河西走廊几种荒漠植物种子萌发特性研究
2022-05-14鲁延芳占玉芳滕玉风甄伟玲
鲁延芳, 占玉芳, 滕玉风, 甄伟玲
河西走廊几种荒漠植物种子萌发特性研究
鲁延芳, 占玉芳, 滕玉风, 甄伟玲
张掖市林业科学研究院, 甘肃张掖 734000
为了解植物种子萌发适应干旱环境的基本特性, 以河西走廊人工梭梭林中重要值较大的梭梭()、红砂()、多枝柽柳()、雾冰藜()、沙蓬()、刺沙篷()、蒙古虫实()、白茎盐生草()、盐爪爪()、花花柴()等10种植物种子为对象, 对其萌发率、萌发开始时间、平均萌发时间和萌发持续时间进行研究。结果表明: (1)10种荒漠植物种子在Log尺度下均呈正态分布,种子大小与萌发持续时间、萌发开始时间和平均萌发时间之间都呈显著正相关(<0.05), 而与萌发率呈极显著负相关(<0.01)。(2)萌发率正态分布曲线呈现单峰型, 除沙蓬、刺沙篷外, 其余8种植物种子萌发率大于20%, 说明荒漠植物具有较高的萌发特性;萌发开始时间呈单峰型, 1—3 d内开始萌发的植物占90%, 说明荒漠种子萌发时间较短; 持续萌发时间呈单峰型, 梭梭持续萌发时间最短; 平均萌发时间呈单峰型,红砂平均萌发时间最长。(3)聚类分析显示, 10种植物种子萌发特性分为快速萌发型和缓慢萌发型, 主要受外界环境长期适应和选择的结果, 也是荒漠植物生存策略的重要环节。
河西走廊; 荒漠植物; 人工沙漠林; 萌发特性
0 前言
河西走廊干旱荒漠区是气候条件最严酷脆弱的地区之一, 也是气候变化和人类活动最敏感的地区, 这种恶劣生境条件对植物的生长、存活极为不利[1]。荒漠植物在严酷的生境下进化出了与之相适应的种子萌发对策来确保种子在适宜的时间和空间萌发[2]。种子萌发是植物生活史中至关重要的一步, 是决定种群动态的关键因子, 它影响到幼苗的建植、存活和竞争[3-5]。研究植物的分布、丰富度、种群动态以及植被更新过程首先要研究的就是种子萌发行为[6]。种子萌发行为的研究对探讨植物进化和未来的发展变化具有重要意义[7]。
干旱荒漠区通过大面积人工造林进行植被恢复,河西走廊民勤人工梭梭林就是治理荒漠化的有效手段, 但是种群更新过程中大多数种子在萌发和幼苗发育阶段失去活力。因此, 探讨种子的萌发特性是目前研究的热点问题。王梦茹和魏岩[8]研究了北疆荒漠心叶驼绒藜()种子的萌发行为,探讨了最适萌发温度及低盐胁迫对种子萌发的影响;刘会良等[9]探讨了古尔班通古特沙漠南缘32种藜科植物种子萌发策略,研究了32中藜科植物萌发特性;张颖娟等[10]在西鄂尔多斯研究了不同土壤水分处理对4种荒漠种子萌发和幼苗生长的响应,研究了土壤水分处理对荒漠种子萌发和幼苗生长的影响。但尚未见对河西走廊沙漠人工梭梭林中优势种种子萌发行为的报道。因此,本文选取河西走廊中部沙漠人工梭梭林下10种主要植物为对象,研究不同种子萌芽特性,探讨荒漠植物生长发育和生态适应性机制,为荒漠区植被的恢复与重建、生态修复、荒漠造林后的水分管理、维持生态系统稳定等提供科学依据。
1 材料和方法
1.1 研究区概况
研究区甘肃省民勤县位于102°02′—104°02′E, 38° 05′—39°06′N, 地处河西走廊东北部, 石羊河流域下游, 全县总面积1.42 万km2, 绿洲面积占总面积的8.9%, 东面、西面和北面被腾格里和巴丹吉林沙漠包围。气候属典型的大陆性荒漠气候, 其特点为: 气候干燥、降水稀少、蒸发强烈、风大沙多、气候环境十分恶劣; 多年平均气温7.6 ℃, 年平均日温差为15.2 ℃, ≥10 ℃积温为3036.4 ℃, 日照时数为3073.5 h, 无霜期176 d。多年平均降水量115.41 mm, 主要集中在每年的7—9月, 占年均总降水量的60%以上; 多年平均蒸发量2664 mm, 是降水量的24倍之多, 造成大面积土壤盐渍化。多年平均风速2.5 m·s-1, 年≥8级大风日数为27.8 d。研究区地下水位在12 m以下, 植物生长和存活完全依靠大气降水。风沙危害是当地主要的自然灾害之一, 全年盛行西北风、西北偏西风, 是全国浮尘、扬沙、沙尘暴发生最严重地区之一, 也是我国沙尘暴的主要策源地之一。自上世纪70年代末以来, 早期营造的梭梭林生长开始衰败, 甚至死亡, 防风固沙效能减弱, 植被大面积退化和衰败。本研究选取河西走廊中部沙漠人工梭梭林下重要值较高的 10 种占优势种的植物作为研究对象, 于成熟季2018年7—10月底采集。采集后放在通风条件下晾干后轻轻揉搓去壳去杂质, 用风选机进行风选, 选取完整籽粒饱满的种子在室温条件下储藏进行发芽试验。
1.2 研究方法
重要值=(相对多度+相对高度+相对盖度)/3 (1)
1.2.1 种子的重量的测定
用电子精密天平(精确至0.0001 g)对种子重量进行称重, 称重时种子带附属物包括绢毛、绒毛、翅等, 因为这些附属物具有植物传播价值[11]。每个物种测定100粒风干繁殖体的重量, 10次重复。
1.2.2 萌发试验
2018年将风干后挑选干净的种子置于化验室冰箱的冷藏中贮藏(温度3℃, RH:30%), 2019年4月在光照培养箱中进行萌发实验。首先在培养皿底部垫两层90 mm的滤纸, 用蒸馏水润湿后确保种子发芽所需的水分条件, 以滤纸润湿倒不出水为宜。将种子整齐摆放在口径为100 mm的培养皿中, 实验模拟春季气候和光照条件, 白天光照9000勒克斯, 时间12 h, 温度25 ℃; 夜晚光照0勒克斯, 时间12 h, 温度10 ℃。每个培养皿放100粒种子, 每个处理重复5次。实验开始后每天统计萌发种子数目, 种子发芽以胚根突破种皮达到种子长度的1/2为标准[12], 即可视为萌发, 同时每天10: 00开始记录种子萌发数量和霉变数量, 视情况补充水分, 持续观察30 d。对没有萌发的种子进行活力检测, 利用四唑(TTC)法对分拣出来的完好种子进行活力测定[13]。
1.3 萌发特性指标
萌发率=种子发芽数量/供试种子数×100 % (2)
萌芽时间是从萌发第一粒种子所需的时间(d)。萌发持续时间是种子开始萌发到结束所需的时间(d)。
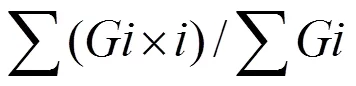
式中,表示从萌发实验开始后的萌发天数,表示第天的萌发个数。
发芽指数=1122+…+ G/X (4)
式中,为每天测定的发芽数,为连续测定相应天数。
1.4 数据处理方法
采用正态分布分析种子大小分布、相关性分析种子大小与萌发特性、主成分分析和聚类分析萌发特性, 运用Microsoft Excel 2007软件处理并进行图表制作, 使用Spss19.0软件进行数据分析和作图。
2 结果与分析
2.1 种子重量及大小的分布特征
共采集10种荒漠植物种子均属种子植物, 分3科10属(见表1)。根据研究区种子重要值(表1)可知, 灌木层中梭梭为优势种, 草本层中雾冰藜为优势种, 10种荒漠种子重量相对较大的种子出现的数量和频度总体高于重量相对较小的种子出现的数量和频度,这是由于荒漠植物种子千粒重相对较大的植物具有较高的结实率, 易于形成土壤种子库而且具有较强的时间拓殖能力[11], 可以更好地分散和占据新的栖息地[12]。而千粒重较小的种子在恶劣的气候环境下易被风吹, 不利于形成适合生长的环境。因此, 在荒漠植被恢复中, 采用飞播造林时选择重量相对较大的种子有利于植被的建植。
河西走廊人工沙漠梭梭林中10种种子百粒重如表1所示, 种子大小在Log尺度下呈正态分布如图1, 研究种子总体千粒重较小, 平均千粒重为2.376 g, 其中千粒重最大的是雾冰藜为6.0098 g, 最小的是多枝柽柳为0.15 g, 千粒重依次为雾冰藜()>红砂()>沙蓬()>刺沙篷()>梭梭()>蒙古虫实()>白净盐生草()>盐爪爪()>花花柴()>多枝柽柳()。
2.2 种子大小对萌发特性的影响
2.2 萌发特性
2.2.1 种子大小对萌发特性的影响
由表2可知, 种子大小与萌发率和开始持续时间呈显著负相关, 与萌芽指数、萌发时间和平均萌发时间呈正相关。说明荒漠种子千粒重相对越大萌芽时间和平均萌发时间越长, 萌发率越低, 萌发持续时间越短, 种子千粒重相对越小其萌发率越高, 萌芽时间和平均萌发时间越短, 理论上荒漠种子多以小种子为主的原因。然而千粒重较小的种子虽然具有较高的萌芽率, 但却不占优势种, 这可能是由于种子千粒重较小的植物易于被风吹, 不利于着床。因此, 荒漠植物种子以重量相对较大的植物为主, 是对外界环境长期适应和选择下形成的荒漠植物。

图1 种子大小分布频率
Figure 1 Frequency of seed size distribution

表1 河西走廊人工梭梭林10种荒漠植物种子萌发参数

表2 种子大小与萌发特性的方差分析
注: *表示差异显著(<0.05), **表示差异极显著(<0.01)。
2.2.2 萌发率
10种荒漠植物萌发率见图2, 梭梭萌发率高达90.5%; 其次是花花柴, 萌发率达84%; 依次是多枝柽柳, 萌发率达76.5%; 雾冰藜萌发率达74.25%; 红砂萌发率达72%; 盐爪爪萌发率达50.25%; 白茎盐生草萌发率达42.5%, 蒙古虫实萌发率达25%, 刺沙篷萌发率达14%, 刺沙篷由于薄膜革质包覆果实导致种子吸水困难萌发率最低为3%,因此在播种前应人为干预提高萌发率。萌发时间最短的是梭梭、雾冰藜、刺沙篷、白茎盐生草、盐爪爪, 都属于藜科, 具有萌发时间短, 萌发率高的特点。萌发率正态分布曲线如图2所示, 呈现单峰型, 萌发率大于20%的物种占80%, 说明荒漠植物在适宜的温度、水分、光照条件下具有较高的萌发特性。
2.2.3 萌发时间
萌发开始时间呈单峰型如图3所示, 10种植物中有9种植物在1—3 d内开始萌发如表1, 占了90%, 只有红砂萌发时间最晚是4 d, 说明荒漠种子具有较短的萌发时间, 是荒漠植物对环境高度适应的表现。荒漠地区降雨比较集中, 而且雨季较短, 加之风大, 地表蒸发强烈, 植物只能充分利用短暂的降雨时间, 迅速完成自己的生活周期, 繁衍后代[16]。

图2 萌发率的分布频率
Figure 2 The distribution frequency of germination rate
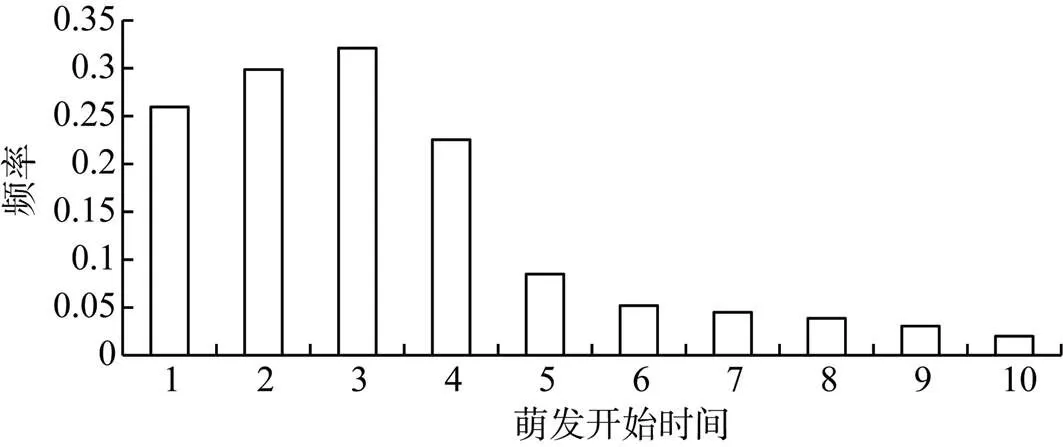
图3 萌发时间分布频率
Figure 3 Distribution frequency of germination onset time
2.2.4 萌发持续时间
持续萌发时间种间的差异较大如表1所示, 其中萌发持续时间小于10 d的物种只有一种是梭梭, 萌发持续时间在10—15 d的物种有花花柴、雾冰藜、刺沙篷、盐爪爪、多枝柽柳, 萌发时间在16—19 d的植物有蒙古虫实、沙蓬、白净盐生草, 萌发持续时间呈单峰型如图4所示。
2.2.5 萌发平均时间
平均萌发时间呈单峰型, 平均萌发时间最长的为红砂, 其次是梭梭和盐爪爪, 其它的平均萌发时间都很短, 说明荒漠植物种子发芽速度快, 发芽能力好, 在适宜的外界条件下就能迅速完成生活周期, 这主要受外界环境长期适应和选择的结果。
2.2.6 萌发指数
10种荒漠植物萌芽指数中萌芽指数较最高的是梭梭, 依次是多枝柽柳>花花柴>雾冰藜>白茎盐生草>红砂>盐爪爪>蒙古虫实>刺沙篷>沙蓬,与萌芽率呈正比。梭梭的发芽指数与其它植物发芽指数存在显著性差异(<0.01), 梭梭活力指数最高, 在说明梭梭种子生命力最强, 在人工治沙树种选择中可作为首选树种。
2.3 萌发特性主成分分析
对种子的4个萌发特性即萌发开始时间、萌发持续时间、萌发率和平均萌发时间进行主成分分析。通过初始特征值和提取载荷平方和表3, 主成分轴1代表10种荒漠植物种子萌发特性变异的45.7%, 主成分轴2代表10种荒漠植物种子萌发特性变异的33.7%。两个主成分轴用萌发特性变量的公式表示:
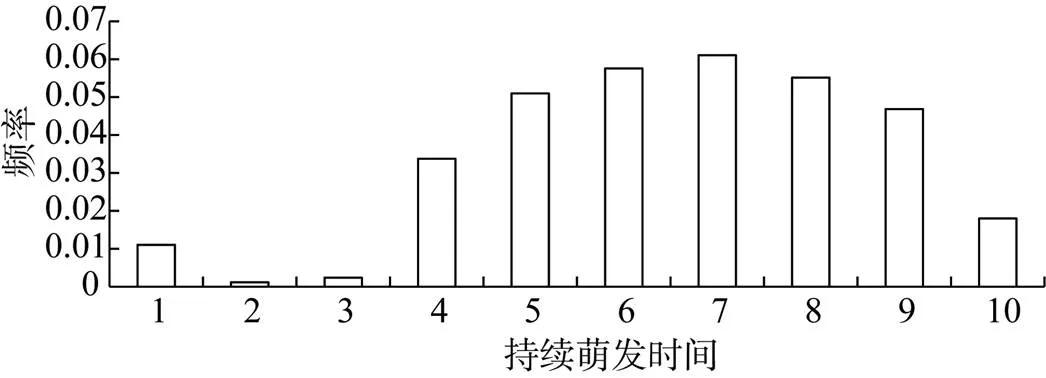
图4 持续萌发时间的分布频率
Figure 4 Distribution frequency of continuous germination time

图5 平均萌发时间的分布频率
Figure 5 Distribution frequency of mean germination time
主成分1=-0.125萌发开始时间-0.740萌发持续时间+0.868萌发率+0.714平均萌发时间
主成分2=0.907萌发开始时间+0.500萌发持续时间+0.139萌发率+0.508平均萌发时间
根据各物种在两个主成分轴上的聚类分析如图7所示, 10种荒漠植物有6种萌发类型属于快速发型, 即种子在浸润后几分钟或几个小时后能够迅速萌发, 分别是梭梭、花花柴、雾冰藜、蒙古虫实、白茎盐生草、盐爪爪等, 其中梭梭10 min就能够快速萌发并开始幼苗的生长, 且萌发具有聚集型, 这是干旱区遇降水后为保证幼苗生长并抵抗随之而来的干旱胁迫而快速萌发完成生活史, 是沙漠植物的一种优势。但是由于种子和幼苗生命阶段更容易受到缺水的影响, 因此有些植物为避免当少量降水后种子快速萌发而幼苗又没法抵抗随之而来的干旱而选择缓慢萌发, 其中有4种属于缓慢萌发型, 分别是红砂、沙蓬、刺沙篷、多枝柽柳等, 当降水量少, 种子通过萌发缓慢、萌发率低的策略适应严酷环境[17]。干旱胁迫下种子萌发率低是植物幼苗存活的策略之一, 缓慢萌发型是干旱区荒漠植物应对长期干旱或水资源短缺时期的应对机制。
3 讨论
由于受气候变化和人类活动的影响导致我国荒漠化面积迅速扩张、植被严重退化。干旱区通过大面积的人工造林来缓解植被退化, 但是播种建植成功率很低, 因此探索种子萌发特性, 对于植被恢复、更新和重建及维持生态系统的稳定和健康可持续发展具有重要的意义。

表2 主成分分析的总方差解释
荒漠植物千粒重较小的种子虽然具有较高的萌芽率, 但却不占优势种, 这可能是由于种子易于被风吹, 不利于着床, 荒漠植物种子以重量相对较大的植物为主。荒漠植物种子具有萌发时间短, 萌发率高的特点, 说明荒漠植物只要在短暂的降雨下就能在短期内完成生活史, 这主要是受外界环境长期适应和选择的结果。荒漠植物种子萌发类型分为快速萌发型和缓慢萌发型, 快速萌发的特性使得种子能够利用短暂有利的条件开始生活史[17], 能够在一系列栖息地压力下例如沙漠干旱年份常见的地区, 使幼苗死亡率降低[18]。但是有些种子为了避免幼苗受干旱胁迫死亡, 选择缓慢萌发, 对干旱具有不同的忍耐程度, 在种子萌发和幼苗生长早期阶段由于受干旱的影响造成干旱脱水, 当再次遇到降雨后又能恢复生长, 表现出对干旱环境的适应性进化特征, 这些都是荒漠种子为适应干旱, 强烈光照, 风蚀及营养贫瘠等沙漠条件而形成的许多旱生特征[19]。
荒漠植物在漫长的生态环境进化更新过程中形成了多样的萌发特性和生存机制来适应外界多变的环境胁迫, 以确保植物能够在适宜的时机和环境中进行种子的萌发与幼苗的建成。有些种子在不同时间的降雨后分批萌发, 减少了大部分幼苗同时受害的危险, 较容易实现种群更新和扩展, 其种子萌发策略有利于其在荒漠地区的广泛分布[20]。人工种子补播在一定程度上可以促进幼苗更新与植被恢复, 幼苗阶段抗逆性强和演替中后期的优势种可作为有效补播物种[21]。对于快速萌发型的物种结合早期有效降雨进行补播, 对于缓慢萌发型物种通过破除休眠的预处理进行补播。
荒漠植物种子结构与功能具有高度统一性, 也说明了生态环境对植物种子形态结构的进化有着深远的影响[17]。本研究只对萌发特性进行了分析, 对种子萌发机理、种子对盐分胁迫及种子附属物对种子萌发的影响及外界环境因子水分、温度、光照、氧气、风火等条件的影响还有待于进一步研究[22]。
4 结论
10种荒漠植物种子均属种子植物, 分3科10属, 种子大小在Log尺度下呈正态分布, 10种荒漠种子中平均千粒重为2.376 g, 其中千粒重最大的是雾冰藜为6.0098 g, 最小的是多枝柽柳为0.15 g, 群落多样性调查中, 梭梭-雾冰藜为优势群落, 说明荒漠植物种子中, 种子重量相对较大的种子具有优势, 然而在发芽试验中种子千粒重相对越小的种子其萌发率越高, 萌芽时间和平均萌发时间越短, 这是由于荒漠区降水稀少且集中, 荒漠植物具有萌发快, 萌发率高的特点。荒漠植物在外界环境长期自然选择下在少量的降水条件下短时间内完成生活史, 是荒漠植物对环境高度适应的表现。持续萌发时间呈单峰型, 梭梭持续萌发时间最短; 平均萌发时间呈单峰型, 平均萌发时间最长的为红砂。对萌发特性进行聚类分析, 分为快速萌发型和缓慢萌发型。在发芽试验中, 种子千粒重相对越大的种子虽然具有较小的萌发率, 如雾滨藜, 可能是由于具有较高的结实率, 且种子千粒重较大, 不易被风吹, 易于形成土壤种子库。因此, 在人工梭梭林群落多样性调查中具有较大的重要值。
[1] 张景光, 王新平, 李新荣, 等. 荒漠植物生活史对策研究进展与展望[J].中国沙漠, 2005, 25 (3): 306–314.
[2] GUTTERMANN Y. Seed Germination in Desert Plants[J]. Journal of Plant Physiology, 1995, 146(5/6):767–768.
[3] BASKIN C, BASKIN J M. Seeds: ecology, Biogeography, and evolution of dormancy and germination[M]. Academic Press, San Diego, 2014.
[4] GUTTERMANN Y. Long-term seed position influences on seed germinability of the desert annual,L[J]. Israel Journal of Plant Sciences, 1994, 42 (3): 197–205.
[5] HUANG Zhenying, GUTTERMANN Y. Artemisia monosperma achene germination in sand: effects of sand depth, sand/water content, cyanobacterial sand crust and temperature[J]. Journal of Arid Environments, 1998, 38: 27–43.
[6] 李珍, 云岚, 石子英, 等. 盐胁迫对新麦草种子萌发及幼苗期生理特性的影响[J]. 草业学报, 2019, 28 (8): 119–129.
[7] 张颖娟, 王玉山. 西鄂尔多斯4种荒漠植物种子萌发对水分条件的响应[J]. 水土保持通报, 2010, 30(6): 60–63.
[8] 王梦茹, 魏岩. 北疆荒漠心叶驼绒藜种子的萌发行为[J].草业科学, 2018, 35(8): 1836–1842.
[9] 刘会良, 宋明方, 段士民. 古尔班通古特沙漠南缘32种藜科植物种子萌发策略初探[J]. 中国沙漠, 2012, 32(2): 413–420.
[10] 张丽坤, 王朔, 冯玉龙. 紫茎泽兰种子形态特征和萌发特性与其入侵性的关系[J]. 生态学报, 2014, 34(13): 3584–3591.
[11] 李照青, 高郯, 张铎, 等. 西藏林芝高山松土壤种子库空间分布特征研究[J]. 西北林学院学报, 2019, 34(4): 43–48.
[12] MULLER-LANDAU H C. The tolerance-fecundity trade-off and the maintenance of diversity in seed size[J]. Proceedings of the National Academy of Sciences, 2010, 107 (9): 4242–4247.
[13] SKARPASS O, SILVERMAN E J, JONGEJANS E, et al. Are the best dispersers the best colonizers? Seed mass, dispersal and establishment in Carduus thistles[J]. Evolutionary Ecology, 2011, 25 (1): 155–169.
[14] 贺慧, 燕玲, 郑彬. 5种荒漠植物种子萌发特性及其吸水特性的研究[J]. 干旱区资源与环境, 2008, 22(1). 184–188.
[15] 曾彦军. 干旱荒漠区几种优势植物种子萌发生态学 研究[D]. 兰州: 兰州大学, 2010.
[16] 王东丽. 黄土丘陵沟壑区植物种子生活史策略及种子补播恢复研究[D]. 杨凌: 西北农林科技大学, 2014.
[17] 敖云娜. 干旱和盐胁迫对华北驼绒藜种子萌发及幼苗生长的影响[D]. 长春: 东北师范大学, 2019.
[18] GUTTERMANN Y. Survival Strategies of Annual Desert Plants [M]. Berlin, Springer, 2002.
[19] 王慧春, 周华坤, 王文颖, 等. 三江源区高寒草甸43种植物繁殖体质量和形状的比较[J]. 西北农林科技大学学报, 2009, 37 (8): 195–203.
[20] 黄振英, GUTTERMANN Y, 胡正海. 白沙蒿种子萌发特性的研究Ⅰ. 粘液瘦果的结构和功能[J]. 植物生态学报, 2001, 25 (1): 22–28.
[21] 刘志民, 李雪华, 李荣平, 等. 科尔沁沙地70种植物繁殖体形状比较研究[J]. 草业学报, 2003, 12 (5): 55–61.
[22] 刘莹, 麻文俊, 杨桂娟, 等. 砂生槐天然种群种实形态和萌发特征地理变异[J]. 西北林学院学报, 2019, 34(6): 22–29.
Study on seed germination characteristics of several desert plants in Hexi Corridor
LU Yanfang, ZHAN Yufang, TENG Yufeng, ZHEN Weiling
Zhangye Forestry, Science Academy, Zhangye Gansu 73400
In order to understand the basic characteristics of plant seed germination adapting to arid environment, the seeds of 10 plant species of greater importance in the artificialforest in Hexi Corridor were selected as objects, such as,,,,,,,,,. The germination rate, germination onset time, average germination time and germination duration were studied. The results showed that: (1) The 10 desert plant species showed a normal distribution on the Log scale, the seed size was significantly and positively correlated with germination duration, germination start time and average germination time, but was extremely significantly and negatively correlated with germination rate. (2) The normal distribution curve of germination rate showed a single peak, except forand, and species with a germination rate greater than 20% accounted for 80%, with high germination characteristics. The germination start time was single peak, and the plants that started germination accounted for 90% within 1-3 days, indicating that the germination time of desert seeds was shorter. The continuous germination time showed a single peak shape, and thehad the shortest continuous germination time. The average germination time showed a single peak, and the longest average germination time was. (3) The germination characteristics were classified into fast germination type and slow germination type by cluster analysis. It is the result of the long-term adaptation and selection of the environment, and is also an important part of the survival strategy of desert plants.
Hexi Corridor; desert plant; artificial desert forest; germination characteristics
鲁延芳, 占玉芳, 滕玉风,等. 河西走廊几种荒漠植物种子萌发特性研究[J]. 生态科学, 2022, 41(3): 222–228.
LU Yanfang, ZHAN Yufang, TENG Yufeng, et al. Study on seed germination characteristics of several desert plants in Hexi Corridor[J].Ecological Science, 2022, 41(3): 222–228.
10.14108/j.cnki.1008-8873.2022.03.026
S157.2
A
1008-8873(2022)03-222-07
2020-09-23;
2020-10-20
甘肃省科技计划资助(21JR7RG880)
鲁延芳(1982—), 女, 甘肃省永登人, 硕士, 高级工程师, 主要从事林业生态工程、荒漠化综合治理和生态恢复与重建研究,E-mail:lkylyf1014@126.com
鲁延芳
