种群间形成基因流,但笔者不认为传粉是一般意义上的植物体传播。植物的传播就是通过传播体实现植物体的位移,改变植物的分布。高等植物营养繁殖发生后,繁殖体(vegetative diaspores或vegetative propagules)会通过各种方式传播,包括匍匐茎的克隆生长;苔藓和蕨类通过无性生殖产生大量孢子后,以孢子作为传播体散布出去。孢子形成之后发育出含孢粉素(sporopollenin)的孢子壁,而孢粉素的形成是对传播的高度适应,因为它能抵抗传播中机械的、化学的和紫外光的伤害[1]。种子植物在有性生殖或无融合生殖完成之后发育出种子,并以种子作为传播体,其中被子植物还能以果实作为传播体,实现植物的位移。
“果实和种子是植物的散布器官”[5]。传播、散布或扩散不是生殖过程,而是生态学过程[22~23]。传播方式的多样性体现了植物生态策略的复杂性[24]。生殖往往伴随增殖,而增殖之后,扩散就成为必然。扩散的意义在于:1)减少种内竞争;2)可在种群间形成基因流,扩大种群遗传多样性,增强种群生存力;3)扩大种群数量而降低物种灭绝风险;4)进入新生境,利于新物种形成。
作为传播体,种子较孢子有更多优势:1)孢子通常是一个细胞,其到达新环境中开始发育,要经历一个较脆弱的早期发育,而种子已是一个幼小的植物体,对严酷环境的抵抗力比由单细胞发育的个体最早期阶段强许多;2)孢子萌发立即进入自养阶段,而种子萌发可借助胚乳或子叶里的营养,仍过着异养生活,对环境的依赖较小;3)孢子休眠能力总体上比种子差,在萌发时机的选择上,种子的优势十分明显。
将种子、果实形成及其功能与有性生殖捆绑在一起是不合理的。被子植物的有性生殖即双受精在进化中没有显著的变化,可是由子房发育而成、为种子服务(保护种子、帮助种子传播)的果实,以及子房中由受精卵发育形成的种子,在形态结构和生理上则表现出极其复杂的多样化[2~13,19~20]。我们不能因种子、果实的巨大多样性而认为被子植物有性生殖具有丰富的多样性。被子植物有性生殖完成后,种子、果实发育的许多生长活动都是在为扩散准备条件,如:种皮、果皮产生可借助风播的翅、毛;种皮肉质化或形成假种皮、果皮肉质化。此外,许多菊科植物的花萼变态成毛状、具倒钩的刺毛等,以适应果实的传播;这些变态的花萼即冠毛是在花发育过程中形成的,与种子和果实的发育没有关系,更不受有性生殖的影响。所以,扩散和生殖是两个不同但相互关联的生物学现象。孢子、种子和果实的传播不仅影响种群动态,也影响群落演替。
总之,需要明确指出、反复强调的是:1)种子的形成不是生殖的结果,而是孢子体的早期发育(从受精卵到幼小植物体——胚);2)种子形成的生物学意义,不是生殖或繁殖,而是对扩散的生态适应;3)高等植物中的孢子植物和种子植物都产生孢子,二者的区别在于前者尚未进化出种子,通过孢子传播,后者的孢子原位发育[17],进化出更优越的传播体——种子。
此外,各教材常将种子植物的生活史或个体发育定义为“从种子到种子”[2,7,9~10,20],这值得质疑。将植物的一生的起点确定为种子显然“民科”色彩太浓,可能是受人类农业生产活动(播种和收获)的影响。讨论高等植物的一生、生殖和生殖器官,都要从孢子体世代和配子体世代两方面分别论述,不能用笼统的说法削弱植物学的科学性和严谨性。种子植物孢子体一生的起点是合子[8]。
动物胎生(vivipary)一词与卵生(ovipary)对应,是指受精卵在母体内发育成胎儿。如此看来,高等植物的受精卵都在母体内发育成胚、甚至成为幼小植物体,就是胎生现象。将一些红树林植物的种子传播之前就萌发的现象称为胎生[25]似有不妥,称为胎萌(embryo germination in situ,即胚的原位萌发,可缩写为EGIS)则较为恰当。红树植物的胎萌是对潮间带这一特殊生境中果实传播及传播后早期生存的适应[25]。由于植物“胎生”通常被看成一种繁殖方式[25],故在此提及,稍作澄清。
4 什么是生殖(繁殖)器官?
植物学教材中,被子植物的器官一般被分为六大类,即根、茎、叶、花、果实、种子,前三者被称为营养器官,后三者为生殖或繁殖器官[2,5~8,19~20]。
Mauseth[12]指出,生殖器官是参与有性生殖和无性生殖的器官,由一系列组织构成,如花、孢子叶球、精子器和颈卵器。由此看来,种子和果实都不是生殖器官。
那么,轮藻、苔藓和蕨类植物的生殖器官是什么?轮藻无世代交替,其植物体(1N)行有性生殖的生殖器官是精囊球(藏精器)和卵囊球(藏卵器)[2,6](表1)。苔藓和蕨类植物的配子体有雄性生殖器官精子器和雌性生殖器官颈卵器[2,6];苔藓孢子体的生殖器官是孢子囊,即孢蒴;蕨类孢子体的生殖器官是孢子囊,其见于枝顶的孢子囊穗或孢子叶、大型叶(fronds)羽片背面。
裸子植物的生殖器官与苔藓、蕨类的既有相似,又有不同。裸子植物的雌配子体的生殖器官,大多数为颈卵器(表1),买麻藤纲的买麻藤目(Gnetales)和百岁兰目(Welwitschiales)没有颈卵器[2,6];买麻藤目的雌配子体的生殖器官退化为一些游离核,百岁兰目雌配子体的生殖器官发育成胚囊管之类的结构[2]。裸子植物的雄配子体(花粉、花粉管)的生殖器官高度退化,其精子器退化为管细胞和生殖细胞,生殖细胞后来分裂为二精子[6,20]。裸子植物孢子体的生殖器官是大、小孢子囊,它们分别是大孢子叶上胚珠的珠心和小孢子叶背面的小孢子囊[2]。
将花说成是被子植物的有性生殖器官,这有些笼统。花是被子植物与生殖密切相关的器官。严格说来,雌蕊中的胚珠之珠心是大孢子囊,雄蕊的药囊是小孢子囊[6],这大、小孢子囊是孢子体的生殖器官。被子植物的配子体中,雌配子体的卵器(两个助细胞和1个卵细胞)是其雌性生殖器官,由颈卵器退化而成(表1);花粉粒、花粉管中的1个营养细胞和两个精子构成一个雄性生殖单位,即为雄性生殖器官(表1)。简言之,被子植物的孢子体器官可分为3类:1)孢子体的营养器官包括根、茎、叶;2)孢子体行无性生殖的生殖器官是花中的珠心和药囊[6,8](表1)。胚珠是大孢子囊[6]的说法是不准确的。无性生殖的无性和雌蕊的雌性、雄蕊的雄性纠缠在一起,常常令人费解。无性生殖的无性指在雌、雄蕊中未经两性细胞的融合即产生新个体(雌、雄配子体)的过程,雌、雄蕊直接参与的生殖是孢子体的无性生殖;而雌蕊、雄蕊二名称中的“雌、雄性别差异”包括了四层含义:孢子体器官雌/雄蕊的差异、雌/雄蕊产生大/小孢子、孢子原位发育成雌/雄配子体、配子体产生雌/雄配子。笔者认为,雌、雄蕊没有性别,真正有性别的是寄生于其内的配子体以及后者产生的配子;3)种子和果实是传播体。被子植物配子体行有性生殖的雌、雄生殖器官分别是卵器和雄性生殖单位[6]。
以上论述采取了严格的生殖器官概念,这是否有必要?当然有必要。植物学中的许多概念过于模糊,降低了植物学的科学性;植物学的科学性、严谨性和准确性必须是首要的,不能让步。严格的生殖器官概念是否会增加教学的难度?用准确的科学概念取代模糊的说法,会让学生欣赏到科学之美,而不是让学生陷入混乱和矛盾之中,这是必要的、值得的。如果采取严格的生殖器官的概念,花是繁殖器官的说法是否应保留?花作为无性世代孢子体的无性生殖器官的载体,其两性、单性如何理解?这些问题可以进一步开展讨论。
植物的生殖与动物的生殖有很大差别,而对于植物生殖的理解易受动物生殖模式的干扰;在定义生殖方式时,学者门往往容易忽略世代交替中不显著的植物体阶段。人类在农业生产活动中形成的传统观念有时也阻碍我们正确理解植物的生殖,如“春种一粒粟,秋收万颗子”的说法,易引导人们将播种这个传播(扩散)行为等同于繁殖,将种子当作生殖器官。在生殖概念上的混乱不易澄清,植物学教材的编者们需要不懈地开展学术探讨。
还有一个棘手的问题是,当研究苔藓植物的论文[14~15]都一致地将孢子生殖纳入有性生殖、将营养繁殖当作无性生殖时,若接受本文关于苔藓植物生殖的观点,是否会被学术界排斥?看来厘清生殖概念之路还会漫长而曲折。
