蛋白质工程改造脂肪酶对映选择性的研究进展
2022-05-05黄金莎代姝函闫云君
黄金莎,代姝函,徐 莉,闫云君
华中科技大学生命科学与技术学院,分子生物物理教育部重点实验室(华中科技大学),湖北 武汉 430074
脂肪酶(lipase E.C.3.1.1.3)具有良好的化学选择性、区域选择性和对映选择性,能够催化多种天然和非天然底物的水解或合成反应[1-2],广泛应用于制药、食品、饮料、化妆品、医疗诊断、洗涤剂和纸张等行业[3-4]。脂肪酶反应条件温和,具有天然耐有机溶剂的特性,在非水相反应中表现出优异的催化能力,可用于外消旋化合物的动力学拆分[5],这些反应中产生的高光学纯度化合物是制药工业的重要组成部分。
脂肪酶的三维结构研究表明,大多数脂肪酶具有共同的α/β水解酶折叠特征、Ser-His-Asp/Glu组成的催化三联体和保守序列G-X1-S-X2-G。脂肪酶的活性位点位于蛋白质结构中β折叠顶部的口袋内部[6]。Pleiss等根据其结合位点的几何形状将脂肪酶分为3类(图1):(1)位于蛋白表面附近的具有疏水性的裂缝状(crevice-like)结合口袋的脂肪酶,例如米黑根毛霉脂肪酶(Rhizomucor mieheilipase,RML),华根霉脂肪酶(Rhizopus chinensislipase,RCL),米根霉脂肪酶(R.oryzaelipase,ROL),及疏棉状嗜热丝孢菌脂肪酶(Thermomyces lanuginosuslipase,TLL)等;(2)具有漏斗状(funnel-like)结合口袋的脂肪酶,如南极假丝酵母脂肪酶B(Candida antarcticalipase B,CALB),洋葱伯克霍尔德菌脂肪酶(Burkholderia cepacialipase,BCL),洋葱假单胞菌脂肪酶(Pseudomonas cepacialipase,PCL),及哺乳动物胰脂肪酶等;(3)具有隧道状(tunnel-like)结合口袋的脂肪酶,如皱褶假丝酵母脂肪酶(Candida rugosalipase,CRL)。大多数的脂肪酶具有“盖子”结构,通常由1个或多个α螺旋组成,通过柔性结构与酶主体结构相连。它在油水界面产生构象变化,使底物能够进入活性位点;相反,当界面消失时盖子闭合,活性位点入口被阻断,酶失去或显示较低酶活性。

图1 脂肪酶底物结合口袋的形状:(a)裂缝状,(b)漏斗状,(c)隧道状Fig.1 Shape of three types of binding site of lipases:(a)crevice-like,(b)funnel-like,(c)tunnel-like
脂肪酶催化机制复杂,其活性位点与底物间相互作用决定了其对特定底物具有高度特异性[7-8]。因此,筛选、挖掘具有特异的对映选择性的脂肪酶资源,开发拆分特定手性分子的新策略极具挑战性。当前改良现有已知的脂肪酶对目标分子的对映选择性的策略主要概括为2方面[9-11]:(1)从自然界中寻找具有所需选择性的特定的脂肪酶资源,但其任务艰巨成功率不高;(2)蛋白质工程改造已有的脂肪酶或者从头设计新酶。其中定向进化(directed evolution)、半理性设计(semirational design)、理性设计(rational design)是目前改善脂肪酶对映选择性的最常用策略[12]。本文通过分析脂肪酶中影响特定底物对映选择性的相关区域的关键氨基酸,梳理如何通过蛋白质工程手段改进其对映选择性。
1 对映选择性的量化参数
在脂肪酶动力学拆分外消旋化合物中,有两个重要参数,对映体比率(enantio selectivity,E)和对映体过剩值(enantiomeric excess,ee)。
对映体过剩值ee表示其光学产率,即2个对映体之间的关系。它可以根据底物(eeS)或产物(eeP)定义,通常以百分比表示(公式1,2):
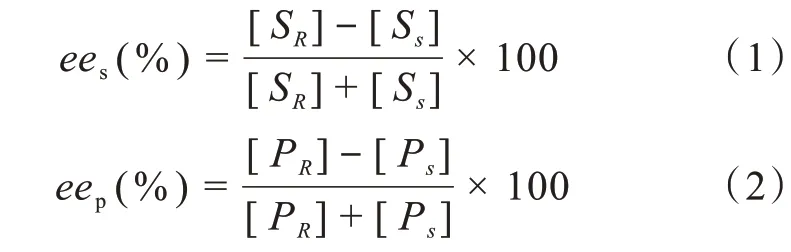
其中S代表手性底物,P代表手性产物,下标R和S分别代表R-对映体和S-对映体。外消旋体的ee值为零,纯的对映体化合物的ee值为100%。
对映体选择率E表示催化剂的拆分效率,是一个综合评价值,大小和对映体选择性的高低成正比,它是由两种底物对映异构体的特异性常数之比给出(公式3):
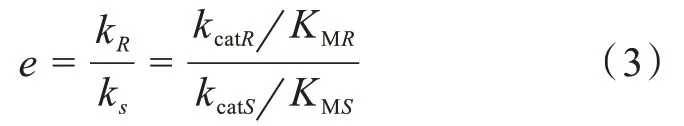
其中kR和kS是特异性常数,kcatR和kcatS是催化常数,KMR和KMS是饱和常数;下标R和S分别表示底物的R-和S-对映体。本文采用ER和ES表示脂肪酶的对映体偏好性,其中下标R和S分别表示底物的R-和S-对映体。
2 改造脂肪酶对映选择性的蛋白质工程策略
蛋白质工程策略主要包括定向进化、半理性设计和理性设计[2,12-14]。随机进化策略和借助生物信息学方法、有针对性地改造蛋白质的半理性设计及理性设计是迄今为止改善脂肪酶对映选择性的最常用定向进化策略。本文主要总结已报道的改善脂肪酶的对映选择性的蛋白质工程方法,如表1和表2所示。
2.1 定向进化(directed evolution)
定向进化通过诱变、筛选和选择的反复循环来模拟体外的自然进化过程,积累有益的突变[1,15],且具有不需要脂肪酶的先验结构和功能背景知识等优点[16]。诱变中使用的主要方法包括以易错PCR(error-prone PCR,epPCR)[17]和饱和突变(saturation mutagenesis,SM)[12]为代表的非重组技术及以DNA shuffling[18]为代表的DNA重组技术。Reetz等[19]使用epPCR生成铜绿假单胞菌脂肪酶(Pseudomonas aeruginosalipase,PAL)突变体,用于水解外消旋酯(R,S)-对硝基苯基-2-甲基癸酸酯,其中一个突变体的S-对映异构体选择性(ES=10)是野生型PAL(ES=1.1)的10倍。使用定向进化获得具有改进对映选择性的突变体的瓶颈在于突变体文库的规模非常大,不利于筛选,可靠且灵敏的高通量筛选方法是成功的必要条件[20]。
2.2 理性设计(rational design)
理性设计是基于对酶的结构和功能的广泛了解[1,14],通过分析酶的结构,对多序列进行比对,并使用计算方法进行分子动力学模拟,可以更精准的评估蛋白质序列中最有可能的目标位点,因此理性设计策略生成的突变体文库比定向进化方法生成的文库小得多[18,21]。计算过程中识别的蛋白质序列中可能的目标位点,通常被称为热点(hotspots)。常用的定点突变的方法是重叠延伸PCR或全质粒单轮PCR[22]。我们团队基于针对酶活性中心的“Inside-out”设计策略,使用量子化学方法计算出稳定过渡态的理论酶,通过RosettaMatch筛选能够容纳该理论酶构象的支架蛋白库,然后RosettaDesign设计优化匹配后构型,最终成功设计了特异性催化对硝基苯酚乙酸酯的酯酶[23]。Moharana等[24]通过分析ω-3脂肪酸的结构信息,计算酶-底物亲和力,缩小了喜热噬油芽胞杆菌脂肪酶(Geobacillus thermoleovoranslipase,GTL)活性位点的脂肪酸结合通道,限制水解过程中对较大的ω-3脂肪酸的识别和结合,获得的突变体具有较强的ω-3脂肪酸富集能力。
2.3 半理性设计(semi-rational design)
半理性设计策略是通过与理性工具相结合的定向进化实现的,克服了每一种孤立方法的局限性。半理性设计方法依靠序列和结构信息,结合计算预测算法生成突变体,突变文库的规模,减小了筛选工作量,提高酶分子改造效率。半理性设计有一个经典的方法-位点饱和突变。20世纪90年代,Reetz教授开发了组合活性中心饱和突变策略(combinatorial active-site saturation test,CAST)及迭代饱和突变技术(iterative saturation mutagenesis,ISM)[25-26]。Sun等[27-28]在CAST基础上开发了三密码子饱和突变技术(triple-code saturation mutagenesis,TCSM),进一步降低了筛选工作量。2019年Reetz教授与吴起团队合作开发了Focused Rational Iterative Site-specific Mutagenesis(FRISM)策略[29],获得了4个在外消旋底物的酯交换反应中具有高度立体互补的南极假丝酵母脂肪酶B突变体。通过迭代位点特异性诱变积累单个突变形成超小突变体文库(<100),得到的突变体对所有4种可能的立体异构体均实现了大于90%的选择性[30]。
3 改善脂肪酶对映选择性的关键突变位点
蛋白质工程改造脂肪酶对映选择性,旨在确定提高对映选择性突变的关键位置,以获得的最佳对映选择性结果。由于酶的活性中心对脂肪酶的催化活性起着至关重要的作用,因此包含催化位点和底物结合位点的催化裂缝(catalytic cleft)成为了脂肪酶改造的热点区域[14,31]。图2显示了洋葱假单胞菌脂肪酶PCL催化裂缝结构中底物结合 位 点 的 几 个 区 域[32]:(1)酰 基 结 合 口 袋(hydrophobic crevice)为脂肪酸酰基链结合区域,是对三酰基甘油的脂肪酸部分具有高亲和力的非极性区域,而在合成反应中,羧酸部分朝着它定向;(2)疏水腔(hydrophobic dent),大且浅的疏水区域,能够结合手性醇的大的疏水取代基;(3)亲水腔(hydrophilic trench),靠近催化Ser“入口区域”,可作为手性醇部分中等大小取代基的结合位点;(4)氧阴离子洞(oxyanion hole),稳定过渡状态下形成的四面体中间体。Bornscheuer等[33]提出将催化裂缝分成羧酸结合部分和酰基结合部分两个区域,其中,羧酸结合部分是较大的疏水口袋;而酰基结合部分是较小的醇结合部分。基于上述分析,定位催化裂缝中关键区域的突变位点,取决于手性底物是羧酸还是醇。即当底物是外消旋羧酸时,最好的突变位于酰基结合口袋、疏水腔和氧阴离子洞。另一方面,当底物是外消旋醇时,突变集中在疏水腔、亲水腔以及氧阴离子洞。

图2 洋葱假单胞菌脂肪酶催化裂缝结构[31]Fig.2 Catalytic cleft of Pseudomonas cepacia lipase
3.1 脂肪酶对外消旋羧酸立体选择性改造
在涉及手性羧酸部分的拆分反应中,反应的方向(合成或水解)影响突变位点的选择。在合成反应中,羧酸是首先进入催化裂隙的底物。已使用计算机模拟了手性羧酸的对映异构体与催化裂隙之间的相互作用,确定控制对映选择性的关键氨基酸位置(表1)。

表1 蛋白质工程改善脂肪酶对外消旋羧酸对映选择性的部分研究Tab.1 Improving enantioselectivity of lipase towards racemic carboxylic acid by protein engineering
这些研究中使用最多的脂肪酶是南极假丝酵母脂肪酶B。Gu等[34]使用分子对接研究了不同底物的酰基部分的变化(乙酰基或丁酰基)对底物与催化裂缝中氨基酸残基间相互作用的影响。被鉴定的关键残基是位于酰基结合口袋和疏水腔中的W104,D134,Q157和I189,以及位于CALB的α-螺旋10上的L278,A282和I285。后来,研究人员[35-37]使用计算机模拟方法识别了不同底物的关键残基,不仅有先前验证的W104,D134,Q157和I189,还有L144,V149,V190和V154[34]。CALB的这些研究结果表明,即使关键残基大多数集中于酰基结合口袋中,催化裂缝裂隙的其他区域(如疏水腔)中的突变也有助于改善CALB的对映选择性。
南极假丝酵母脂肪酶A在合成反应中的使用少于CALB。但是,它具有的一些非同寻常的特性(如热稳定的,能在90℃以上的温度下起作用),已经激发了越来越多研究者的兴趣,以期提高其对映选择性。对三酰基甘油酯的水解具有特定的sn-2特质,并且可以催化侧链基团较大的底物,例如叔醇[38]。Engström等[39]组合突变酰基结合口袋中的F149Y,I150N和F233G,得到的突变体对7种不同酯的E值为45-276,与野生型的2-20相比,有很大的提高。且对于大多数底物而言,活性也增加了30倍。更有趣的是,Frushicheva等[40]将量子力学/分子力学(quantum mechanics/molecular mechanics,QM/MM)方法用于硝基苯基2-甲基庚酸酯的水解反应。与野生型相比,得到的组合突变体(F149Y/I150N/F233G)实现了R-向S-对映异构体的逆转。
洋葱伯克霍尔德菌脂肪酶BCL常作为模型用于改造对映选择性的突变研究。野生型脂肪酶对于(R,S)-1,4-二氢吡啶的水解是R-对映选择性的,突变体V266L,L287I和F221L(疏水腔中)却表现出相反的对映选择性(S-对映选择性)[41]。Koga等[5]确定了4个关键氨基酸:氧阴离子洞中的L17、酰基结合口袋中的F119和L167、疏水腔中的L266;获得的2个突变体(突变体1:L17F/F119L/L167G/L266V/T251A,突 变 体2:L17F/F119L/L167A/L266V/D21N))具有反向对映体选择和相对高的对映选择性。其中L167被较小的甘氨酸Gly和丙氨酸Ala取代扩大了催化裂缝的体积,促进了与对映体的结合。
Reetz等[19]使用定向进化改进PAL在(R,S)-对硝基苯基-2-甲基癸酸酯水解中的对映选择性,获得了具有S-对映选择性的PAL突变体(ES=11)。Prasad等[42]在PAL的几个特定氨基酸(氧阴离子洞中的M16和L17,酰基结合口袋中的L159、L162、L231和V232)进行了迭代饱和诱变,试图提高其对(R,S)-2-甲基癸酸酯的对映选择性,获得的最佳组合突变体(M16A/L17F/L159V/L162V)ES值(S-对映选择性)提高到436。
2011年,Ni等[43]系统地研究了枯草芽孢杆菌脂肪酶(Bacillus subtilislipase A,LipA),使用QM/MM及分子对接策略,解析了其对(R,S)-酮洛芬乙烯基酯的立体选择性。实验结果与理论预测相吻合:M 78(氧阴离子洞中)和V136(疏水腔中)可通过稳定R-酮洛芬乙烯基酯末端苯基影响对映选择性。
皱褶假丝酵母脂肪酶CRL能特异性水解S-酮洛芬甲酯,Manetti等[44]使用分子对接技术分析了影响(R,S)-酮洛芬甲酯水解的关键氨基酸,试图通过组合突变F344V/F345V(疏水腔)提高R-对映异构体的反应速率。但突变影响了苯丙氨酸Phe侧链与酮洛芬的芳香族的相互作用,降低了酶与底物结合的自由能,ES从27降低到5。
综上所述,影响羧酸手性识别的主要位点在疏水腔、氧阴离子洞和酰基结合口袋中。一般而言,用较小的氨基酸替换较大的氨基酸残基可能会增加脂肪酶的对应选择性,或逆转脂肪酶的对映选择性。
3.2 脂肪酶对外消旋醇的立体选择性改造研究
正如上述外消旋羧酸研究中所观察到的,某些关键位置会影响脂肪酶对手性醇的对映选择性,这些位点多数位于疏水腔和亲水腔中。表2总结了近年来蛋白质工程改善脂肪酶对外消旋醇对映选择性的部分研究。
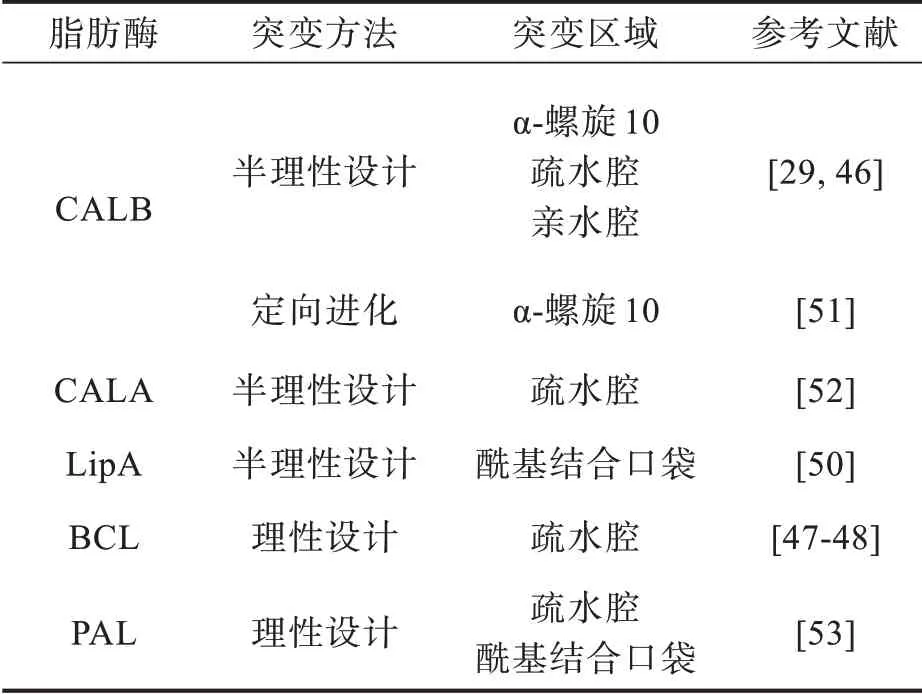
表2 蛋白质工程改善脂肪酶对外消旋醇对映选择性的部分研究Tab.2 Improving enantioselectivity of lipase to racemic alcohols by protein engineering
Liu等[46]使用FlexX对接(网址为https://www.biosolveit.de/products/#FlexX)和动力学模拟技术研究了CALB与外消旋仲醇的丙烯酸的相互作用,以L278、W104和S47为靶位,构建了104位和278位双位点饱和突变文库。然后筛选用于水解丙烯酸羟丙基氨基甲酸酯的文库。最佳突变体L278A、L278V、L278A/W104F和L278A/W104F/S47A的水解转化率和酯交换率均高于30%。该研究结果证实了通过改变催化裂缝的体积调节S-对映异构体与S47,W104和α-螺旋10中L278的相互作用,改变其对应选择性,实现外消旋醇和酯的高效高活性转化。
Ema等[47]通过取代位于疏水腔中的I287,改变了BCL对(R,S)-1-苯乙醇的对映选择性。与野生型BCL的ER值相比,用苯丙氨酸Phe(I287F)替代减少了催化裂隙的体积,使ER值翻倍;而用丙氨酸Ala(I287A)替代增加了催化cleft的体积,ER值降低了18倍。2010年,Ema等[48]在突变体I287F的疏水腔处插入了第二个突变体I290A,得到的组合突变体I287F/I290A对应对映选择性显著提高,ER>200(野生型的ER=5)。表明减小催化裂缝的体积能够提高对仲醇的对映选择性。2012年,Ema等[49]又检测了其他12种羟基两侧具有较大取代基的仲醇底物的动力学拆分,对于所有底物,野生型BCL的E值范围为1-143,突变体I287F/I290A的ER值均超过200。
Aileen等[50]以(R,S)-1-(2-萘基)乙酸乙酯的水解作为模型反应,使用QM/MM方法识别枯草芽孢杆菌LipA中突变的目标位点。亲水腔中靠近催化S77的残基H76被确定为影响LipA对映选择性的关键氨基酸残基。与野生型LipA的ER=156相比,突变H76A具有相反的对映选择性(ES=9),这可能与76位氨基酸残基的电荷、极性和取向对反应活化能的影响有关。
4 结论与展望
通过分析脂肪酶催化裂缝的特性对动力学拆分中对映选择性的影响,提出了通过蛋白质工程方法在关键区域引入突变以提高脂肪酶的对映选择性。在外消旋羧酸的拆分中,影响羧酸手性识别的主要位点在疏水腔、氧阴离子洞和酰基结合口袋中;在外消旋醇的拆分中,影响脂肪酶对映选择性的关键突变位点主要位于亲水腔、疏水腔和氧阴离子洞。但目前酶结构和功能解析不够全面,通过对酶催化裂缝结构进行改造,定向设计具有特定活性的生物催化剂极具挑战。
随着计算机技术的发展、运算能力的提升,酶的三维结构的解析、催化机理的挖掘,结构和功能之间的关系越来越清晰,计算机辅助设计及人工智能在蛋白质设计与改造方面受到前所未有的关注,使得基于特定目的“量身定做”的酶分子成为可能。CASCO[54]是利用RosettaDesign和HTMIMD重新设计酶的催化选择性的新策略,RosettaDesign生成预定义的底物-结合位点或与过渡态模型形状互补的位点的蛋白质序列,使用高通量动力学模拟HTMI-MD预测相对活性和对映选择性取代大多数的实验筛选,提高蛋白质工程工作的可靠性和效率。FuncLib[55]是专门为结合位点添加多点突变而设计的,在进行进化分析和能量计算后,利用预测的稳定自由能(DDG)对单点突变进行组合和排序。FuncLib确保不会引入有害的突变,并且它可以解释合并多个突变所产生的潜在上位性影响。多底物酶特异性工程是促进生物催化在工业上应用的迫切需要,多化学态分析(multichemical state analysis,MCSA)策略[56]利用大的结构集合和序列空间针对多个目标底物特异性重新设计酶,为广泛特异性生物催化剂和特异性多底物酶的设计打开了大门。因此,基于FuncLib、RosettaDesign、CASCO、MCSA等方法重塑底物结合口袋,有望在脂肪酶立体选择性、区域选择性和化学选择性以及催化活力的定向进化中设计更高质量的突变体库,使新型催化剂精准设计变为现实。
