脱毒对大蒜生长、碳氮代谢的影响
2022-05-13刘灿玉杨峰陆信娟赵永强张碧薇杨青青葛杰樊继德
刘灿玉 杨峰 陆信娟 赵永强 张碧薇 杨青青 葛杰 樊继德
摘要:以徐蒜917常规栽培种作为对照(CK),选用脱毒F1、F2、F3、F7代作为试验材料,研究脱毒对大蒜生长、碳氮组分含量及其关键酶活性的影响。结果表明,脱毒有利于提高大蒜农艺性状指标,且随繁殖世代的增加,其综合生长势呈先增后降的趋势,以F2代综合长势较好,F7代除假茎粗、叶长和叶宽仍显著高于CK外,其他与CK无显著差异。脱毒大蒜叶片色素含量随繁殖世代的增加呈下降趋势,且处理间差异在抽薹期大于鳞茎膨大期。在抽薹期和鳞茎膨大期,脱毒大蒜叶片总糖、还原糖和蔗糖含量均高于CK,蔗糖磷酸合成酶(SPS)活性亦显著增加,且均以F2代最高。随繁殖世代的增加,脱毒大蒜叶片蔗糖合成酶(SS)活性在抽薹期呈先降后升的趋势,而在鳞茎膨大期呈先升后降的趋势。脱毒有利于提高大蒜叶片中抽薹期和鳞茎膨大期NO-3、可溶性蛋白和游离氨基酸含量,硝酸还原酶(NR)、谷酰胺合成酶(GS)、谷氨酸合酶(GOGAT)活性显著增加,脯氨酸含量在鳞茎膨大期亦显著增加。综上所述,脱毒有利于大蒜生长,增强其碳氮代谢关键酶活性,促进碳氮吸收同化、糖的积累和蛋白质合成,但随着脱毒大蒜繁殖世代的增加,其生长优势会逐渐丧失。
关键词:脱毒大蒜;抽薹期;鳞茎膨大期;生长;碳代谢;氮代谢;碳氮代谢关键酶
中图分类号: S633.401 文献标志码: A
文章编号:1002-1302(2022)08-0141-04
大蒜属无性繁殖作物,主要通过分瓣或形成气生鳞茎的方式进行繁殖,母本鳞茎一旦被病毒侵入,就会导致病毒在大蒜植株体内世代积累[1-2]。随着多年种植,主产区大蒜病毒病日趋严重,危害程度日益加剧,已成为限制大蒜优质高产的主要因素之一。病毒病可造成大蒜品种退化,产量显著降低[3-4],严重时可造成大蒜僵苗不发,甚至绝产不收[5],同时会造成大蒜鳞茎整齐度差,进而影响大蒜商品性,降低其市场竞争力和经济效益。当前,培育和栽培无病毒种苗是防治病毒病的根本措施[6]。吴青青等认为,控制病毒病的传播目前最有效的做法是种苗脱毒[7]。赵硕等研究发现,脱毒大蒜原原种和原种的长势和鳞茎质量均显著好于常规大蒜种植[8];徐培文等研究发现,太仓大蒜脱毒后再次感染较慢,可在5代内仍保持高产[9];刘文英等研究表明,苍山大蒜脱毒后种至第4代,仍有显著增产效果,经济效益明显,脱毒原种在产区可利用至第5代[10]。与常规栽培种相比,大蒜脱毒后长势旺盛,可显著提高其产量和品质,但目前大多数报道仅局限于观察、比较脱毒大蒜的增产效果,而对于脱毒影响大蒜生长的生理机制的研究较少。碳氮代谢作为植株最基本、最重要的生理代谢过程[11],其在作物生育期间的动态变化直接影响光合产物的形成、转化及蛋白质的合成等过程,对植物正常生长发育有着非常重要的影响[12]。本试验研究脱毒对大蒜生长、碳氮组分含量及其关键酶活性的影响,以期为深入研究脱毒大蒜增产机理提供依据。
1 材料与方法
1.1 试验材料与试验设计
试验在江苏徐淮地区徐州农业科学研究所试验示范基地进行,脱毒917大蒜由茎尖脱毒组培技术获得试管苗(已病毒检测为无病毒株),然后驯化获得原原种,即为脱毒F1代,由原原种做种蒜获得原种,即为脱毒F2代,以此类推。分别选择脱毒F1、F2、F3、F7代作为试验材料,以917常规栽培种作为对照(CK),于2019年10月4日播种于网室中,并加盖防虫网,每个试验材料重复3次,随机区组排列,每个小区面积为20 m2,其他栽培方式按照徐州当地高产栽培水平进行。
1.2 测定项目与方法
根据《大蒜种质资源描述规范和数据标准》[13]调查大蒜农艺性状。分别于抽薹期(2020年4月7日)、鳞茎膨大期(2020年5月7日)取大蒜叶片,用液氮速冻后放于-80 ℃冰箱中,用于指标测定。采用乙醇提取法测定叶片色素含量[14];硝态氮(NO-3-N)含量的测定采用赵世杰的方法[14];参考Saladin等的方法测定铵态氮(NH+4-N)、游离氨基酸和游离脯氨酸含量[15];采用考马斯亮蓝法测定可溶性蛋白含量[16];总糖和蔗糖含量采用蒽酮比色法测定[17];还原糖含量的测定采用3,5-二硝基比色法[18];硝酸还原酶(NR)活性的测定采用活体法[14];参照Wang等的方法测定谷酰胺合成酶(GS)活性[19];参照Groat等的方法测定谷氨酸合酶(GOGAT)活性[20];蔗糖磷酸合成酶(SPS)、蔗糖合成酶(SS)活性参照高俊凤的方法测定[18]。
1.3 数据处理
分别采用Microsoft Excel 2003、DPS 7.5软件进行试验数据处理和处理间差异显著性检验(Duncans新复极差法)。
2 结果与分析
2.1 脱毒对大蒜农艺性状的影响
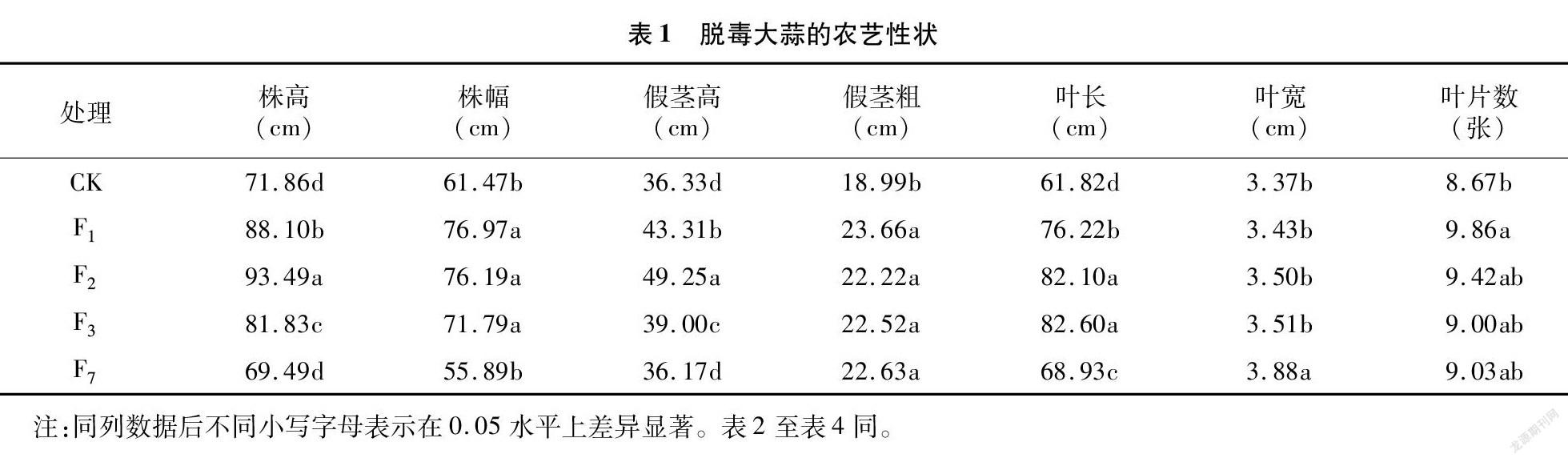
由表1可知,脱毒有利于提高大蒜农艺性状指标。随繁殖世代的增加,脱毒大蒜株高、假茎高及叶长均呈先增后降的趋势,大体以F2代最好,F1代次之,F7代仅叶长仍显著高于CK,其株高和假茎高与CK无差异。脱毒大蒜处理组间株幅呈下降趋势,且F7代与CK无差异,显著低于其他世代。脱毒大蒜间假茎粗无差异,均显著高于CK。脱毒大蒜间叶宽以F7代最宽,显著高于其他世代。脱毒大蒜叶片数仅F1代显著高于CK,其他与CK无显著差异。
2.2 脱毒对大蒜叶片色素含量的影响
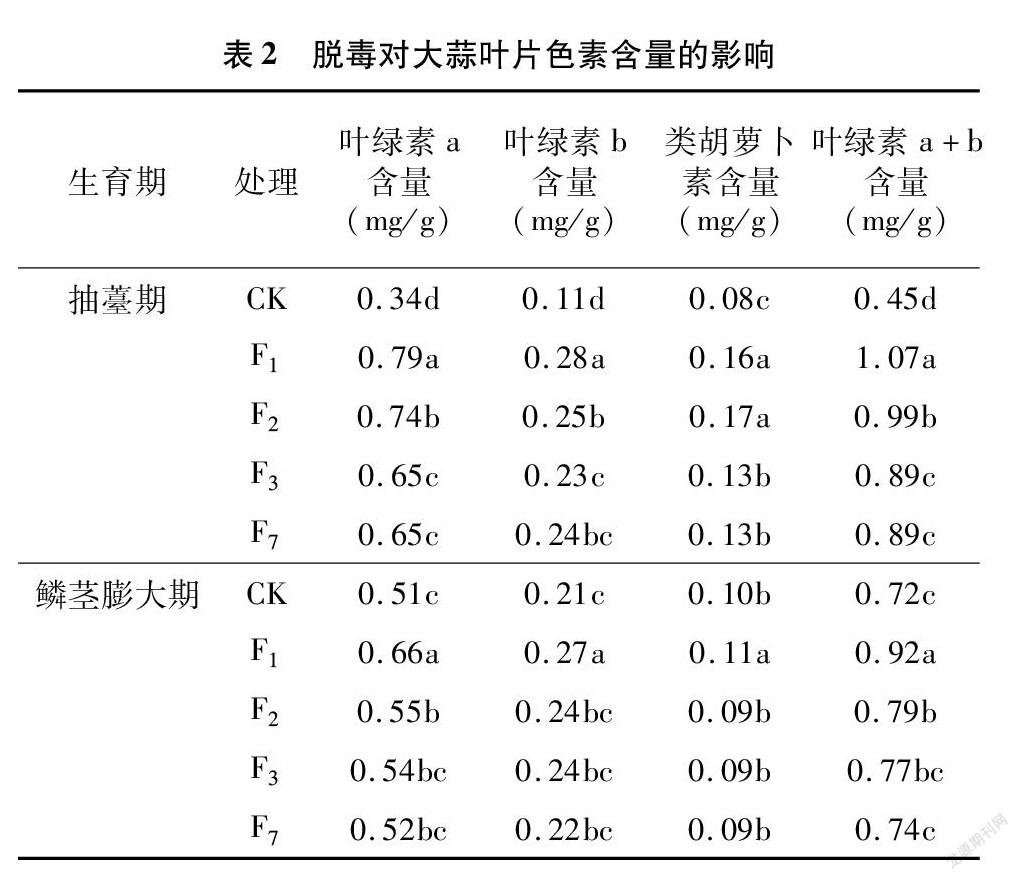
由表2可知,脱毒有利于提高大蒜葉片色素含量。在抽薹期,与CK相比,脱毒大蒜叶绿素a、叶绿素b、类胡萝卜素含量均显著增加,但随繁殖世代的增加,脱毒大蒜的叶片色素含量呈下降趋势,叶绿素a和b均以脱毒F1代含量最高,F2代次之,F3代与F7代差异不显著,类胡萝卜素含量在F1代和F2代间无差异,显著高于F3代和F7代。至鳞茎膨大期,脱毒大蒜叶片色素含量随繁殖世代的增加亦呈下降趋势,但仅F1代显著高于其他世代。
2.3 脱毒对大蒜叶片糖含量及酶活性的影响
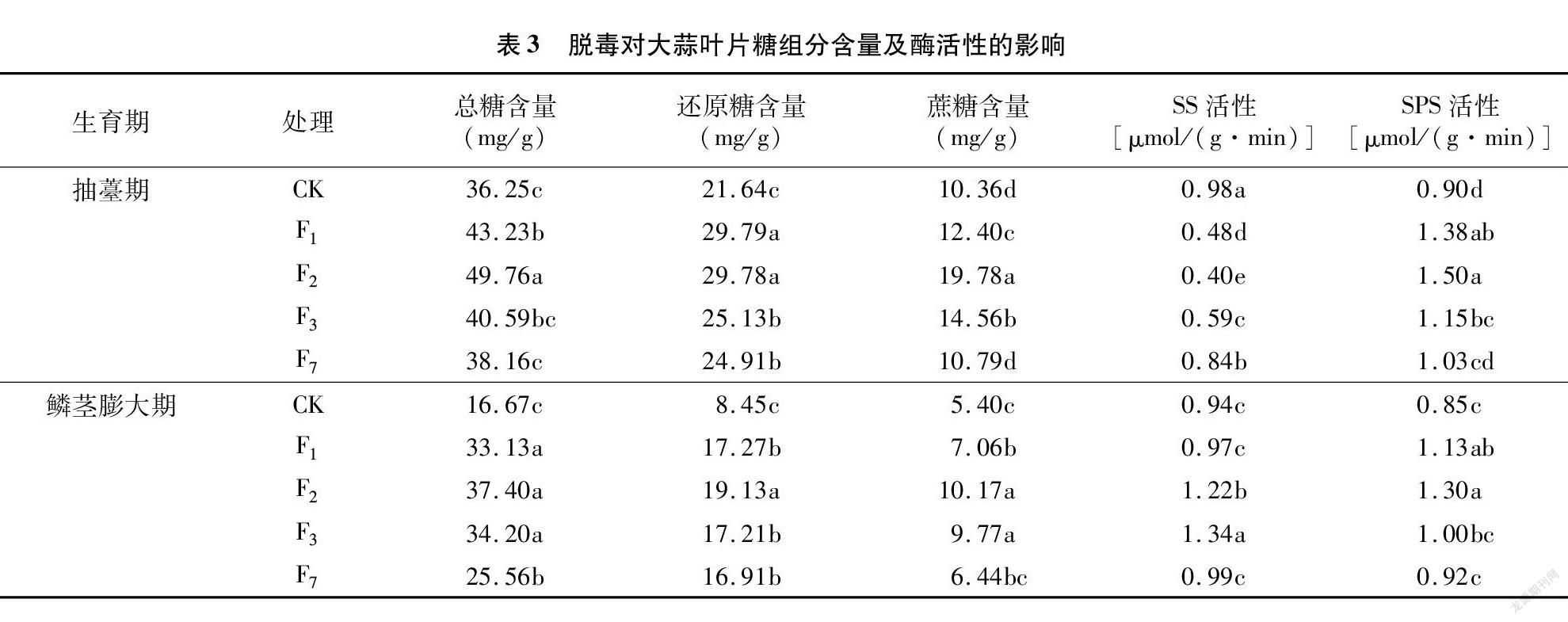
由表3可知,与CK相比,脱毒有利于提高大蒜叶片糖含量。在抽薹期,随繁殖世代的增加,脱毒大蒜叶片总糖、蔗糖含量及SPS活性均呈先增后降的趋势,且均以F2代最高,而SS活性呈先降后升的趋势,以脱毒F2代活性最低,与此同时,还原糖含量呈下降趋势,脱毒F1代和脱毒F2代差异不显著。鳞茎膨大期的大蒜叶片总糖、蔗糖含量及SPS活性等的变化趋势与抽薹期基本一致,但SS活性与之相反,呈先升后降的趋势,以脱毒F3代活性最高,脱毒F2代次之。
2.4 脱毒对大蒜叶片氮代谢相关物质含量及酶活性的影响
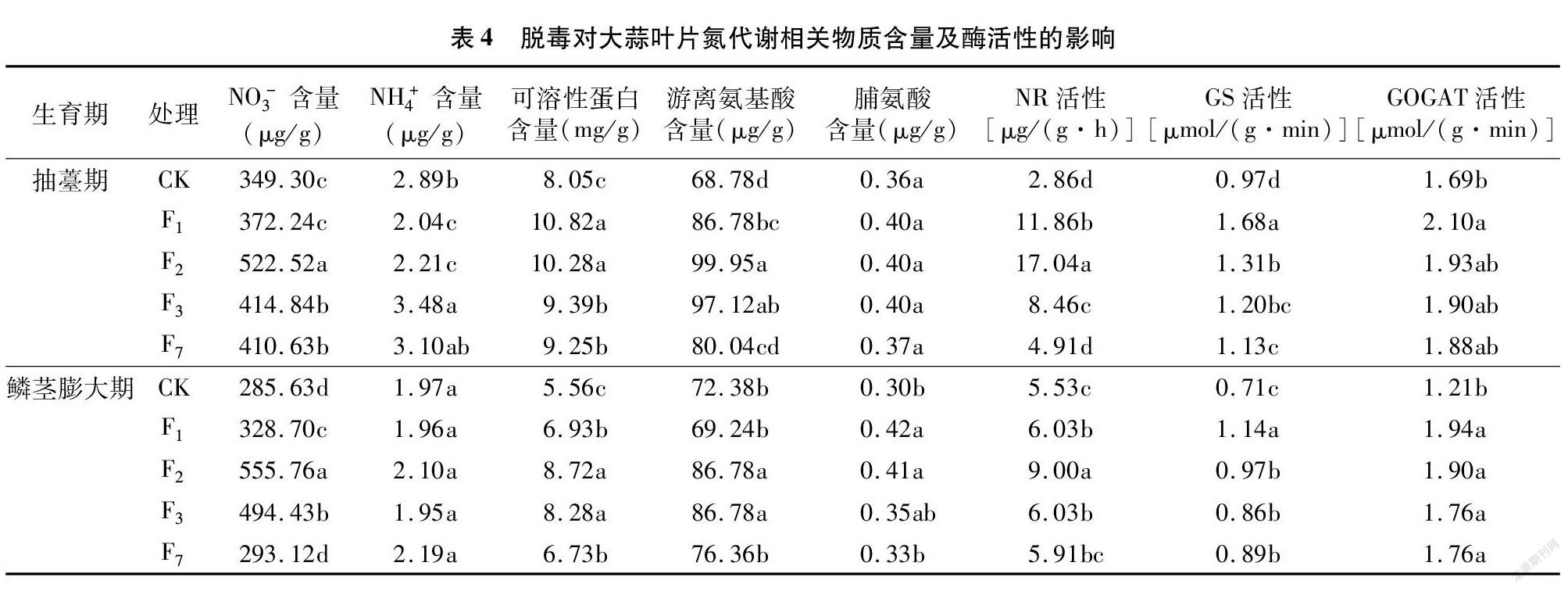
由表4可知,与CK相比,脱毒有利于提高大蒜叶片的NO-3、可溶性蛋白、游离氨基酸及脯氨酸含量。抽薹期,随繁殖世代的增加,脱毒大蒜叶片NO-3、游离氨基酸含量及NR活性均呈先增后降的趋势,以脱毒F2代含量最高;与此同时,NH+4含量呈增加趋势,以脱毒F3含量最高;而可溶性蛋白含量及GS、GOGAT活性均呈下降趋势,以脱毒F1代最高,脱毒F7代最低,且均显著高于CK;各处理间脯氨酸含量差异不显著。
至鳞茎膨大期,各处理间大蒜叶片NH+4含量无显著差异;可溶性蛋白和游离氨基酸含量均随繁殖世代的增加呈先增后降的趋势,以脱毒F2代含量最高,脱毒F1代和脱毒F7代间差异不显著;脯氨酸含量呈下降趋势,以F1代最高;大蒜叶片NO-3含量及NR、GS、GOGAT活性变化趋势与抽薹期基本一致。
3 讨论与结论
前人研究认为,无性繁殖作物在长期生产上易受到病毒侵染和积累[4,21],可导致植株生长势减弱,叶片卷曲或坏死,最终导致产量和品质下降[22-23]。目前,种苗脱毒是减少和防止病毒造成损失的最有效方法[24-25]。蒋明权研究发现,甘薯脱毒后综合生长势优于常规栽培种[26]。本试验结果表明,大蒜脱毒有利于提高其生长势,且以脱毒F2代综合生长势较好,这可能是因为脱毒F2代无病毒侵染或再次侵染后病毒积累尚较少,且繁殖营养体较大利于后期营养生长[27]。随着繁殖代数的增加,脱毒F7代的生长势已逐渐趋于常规栽培种,表明脱毒大蒜的生长优势会因病毒的再侵染和积累而逐渐丧失。
已有研究表明,植物被病毒侵染会造成其叶绿体结构变化,功能遭到破壞,影响碳同化和代谢[28]。蒋明权研究发现,种苗脱毒可提高甘薯整个生育期的叶绿素含量[26]。在本试验中,大蒜脱毒后叶片色素含量显著增加,这与刘文英等的研究结果[10]一致,表明种苗脱毒可避免病毒对大蒜叶绿体结构和功能的不良影响,从而为增强碳代谢提供基础,但随着繁殖世代的增加,叶片色素含量开始下降,这可能是由病毒的再侵染和积累造成的。目前有关脱毒对大蒜碳代谢影响的研究尚有欠缺,本试验对此进行了初步研究,结果发现,脱毒大蒜总糖、还原糖和蔗糖含量在抽薹期和鳞茎膨大期2个生育期内均较高,且以F2代最高,SPS活性亦显著增加,表明脱毒可提高大蒜叶中糖的合成代谢。此外,脱毒大蒜叶片SS活性在抽薹期低于常规栽培种,而在鳞茎膨大期反而上升,表明在抽薹期,大蒜脱毒在促进叶片碳同化和转化的同时还降低了糖的分解代谢;而在鳞茎膨大期,脱毒大蒜叶片中糖的分解代谢增强,这可能与光合产物从叶中向地下鳞茎输出有关。
前人研究发现,脱毒植株氮代谢活跃,叶片NR活性增强[26],游离脯氨酸含量增加[24]。本试验中,抽薹期和鳞茎膨大期脱毒大蒜叶片中NO-3含量和NR活性均显著增加,至F7代时与常规栽培种差异逐渐变小,说明脱毒可增强植株对NO-3的吸收和同化能力,但随繁殖世代的增加,病毒的再侵染和积累会导致其能力下降。与此同时,脱毒大蒜叶片中可溶性蛋白和游离氨基酸含量亦显著增加,这可能与脱毒诱导了GS和GOGAT活性增加有关。脱毒F1代和F2代鳞茎膨大期脯氨酸含量明显高于常规栽培种,而较高的脯氨酸含量可提高大蒜耐高温能力[29],避免此生育期内高温引起的大蒜早衰,这或许是脱毒大蒜增产的原因之一。
在本试验中,脱毒可提高大蒜长势,叶绿素含量亦显著增加,碳氮代谢关键酶活性增强,利于糖分积累和NO-3吸收同化,但随着繁殖世代的增加,病毒的再侵染和积累可导致生长优势下降,种性退化。
参考文献:
[1]高山林,金雍安,蔡朝晖,等. 大蒜分生组织培养脱病毒和快速繁殖技术[J]. 植物资源与环境学报,2000,9(3):15-18.
[2]陈世儒,黄菊辉.大蒜离体快繁及脱毒[J]. 园艺学报,1991,18(3):245-250.
[3]李金娟. 大蒜病毒病原的分子鉴定与序列分析[D]. 兰州:甘肃农业大学,2011.
[4]Fan B L,He R F,Shang Y T,et al. System construction of virus-free and rapid-propagation technology of Baodi garlic (Allium sativum L.)[J]. Scientia Horticulturae,2017,225:498-504.
[5]高 畅,杨 飞,沈若刚,等. 嘉定白蒜脱毒微鳞茎诱导的快繁技术研究[J]. 中国蔬菜,2019(3):66-72.
[6]王 涛. 大蒜茎尖脱毒技术研究及酶活性分析[D]. 大连:大连工业大学,2013.
[7]吴青青,王维泽,崔 嵬,等. 百合茎尖培养材料的筛选及其组培配方的优化[J]. 贵州农业科学,2019,47(9):69-73.
[8]赵 硕,黄丹枫,高 畅,等. 组培脱毒嘉定白蒜原原种生产的技术关键点试验研究[J]. 中国蔬菜,2019(10):65-71.
[9]徐培文,孙慧生,孙瑞杰,等. 大蒜茎尖培养脱毒及增产效果的研究[J]. 山东农业科学,1991,23(6):6-10.
[10]刘文英,李 辉,张凤伦,等. 脱毒苍山大蒜增产效果及生物学基础[J]. 华北农学报,1998,13(4):74-79.
[11]李 祎,杨顺瑛,郝东利,等. 利用gs1.1和gs1.2突变体研究外源蔗糖对高铵胁迫拟南芥碳氮代谢的影响[J]. 土壤,2020,53(1):21-29.
[12]张建新,葛淑芳,吴玉环,等. 干旱胁迫对紫金牛叶片碳氮代谢的影响[J]. 水土保持学报,2015,29(2):278-282.
[13]李锡香,朱德蔚. 大蒜种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2006.
[14]赵世杰. 植物生理学实验指导[M]. 北京:中国农业科学技术出版社,2002.
[15]Saladin G,Magné C,Clément C.Stress reactions in Vitis vinifera L. following soil application of the herbicide flumioxazin[J]. Chemosphere,2003,53(3):199-206.
[16]王月福,于振文,李尚霞,等. 氮素營养水平对冬小麦氮代谢关键酶活性变化和籽粒蛋白质含量的影响[J]. 作物学报,2002,28(6):743-748.
[17]高 松,刘 颖,刘学娜,等. 光质对大葱叶片碳氮代谢的影响[J]. 植物生理学报,2020,56(3):565-572.
[18]高俊凤. 植物生理学实验指导[M]. 北京:高等教育出版社,2006.
[19]Wang L,Zhou Q X,Ding L L,et al. Effect of cadmium toxicity on nitrogen metabolism in leaves of Solanum nigrum L. as a newly found cadmium hyperaccumulator[J]. Journal of Hazardous Materials,2008,154(1/2/3):818-825.
[20]Groat R G,Vance C P.Root nodule enzymes of ammonia assimilation in alfalfa (Medicago sativa L.) developmental patterns and response to applied nitrogen[J]. Plant Physiology,1981,67(6):1198-1203.
[21]Dijk P. Carlavirus isolates from cultivated Allium species represent three viruses[J]. Netherlands Journal of Plant Pathology,1993,99(5/6):233-257.
[22]Pradhan S,Regmi T,Ranjit M,et al. Production of virus-free orchid Cymbidium aloifolium (L.) Sw.by various tissue culture techniques[J]. Heliyon,2016,2(10):176.
[23]Ling K S,Jackson D M,Harrison H,et al. Field evaluation of yield effects on the USA heirloom sweetpotato cultivars infected by sweet potato leaf curl virus[J]. Crop Protection,2010,29(7):757-765.
[24]陈选阳,陈凤翔,袁照年,等. 甘薯脱毒对一些生理指标的影响[J]. 福建农业大学学报,2001,30(4):449-453.
[25]沈升法,吴列洪,项 超. 甘薯脱毒微型薯育苗技术研究[J]. 江苏农业科学,2020,48(5):97-100.
[26]蒋明权. 甘薯茎尖脱毒、快繁技术及其脱毒苗增产机理的研究[D]. 合肥:安徽农业大学,2004.
[27]严根元,顾丁元. 种瓣大小对大蒜生长发育及产量的影响[J]. 上海蔬菜,1990(3):29.
[28]Lehrer A T,Moore P H,Komor E.Impact of sugarcane yellow leaf virus (ScYLV) on the carbohydrate status of sugarcane:comparison of virus-free plants with symptomatic and asymptomatic virus-infected plants[J]. Physiological and Molecular Plant Pathology,2007,70(4/5/6):180-188.
[29]邱念伟,杨翠翠,付文诚,等. 高盐和高温胁迫下外源脯氨酸对PSⅡ颗粒的保护作用[J]. 植物生理学报,2013,49(6):586-590.
