2株源于红树林具有黑曲霉抗性细菌的筛选及发酵条件优化
2022-05-13阮羽萱郑添鹏魏宝阳刘博宇阮颖黄勇
阮羽萱 郑添鹏 魏宝阳 刘博宇 阮颖 黄勇








摘要:黑曲霉(Aspergillus niger)是一种常见的病原微生物,极易造成粮食霉变、果实腐烂等问题,产生的毒素严重危害人体健康。红树林作为一种特殊的生态系统,具有重要的微生物学资源与经济价值。首先采用常规细菌筛选方法分离菌株,通过观察形态特征以及16S rRNA基因序列进行鉴定。然后通过单因素试验并结合正交试验确定菌株 Z-2001、R-2008生长的最佳培养条件,并通过拮抗试验确定Z-2001、R-2008菌株对黑曲霉的抑制效果。发现 Z-2001、R-2008菌株最佳基础培养基为营养琼脂培养基(NA)。Z-2001菌株最佳培养基配方为葡萄糖(2.0%)+蛋白胨(0.8%)+MgSO4·7H2O(0.30%)+牛肉膏(0.3%)+NaCl(0.5%),最佳pH值、温度、转速分别为8、37 ℃、210 r/min,黑曲霉抑菌率为76.80%。R-2008菌株最佳培养基配方为葡萄糖(2.0%)+酵母粉(2.4%)+MgSO4·7H2O(0.20%)+牛肉膏(0.3%)+NaCl(0.5%),最佳pH值、温度、转速分别为8、28 ℃、180 r/min,黑曲霉抑菌率为78.70%。结果表明,Z-2001、R-2008菌株能够有效抑制黑曲霉生长,且对生长环境以及条件要求并不严苛,具有较好的开发前景。
关键词:红树林;芽孢杆菌;抗性细菌;黑曲霉;培养基优化
中图分类号:S182;S188+.4 文献标志码: A
文章编号:1002-1302(2022)08-0133-08
黑曲霉(Aspergillus niger)是一种重要的植物病原菌,广泛分布于粮食、植物性产品和土壤中,极易大量繁殖导致粮食霉变。据联合国粮农组织(Food and Agriculture Organization of the United Nations,FAO)统计,全世界每年大约有3%的粮食因为霉变而不能食用。霉菌腐败是危害粮食安全储藏的关键因素[1]。黑曲霉能够导致剑麻茎腐病、枣果霉烂病、采后芒果果腐病、花生冠腐病、蓝莓及冬葡萄果实腐烂、灰枣果实病害等植物病害[2-3],由此看来,黑曲霉的防治尤为重要。研究发现,从陆地向海洋逐渐过渡的红树林生态系统蕴藏着大量的生物资源,张起畅等在东寨港红树林淤泥中检测出53个门、909个属的微生物类群[4],具有巨大的经济价值[5]。红树林具有的盐渍化、强酸性、沼泽化等特殊的生态环境,使生长于此的微生物种群及其代谢产物等都具有一定的特异性。目前已知红树林微生物中有许多菌株能够产生具有生物活性的抗性物质,其产物肽类化合物、酯类化合物、胞外多糖等已广泛应用于抗生素、免疫抑制剂、蛋白黏合抑制剂、食品添加剂的研制等方面[6-7]。郑红芸等在广西茅尾海红树林根围淤泥中发现了具有金黄色葡萄球菌抗性的3株链霉菌[8];Chi等发现了28株对枯草芽孢杆菌(Bacillus subtilis)、金黄色葡萄球菌(Staphylococcus aureus)、大肠杆菌(Escherichia coli)、白色念珠菌(Monilia albican)和新型隐球菌(Cryptococcus neoformans)有抑制作用的红树林真菌[9]。虽然红树林微生物资源丰富,发现的抗性菌种类、数量日益增多,但少见黑曲霉拮抗的微生物的报道。
目前对黑曲霉的防治主要有物理处理法和化学杀菌法,但这些方法在一定程度上会对食物本身或人体造成危害。近年来,生物抑菌剂因其低风险、低成本和高效率等特点已成为当今确保食品安全生产的主流方式,应用前景日趋广泛[10]。黑曲霉的生物防治主要集中在植物提取物或抗生素等方面,如茶树精油中的α-松油醇、萜烯-4-醇,冬青油所含的水杨酸甲酯、肉桂与山苍子复合精油均可有效抑制黑曲霉生长[11-13],黄曲霉毒素B1(AFB1)能够高效降解黑曲霉[14]。然而,目前发现的具有黑曲霉抑制作用的拮抗细菌较少,解淀粉芽孢杆菌CP 2014 无细胞提取液有明显抑制效果[15],Paenibacillus sp. 512、Brevibacterium sp. 90 等能夠有效防治黑曲霉引起的茎腐病[16],而食品领域尚未见报道。
本研究从深圳坝光红树林海泥中分离鉴定出2株对黑曲霉具有显著抑制效果的细菌,可能为新菌株,对其进行培养条件优化并确定了抑菌效果。研究结果可为充分利用红树林微生物资源、制备有效黑曲霉防治生物抑菌剂提供重要资料,具有较好的理论意义与应用价值。
1 材料与方法
1.1 材料与试剂
1.1.1 样品采集点的概况 样品的采集点位于深圳坝光红树林(114°30′E 、22°39′N),地处深圳市东部龙岗区银叶树(Heritiera littoralis Dryand)湿地公园,此地位置偏远,人为破坏与污染较少,附近海滩浅,相对封闭,水土一直保持较好[17]。该地区其他动植物、微生物资源丰富,具有较高的科研价值,是红树林生态系统研究的理想基地。试验材料于2018年6月采集,全部试验工作均在作物表观遗传调控与发育湖南省重点实验室完成。
1.1.2 主要培养基 Luria-Bertani培养基(LB)、牛肉膏蛋白胨培养基(ND)、酵母液体培养基(YEB)、营养琼脂培养基(NA)、营养肉汤培养基(NB)、马铃薯培养基(PDA)。
1.1.3 主要试剂 DNA聚合酶、dNTPs和引物,均购自湖南擎科生物技术有限公司。
1.2 仪器与设备
PCR 仪,购自Bio-Rad 公司;紫外可见分光光度计,购自北京普析通用仪器有限责任公司。
1.3 试验方法
1.3.1 海泥微生物多样性研究及抗菌活性的筛选 取少量海泥匀浆样品,在无菌试管中进行常规10倍梯度稀释,从10-5、10-6、10-7 等3个浓度梯度样品分别取200 μL稀释液,分别涂布于LB、NA、PDA、ND固体分离培养基上,37 ℃培养2~3 d后,从平板中挑取典型菌落利用三区划线法接种于对应的分离培养基上,获取纯单菌株。挑选已成熟的菌株接种在发酵培养基中,200 r/min、37 ℃恒温振荡培养2 d。选取革兰氏阴性菌2株[大肠杆菌和变形杆菌(Proteus vulgaris)],革兰氏阳性菌3 株[枯草芽孢杆菌、藤黄八叠球菌(Sarcine luted)和金黄色葡萄球菌],真菌1株(黑曲霉)作为敏感指示菌,采用琼脂扩散法[18]进行抗菌活性筛选。设置3个重复组,培养1~2 d,测量抑菌圈直径,比较抗菌活性。
1.3.2 菌株的观察与鉴定 将菌株Z-2001、R-2008接种于LB培养基上,观察并记录菌落的各项状态,芽孢染色后在光学显微镜下观察并记录其结果。采用常规方法提取基因组[19],利用16S rRNA通用引物进行PCR扩增,产物由湖南擎科生物技术有限公司测序。测序结果用MEGA 7.0构建系统进化树。
1.3.3 菌株培养条件的优化
1.3.3.1 菌株基础培养基的筛选
將活化后的菌株接种于LB液体培养基中制成种子液。选用LB、PDA、YEB、NA、NB作为基础培养基,将等量的种子液分别接种于液体基础培养基中,200 r/min、37 ℃恒温振荡培养,定期检测培养液D600 nm并比较结果。
1.3.3.2 菌株生长曲线的测定
挑取活化后的菌株,接种于基础液体培养基中,培养方法同上,无菌环境下每隔2 h取样1次,检测培养液的D600 nm连续测量24 h,绘制生长曲线。
1.3.3.3 培养基组分种类及浓度优化
以NA为基础培养基,研究不同碳源(葡萄糖、木糖、淀粉、蔗糖),氮源(酵母粉、蛋白胨、NH4NO3、KNO3),无机盐(NaCl、MgSO4·7H2O、KH2PO4+K2HPO4(1 ∶1)复合盐、ZnSO4·7H2O)的培养效果。接入3%种子液(3个重复),培养及测量方法同上,筛选出最佳培养基组合[20]。在此基础上研究碳源(0.10%、0.20%、0.25%、0.40%、1.00%、2.00%),氮源(0.3%、0.6%、0.8%、1.0%、1.2%、1.5%),无机盐(0.2%、0.3%、0.4%、0.5%)浓度对菌株培养效果的作用,获得最优组分浓度。进而采用3因素3水平L9(33)进行正交试验,以此确定培养基中各组分的最佳配比[21]。
1.3.3.4 发酵条件的优化
根据培养基优化结果,通过控制初始pH值(6、7、8),温度(28、37、46 ℃),转速(150、180、210 r/min)进行摇瓶发酵,获得最佳初始发酵条件。
1.3.4 菌株的抑菌效果 在超净工作台上,挑取活化后的单菌落接种于最佳配比液体培养基中,按照“1.3.3”节方法培养12 h,制成种子液。将得到的菌液离心,过滤后留取滤液。分别按照2、4、6、8 mL的浓度梯度,加入到PDA固体培养基中混匀,再将等量黑曲霉接种于PDA平板中间,设置3组重复,28 ℃恒温培养。每天观察并记录数据,根据公式算出抑菌率。
菌落生长抑制率=(对照生长直径-处理生长直径)/对照生长直径×100%。
2 结果与分析
2.1 抗性菌的筛选
本研究运用4 种分离培养基对采集的海泥进行微生物分离,最终获得58 株纯培养物。并用滤纸片法[22]初筛分离的58 株菌株的抗菌活性,除去试验过程中平板被污染以及其他原因无法得到测试结果的,发现有14 株菌株的发酵液至少对其中1 株敏感指示菌有不同程度的拮抗作用。在对获得的14 株具有拮抗作用的菌株进行复筛时,发现其中有2 株菌株(Z-2001、R-2008)的发酵液对黑曲霉具有较强的拮抗作用。
2.2 Z-2001、R-2008菌株的鉴定
由表1、图1可知,Z-2001、R-2008菌株二者形态相似,均为芽孢杆菌,呈革兰氏阳性。根据测序结果和进化树(图2)分析显示,Z-2001、R-2008与已知菌亲缘关系较远,可能为新种。
2.3 Z-2001、R-2008菌株培养基的单因素优化
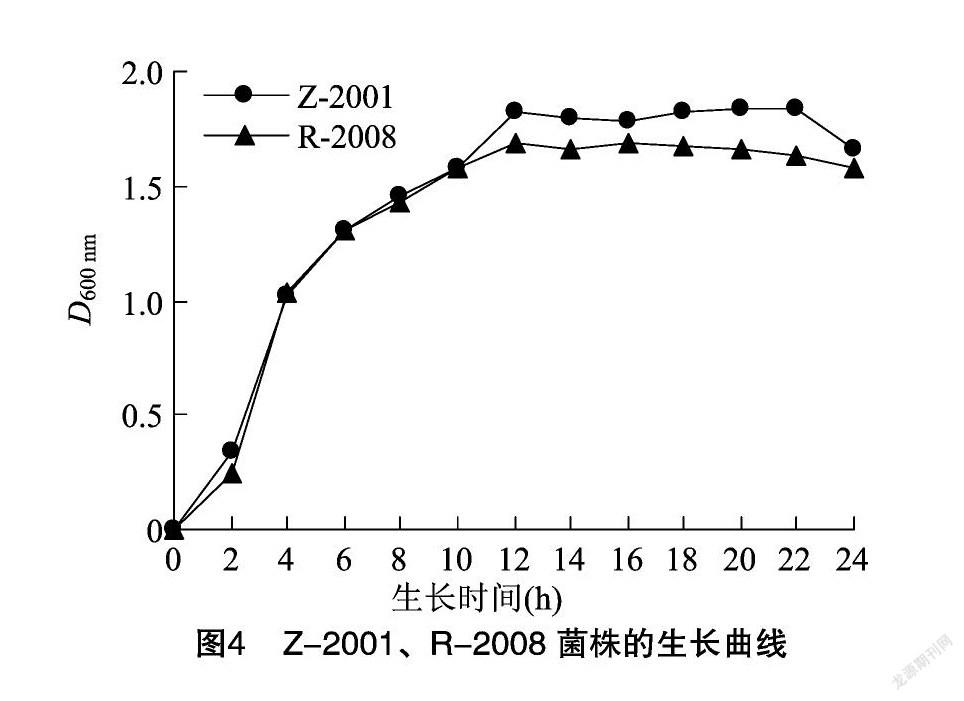
由图3可知,Z-2001、R-2008均在NA培养基上生长效果最佳,因此选用NA培养基作为后续研究的基础培养基。在连续的24 h内每2 h测定1次菌液浓度,并绘制生长曲线。由图4可知,在基础培养基上Z-2001、R-2008的生长曲线均为典型的“S”形曲线,并且都在12 h左右达到生长量的高峰。
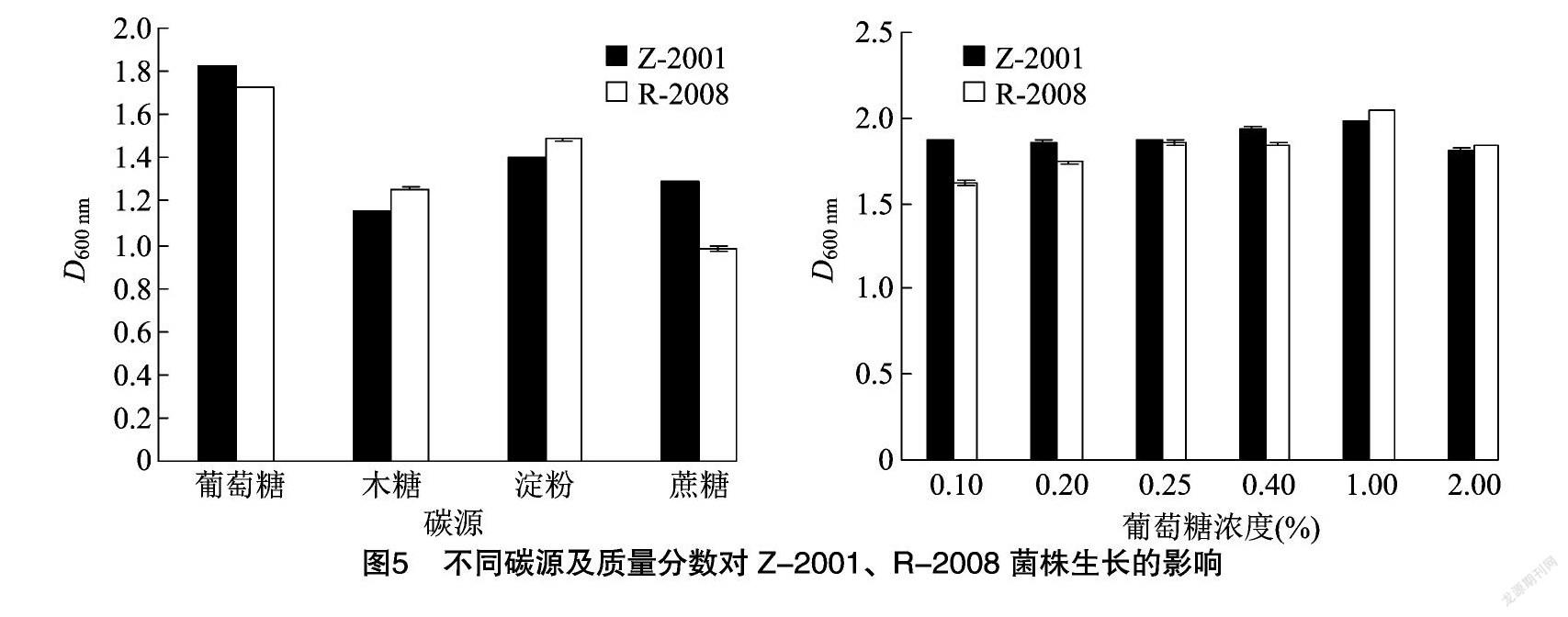
由图5可知,Z-2001、R-2008菌株对葡萄糖的利用能力最强,其次是淀粉。因此笔者所在课题组选用葡萄糖作为发酵培养基中的最佳碳源。选取5个梯度探究葡萄糖质量分数对菌株生长的影响,当葡萄糖质量分数为1.00%时,Z-2001、R-2008菌体生长量最大,D600 nm值分别为1.981、2.044。因此发酵培养基中葡萄糖的最佳质量分数为1.0%。
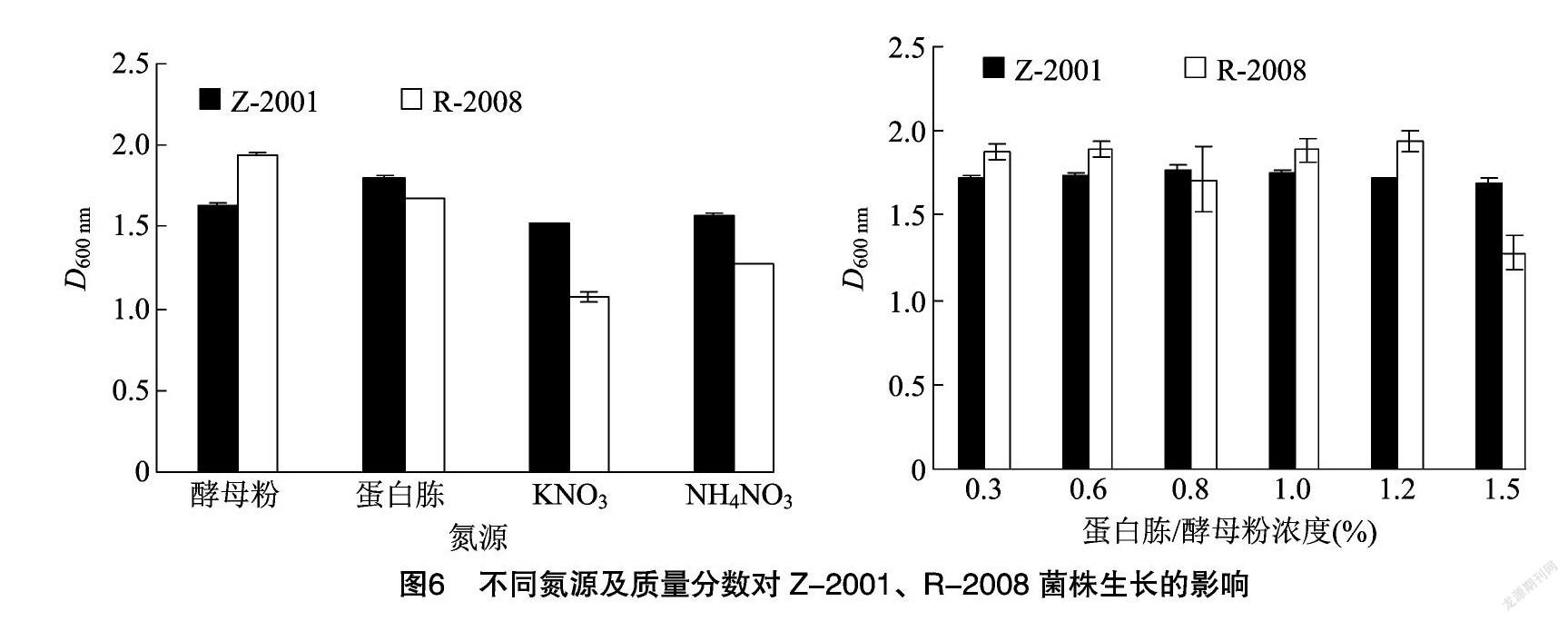
由图6可知,选取酵母粉、蛋白胨、KNO3、NH4NO3 4种氮源进行筛选,发现蛋白胨和酵母粉作为氮源时菌体的生长情况明显优于其他氮源,说明Z-2001、R-2008菌株对有机氮源的利用能力更强。原因可能在于有机氮源中还包含着碳源和少量无机盐,而无机氮源成分相对简单,可能营养不足。因而分别选用蛋白胨、酵母粉作为Z-2001、R-2008 菌株发酵培养基中的最佳氮源。选取6个梯度探究蛋白胨、酵母粉对菌株生长的影响,发现 Z-2001 菌株的D600 nm值随着蛋白胨质量分数的增加呈先上升后下降的趋势。当蛋白胨质量分数达到0.8%时D600 nm值达到峰值,随后开始缓慢下降,从而确定蛋白胨的最佳质量分数为0.8%。R-2008菌株的 D600 nm值在酵母粉质量分数为0.3%~1.0%之间变化不大,当达到1.2%时D600 nm值达最大值:1.942,之后D600 nm值开始缓慢下降。因此酵母粉的最佳质量分数为1.2%。
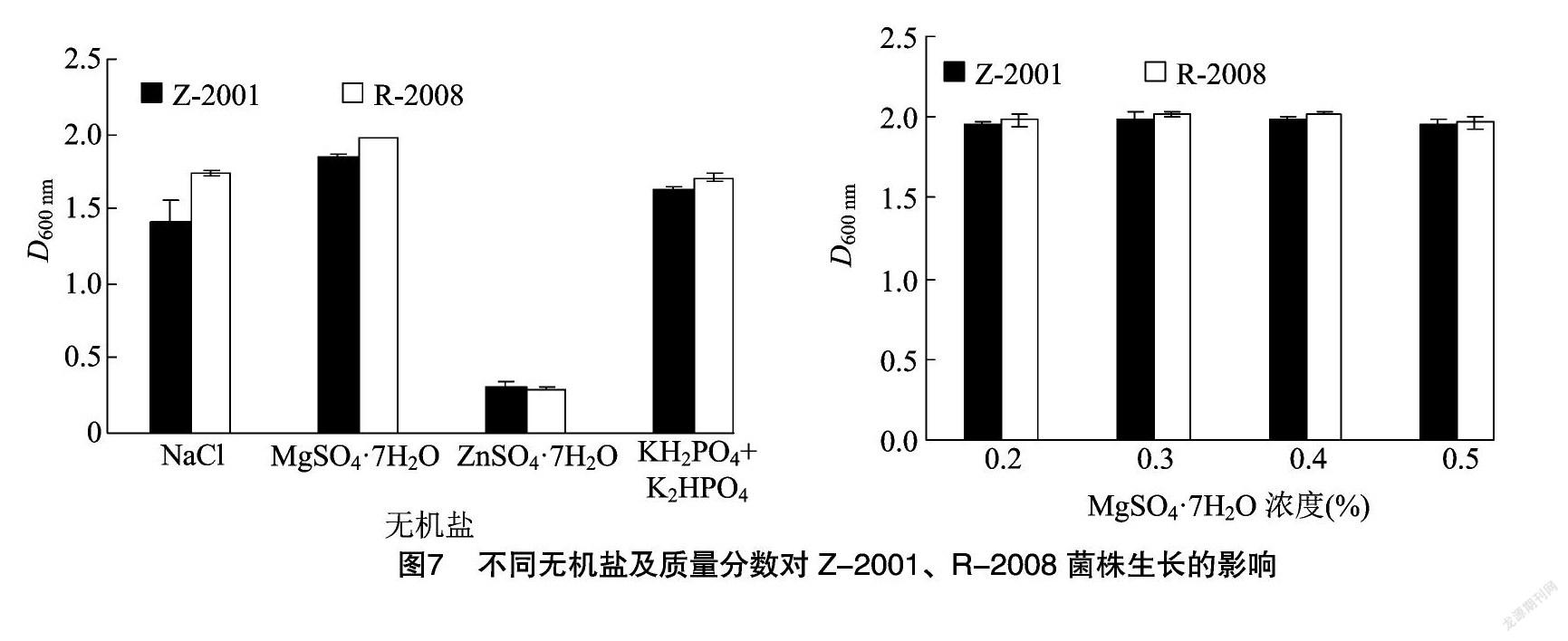
由图7可知,选取NaCl、MgSO4·7H2O、ZnSO4·7H2O以及KH2PO4+K2HPO4(1 ∶1)复合盐进行无机盐筛选。除了锌离子以外,其他离子都能够促进Z-2001、R-2008菌株的生长,相比之下镁离子作用较为明显,因此2种菌生长的最佳无机盐为MgSO4·7H2O。选取4个梯度探究最佳无机盐离子质量分数,Z-2001、R-2008菌株的D600 nm值随MgSO4·7H2O质量分数的增大呈现先上升后下降的趋势。对于Z-2001菌株来说,镁离子质量分数达到0.3%时,菌体的生长量处在较高的水平,D600 nm值也达到了最大值。而R-2008菌株则是在MgSO4·7H2O质量分数处于0.4%时达到生长最高水平。此结果与孔高飞对短小芽孢杆菌的研究结果[23]相似,因此Z-2001菌株的最优无机盐MgSO4·7H2O最佳质量分数为0.3%,R-2008为0.4%。
2.4 正交试验结果分析
由以上试验得到了单因素的结果,而这些组分组合在一起可能会相互影响,导致培养效果变差,因此需要正交试验来确定各组分配比,正交试验因素和水平见表2。由表3可知,影响Z-2001菌株菌体生长的因素主次顺序为MgSO4·7H2O>葡萄糖>蛋白胨,影响R-2008菌株菌体生长的因素主次顺序为酵母粉>葡萄糖>MgSO4·7H2O。由正交試验结果可以看出,Z-2001与R-2008最佳组合分别为A3B3C2、A3B3C2,而由均值得到的最佳组合分别为A3B2C2、A3B3C1。对这2种情况分别进行试验,结果如图8所示。因此Z-2001菌株菌体的最适培养基为葡萄糖2.0%、蛋白胨0.8%、MgSO4·7H2O 0.30%,小牛浸膏0.3%,NaCl 0.5%;R-2008 菌株菌体的最适培养基为葡萄糖2.0%,酵母粉2.4%,MgSO4·7H2O 0.2%,小牛浸膏0.3%,NaCl 0.5%。
2.5 菌株发酵条件的单因素优化
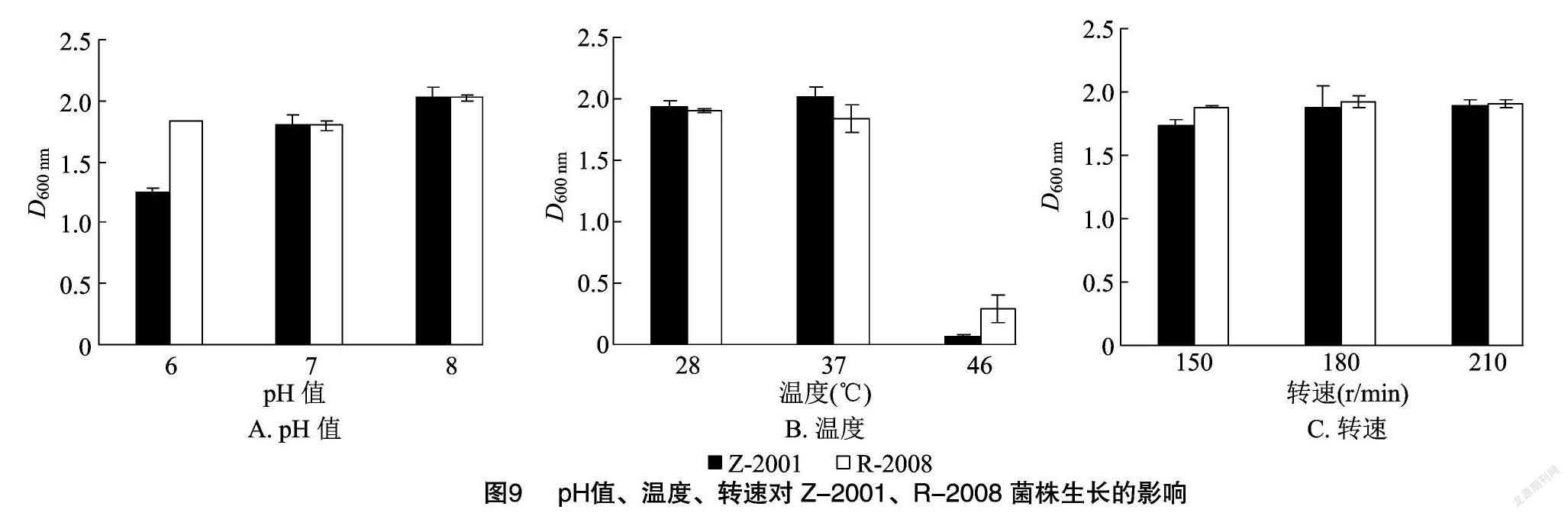
由图9-A可知,随着pH值的不断增大,Z-2001、R-2008菌株的D600 nm值也随之增大,在pH值=8时达到了最大值。说明Z-2001、R-2008 菌株在弱酸性和中性条件下可以生长,但偏碱性条件更适宜生长,因此选择pH值=8为 Z-2001、R-2008 菌株的最适pH值。
温度是影响细菌生长的一个重要因素,它能够影响菌体代谢过程中酶的反应效率。由图9-B可知,Z-2001、R-2008菌株在46 ℃培养时菌体中的酶因为温度过高而逐渐失活,菌体生长量明显变小,D600 nm值急剧下降,因此在高温条件下并不适合Z-2001、R-2008的菌株生长。而Z-2001、R-2008 菌株分别在37、28 ℃时长势良好,D600 nm值达到了最大。因此笔者所在课题组选择 37 ℃作为 Z-2001 菌株的最适温度,28 ℃作为R-2008菌株的最适温度。
在摇床的振荡培养下能够使氧气较多地进入培养液以供菌株使用,由图9-C可知,Z-2001、R-2008 菌株分别在转速为210、180 r/min时D600 nm值最大。因此笔者所在课题组选择210 r/min为Z-2001菌株的最佳转速,180 r/min为R-2008菌株的最佳转速。
2.6 黑曲霉抗性
将过滤后的2种菌液以不同含量与PDA培养基混合制成带药培养基,再将等量黑曲霉接种在平板中央,测定2种菌对黑曲霉的抑制率。由图10可知,随着发酵液量的不断增加其抑制效果逐渐显现,特别是菌液量在6 mL时抑制效果变得明显,8 mL 时基本可以抑制住黑曲霉孢子的生长。Z-2001 菌株8 mL的发酵液对黑曲霉的抑菌率可达到76.80%,R-2008菌株8 mL抑菌率可达到78.70%。
3 讨论与结论
黑曲霉是世界卫生组织认定的一类病原微生物,对人体安全、食品安全产生了极大的威胁。目前,红树林是已知生态系统中种类丰富,高利用率的湿地生态系统之一[24],蕴含着优质的微生物资源,许多微生物能产生大量有拮抗活性且效果明显的代谢物质。在现行有效的黑曲霉防治方法中,生物法具有物理、化学法无法比拟的低投资、低风险、高效率、高收益等优势。且生物法中大多利用的是植物与微生物,对环境友好且不会产生较大危害。许多植物提取物中含有重要的生物活性物质,已有研究表明肉桂提取液、丁香提取物对黑曲霉均有较强的抑制作用,肉桂抑菌圈直径可达17.49 mm,丁香抑菌率可达50%[25-26]。本研究中,芽孢杆菌 Z-2001、R-2008最大抑菌率分别可达76.80%、78.70%,相比之下优势明显,且经过试验,2种菌株大量培养所需条件简单,可以认为其具有生物抑菌剂的开发价值。
微生物的抑菌机制较为复杂,一般认为芽孢杆菌发酵产生的拮抗活性物质主要包括由核糖体组成的化合物和由非核糖体组成的化合物这两大类[27-28]。这些具有拮抗活性的物质能够使菌丝生长受阻,产生畸形或推迟分生孢子的萌发时间,导致其芽管和菌丝畸形而不能继续生长,从而实现拮抗作用[29]。本研究发现,芽孢杆菌Z-2001、R-2008代谢产物对黑曲霉拮抗效果显著,但关于2种菌株抑制黑曲霉的有效成分、作用机制尚不清楚,需要进一步研究。
Z-2001、R-2008菌株为芽孢杆菌属,培养条件简单、成本低。菌株代谢产物能有效抑制黑曲霉菌丝生长,抑菌作用显著,最高抑菌率分别达到76.80%和78.70%。研究结果可为生物抑菌剂的工业化生产提供理论依据,也可为后续深入研究奠定基础。
参考文献:
[1]冯蒙蒙.贮藏期间粮食霉变控制方法的研究[J]. 食品安全导刊,2019(15):138-139.
[2]黄雪兰,李菊馨,周海兰,等. 3种生防真菌对剑麻茎腐病病菌黑曲霉的抑制效果[J]. 农业研究与应用,2019,32(1):16-20.
[3]陈 燕,张 健,魏 佳,等. 一氧化氮熏蒸抑制干制灰枣黑曲霉病及贮藏品质保持[J]. 农业工程学报,2019,35(12):297-303.
[4]张起畅,张文飞,殷浩能,等. 宏基因组测序分析东寨港红树林淤泥和水体微生物的多样性[J]. 基因组学与应用生物学,2020,39(1):116-122.
[5]Tanner M K,Moity N,Costa M T,et al. Mangroves in the Galapagos:ecosystem services and their valuation[J]. Ecological Economics,2019,160:12-24.
[6]徐志勇,冯 昭,徐 静.红树林微生物抗菌活性成分研究进展[J]. 中国抗生素杂志,2017,42(4):241-254.
[7]Balakrishnan B,Ranishree J K,Thadikamala S,et al. Purification,characterization and production optimization of a vibriocin produced by mangrove associated Vibrio parahaemolyticus[J]. Asian Pacific Journal of Tropical Biomedicine,2014,4(4):253-261.
[8]郑红芸,吴 越,叶景静,等. 三株红树林来源链霉菌的鉴定及抗菌活性研究[J]. 中国药理学通报,2019,35(7):985-990.
[9]Chi W C,Pang K L,Chen W L,et al. Antimicrobial and iNOS inhibitory activities of the endophytic fungi isolated from the mangrove plant Acanthus ilicifolius var. xiamenensis[J]. Botanical Studies,2019,60(1):4.
[10]魏 晨,郭永鹏,计 成,等. 黄曲霉毒素生物降解技术在反刍动物生产上的应用前景[J]. 饲料工业,2018,39(19):60-64.
[11]安培培.茶树精油对采后葡萄黑曲霉和赭曲霉的抑制效果及作用机理研究[D]. 西安:陕西师范大学,2019.
[12]闫 佳,牛延菲,史正军,等. 滇产5种植物精油的抑菌性能及其成分分析[J]. 林业工程学报,2021,6(1):98-104.
[13]卢 锟,唐雅珂,龚吉军,等. 不同植物精油对大米青霉和黑曲霉的抑菌效果[J]. 食品工业科技,2020,41(3):110-113,119.
[14]Qiu T Y,Wang H M,Yang Y,et al. Exploration of biodegradation mechanism by AFB1-degrading strain Aspergillus niger FS10 and its metabolic feedback[J]. Food Control,2021,121:107609.
[15]贾瑞娟.山西老陈醋源芽孢菌抑菌机理及对腐败菌群体感应抑制的研究[D]. 太谷:山西农业大学,2019.
[16]Barbosa L O,Lima J S,Magalhes V C,et al. Compatibility and combination of selected bacterial antagonists in the biocontrol of sisal bole rot disease[J]. BioControl,2018,63(4):595-605.
[17]邓太阳,赵丹阳,邱华龙,等. 深圳坝光银叶树湿地园林木有害生物调查及防控[J]. 林业与环境科学,2019,35(1):54-60.
[18]谭才邓,朱美娟,杜淑霞,等. 抑菌试验中抑菌圈法的比较研究[J]. 食品工业,2016,37(11):122-125.
[19]王 伟,王玉琢,舒 鹏,等. 16S核糖体DNA宏基因组测序中细菌核酸提取方法的比较研究[J]. 生物技术通讯,2015,26(4):551-555.
[20]焉兆萍,宋士良,陆克文.枯草芽孢杆菌发酵培养基及发酵条件优化[J]. 国外畜牧学(猪与禽),2019,39(1):51-55.
[21]邹玉叶.正交实验设计优选教学方法的应用研究[J]. 科教导刊,2018(16):108-111.
[22]李小俊,成丽霞,吴彦彬,等. 拮抗菌抗菌谱及发酵液拮抗能力测定的新方法[J]. 生物技术,2007,17(1):55-58.
[23]孔高飛.一种短小芽孢杆菌分离鉴定及培养条件研究[D]. 杭州:浙江理工大学,2014.
[24]刘慧杰,张虎山,田 蕴,等. 红树林湿地微生物对主要污染物的净化作用[J]. 广州环境科学,2013,28(1):9-17.
[25]张 炜,吴正云,罗 力,等. 药食两用植物提取液对李果实采后常见致腐真菌抑制作用[J]. 中国酿造,2019,38(3):166-169.
[26]蒋志国,施瑞城.10种中草药提取物对常见果蔬致腐真菌的抑制作用及有效成分分析[J]. 食品科技,2006,31(4):68-71.
[27]李琪敏,周婷婷,秦春秀,等. 枯草芽孢杆菌HAB-8菌株分离筛选鉴定及抑菌机理初步研究[J]. 黑龙江农业科学,2019(2):28-36.
[28]吴海霞,田志芳. 银杏果实(白果)多糖提取工艺优化及其抑菌活性分析[J]. 江苏农业学报,2020,36(6):1551-1558.
[29]刘振宇,柳韶华,赵春青,等. 枯草芽孢杆菌(Bacillus subtilis)对桑炭疽病菌的抑制作用[J]. 蚕业科学,2005,31(4):409-412.
