石羊河中下游不同生境柽柳群落物种多样性及土壤水分、盐分关系
2022-05-04韩福贵满多清郑庆钟张裕年付贵全
韩福贵, 满多清, 郑庆钟, 肖 斌, 张裕年, 付贵全, 杜 娟
(1.甘肃省治沙研究所甘肃河西走廊森林生态系统国家定位观测研究站,肃 武威 733000; 2.甘肃省治沙研究所荒漠化与风沙灾害防治国家重点实验室培育基地, 兰州 730070)
湿地与人类的生存、繁衍、发展息息相关,是自然界最富生物多样性的生态景观和人类最重要的生存环境之一,它不仅为人类提供了许多动植物资源,而且在控制土壤侵蚀、促淤造陆、调节径流、调节气候、降解环境污染等方面起着极其重要的作用[1]。然而湿地随着社会经济发展、人口的不断增加和城市化进程的加速,导致地下水持续下降,越来越多的湿地严重退化甚至消失[2],导致湿地生态系统功能极不稳定,严重威胁当地区域生态安全[3-4]。石羊河是中国西北干旱区典型的内陆河流域,总面积为4.16万km2,红崖山水库和青土湖湿地分别位于石羊河流域的中游和下游。为了区域补偿地下水和改善区域水环境,促进河流系统及区域生态系统恢复,由政府主导,从2010年,红崖山水库首次向青土湖人工输入生态用水起,逐渐扩大了湿地面积,到如今,青土湖已变成水草肥美的天然绿洲[5]。多年来,在地方政府的大力支持下,通过逐年增加退耕还林(草)、退地还湿区域,进一步加强石羊河流域中下游的湿地保护、防沙治沙力度,探索和运用生态补偿机制,为认识区域生态用水过程等方面发挥了至关重要的作用[6-8]。
柽柳(Tamarixchinensis)属柽柳科(Tamaricaceae)柽柳属(Tamarix),产于中国各地,为乔木或灌木,广泛分布于我国北方地区,其中内蒙和西北的沙漠地区呈片状分布,常生于河岸,海滨沙地、滩涂、潮湿盐碱地和沙荒地[9],固沙能力强,是防风固沙先锋树种。因受地形、地貌和气候等条件的影响,在河西地区柽柳适宜大量生存,在含盐量较高的河岸边、青土湖湿地外围等区域,有天然和人工栽植的柽柳灌丛群落,成片的连续分布,具有地带性分布特征。随着石羊河流域综合治理的不断推进,河岸柽柳植被得以恢复,柽柳逐渐成为石羊河流域中下游湿地重要的植被建群种之一。
前人主要对石羊河中下游柽柳灌丛及人工输水对生态环境的影响进行深入研究,屈艳萍等[10]对武威石羊河流域人工长穗柽柳树干液流量变化规律进行研究;于军等[11]对长穗柽柳种子形态特征与萌发特性进行研究;徐高兴等[12]分析民勤绿洲边缘地下水埋深对柽柳灌丛生长及物种多样性的影响。目前,对石羊河中下游河岸带不同生境柽柳群落物种多样性及其土壤水分的关系研究较少。本项目通过对石羊河流域中下游不同立地类型柽柳群落物种多样性特征的分析,掌握柽柳群落物种多样性对土壤水分变化的响应机制,为石羊河流域中下游柽柳群落保护与利用提供科学依据,同时为盐碱湿地开展柽柳人工造林,恢复湿地植被提供科学依据,为石羊河中下游不同湿地生态系统功能恢复及群落结构稳定性提供基础数据和参考依据。
1 材料与方法
1.1 研究区概况
试验地选择在民勤县西渠镇北部青土湖湿地与扎子沟河岸边带,海拔1 297.5~1 389.1 m,地理坐标为38°11′35″—39°09′18.27″N,102°46′10″—103°36′10″E,东、西、北3个方向均为原青土湖湿地(20世纪50年代)的湖面区域[13]。年均气温7.4℃,极端最高气温38.1℃,极端最低气温 - 28.8℃,年平均日较差15.9℃,最剧烈可达28.3℃,≥10℃积温为3 248.8℃[14]。年日照时数2 832.1 h;多年平均降水量113.2 mm,主要集中在7—9月,年平均蒸发量2 604.3 mm;是降水量的23.6倍;全年风沙日数达83 d,多集中在2—5月份,年平均风速2.3 m/s,最大风速23.0 m/s[15]。区域以旱生灌木、半灌木和一年生草本为主,其中灌木主要有柽柳(Tamarixchinensis)、白刺(Nitrariatangutorum)、黑果枸杞(Lyciumruthenicum)、小果白刺(Nitrariasibirica)、枸杞(Lyciumchinois)等,草本主要有白茎盐生草(Halogetonglomeratus)、雾冰藜(Bassiadasyphylla)、虎尾草(ChlorisvirgataSw)、赖草(Leymussecalinus)、针茅(Stipacapillata)、独行菜(L.ageratumapetalum)、鹅绒藤(Cynanchumchinense)、骆驼蓬、Peganumnigellastrum、西伯利亚滨藜(Atriplexsibirica)、芦苇(Phragmitesaustralis)等。根据研究区的土壤类型和地貌特点,将柽柳种群分布划分为退耕湿地(退耕还湿地)、丘间低地、固定盐碱沙地、盐碱地等4种立地类型见表1。
1.2 研究方法
1.2.1 样地调查方法 在石羊河流域下游民勤青土湖选择退耕湿地、固定盐碱沙地、丘间湿地各一个样地,在扎子沟选择盐碱湿地一个样地,共4个样地(图1)。在每个样地设置100 m ×100 m的大样方,采用5点法在每个大样方设置5个10 m ×10 m的灌木小样方,在设置的大样方内随机抽取1 m × 1 m的草本层样方3个,每个样地3个重复。植被调查时间为2019年6-9月,用荒漠生态调查方法对样方内植物种类、数量、高度、冠幅等指标进行调查,同时用GPS定位记录每个样地的经纬度、海拔。

表1 不同地理位置柽柳灌丛优势种群及土壤类型

图1 石羊河中下游不同湿地柽柳灌丛群落调查样地分布
1.2.2 物种多样性指数分析方法 选取Margalef丰富度指数(Dma)、Simpson物种多样性指数(D)、Shannon-Wiener多样性指数(H)衡量植物群落物种多样性特征;Pielou均匀度指数(Jsw)、群落优势度指数(D1)和Alatalo均匀度指数(Ea)来衡量植物群落物种的分布均匀程度[16-17]。计算公式如下:
Margalef丰富度(richness indices):
Dma=(S-1)/lnN
(1)
Simpson多样性指数:
(2)
Shannon-Wiener多样性指数:
(3)
Pielou均匀度指数(Jsw):
Jsw=(∑pilnpi)/lnS
(4)
Alatalao均匀度指数:
(5)
群落优势度指数:
(6)
式中:S为样方中物种总数;pi=Ni/N,第i个物种的相对多度;Ni为样方中第i个物种的个体数;N为样方中记录的个体总数。
1.2.3 土壤水分的测定方法 采样时间2019年7月,采用烘干法测定土壤含水率,具体方法为:以0—10 cm,10—20 cm,20—30 cm,30—40 cm,40—60 cm,60—80 cm 6个土层深度分别用土钻进行取样,取样后放入土壤盒称重,每个深度重复取样2次,然后在105℃的恒温箱内烘干,取平均值为各土层绝对含水率。
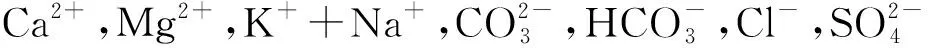
1.2.5 数据统计分析 采用SPSS数据处理软件进行统计分析,为检验不同样地各统计指标的差异性,采用邓肯检验法对均值差异显著性进行检验,用Excel进行绘图,线性回归均值分析。
2 结果与分析
2.1 不同湿地类型柽柳灌丛群落物种组成和结构特征
2.1.1 研究区植被概况 调查结果表明,在研究样地中出现物种共计9科20属22种,其中藜科5属5种,蒺藜科2属3种,禾本科6属6种,茄科1属2种,柽柳科2属2种,其余毛茛科、十字花科、豆科、萝藦科均为1属 1种。在不同生境类型的样地中,丘间低地物种组成数目最多、其次为盐碱沙地、退耕湿地、固定盐碱沙地较少。研究区内植被群落结构比较简单,存在单科单属单种现象,灌木层主要有柽柳科、茄科和蒺藜科等植物,草本层主要是能适应干旱盐碱化的藜科和禾本科等植物。从生活型上看,灌木层物种(8种)、多年生草本与一年生草本物种数相同,均为7种(表2)。分别占总物种数的36.36%,31.82%,31.82%。

表2 植物群落物种组成
2.1.2 不同湿地类型柽柳种群主要生物学特征 不同生境柽柳群落植被株数、株高、新梢长、冠幅、密度和盖度等种群生物学特征差异均显著(p<0.05)(表3)。样方内(10 m×10 m)柽柳群落株数从大到小依次排列为固定盐碱沙地>盐碱沙地>退耕湿地>丘间低地,各湿地类型退耕地和盐碱地差异不显著(p>0.05),其余差异显著(p<0.05) ;株高从大到小依次排列为丘间低地>退耕湿地>固定盐碱沙地>盐碱沙地,各湿地类型丘间低地和固定盐碱沙地差异不显著(p>0.05),其余差异均显著(p<0.05);冠幅从大到小依次排列为丘间低地、固定盐碱沙地、退耕湿地和盐碱沙地,各湿地类型盐碱地和丘间低地差异不显著(p>0.05),其余差异显著(p<0.05);密度从大到小依次排列为退耕湿地和盐碱沙地最大、固定盐碱沙地次之、丘间低地最小,且各湿地类型差异不显著(p>0.05) ;新梢长从大到小依次排列为丘间低地、固定盐碱沙地、盐碱沙地、退耕湿地,各湿地类型退耕湿地和盐碱地差异不显著(p>0.05),其余差异显著(p<0.05) ;盖度从大到小依次排列为固定盐碱沙地最高、丘间低地和盐碱沙地次之、退耕湿地最小,其中盐碱沙地和丘间低地盖度相同差异不显著(p>0.05),其余差异显著(p<0.05)。
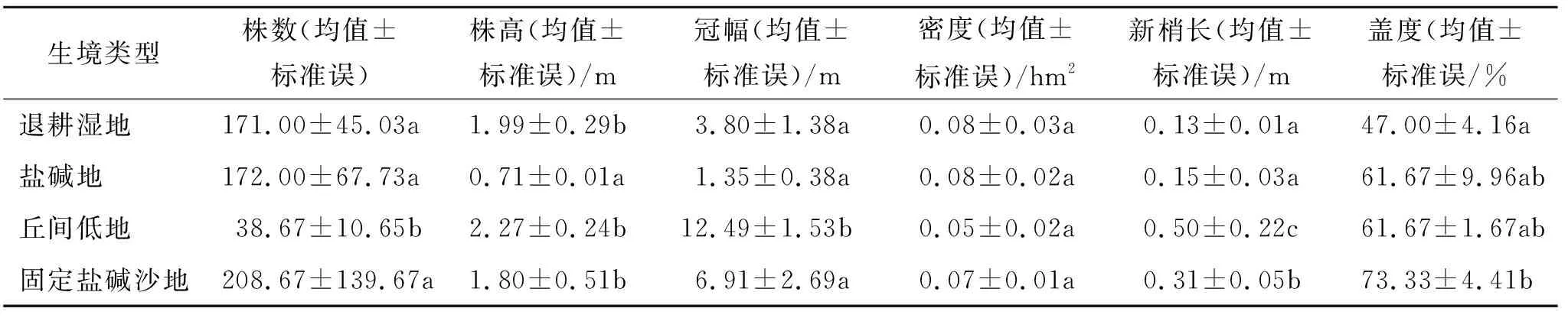
表3 不同湿地类型柽柳灌丛种群生物学特征
2.1.3 不同湿地类型柽柳群落的物种多样性特征 由调查结果得知(见图2—3),丘间低地物种Margalef丰富度指数(Dma)、Simpson物种多样性指数(D)、Shannon-Wiener多样性指数(H)、Alatulo群落均匀度指数(Ea)显著高于盐碱地、退耕湿地和固定盐碱沙地,其作用比较显著(p<0.05),当Alatulo群落均匀度指数越高时,群落优势度指数相对越低,物种比较丰富,群落结构相对稳定; Margalef丰富度指数和Shanon-wiener多样性指数,当群落优势度指数较高,群落Pielou均匀度指数相应的呈降低趋势,群落的建群种逐步趋向单一的物种发展。但是群落优势度指数丘间低地明显低于退耕湿地、盐碱地和固定盐碱沙地,从Alatulo群落均匀度指数来看,丘间低地>盐碱地>固定盐碱沙地>退耕湿地,且差异均具有统计学意义(p<0.05),但是退耕湿地和固定盐碱沙地植物种类较少,结构简单,物种丰富度也较低,群落垂直分层不明显,群落较为不稳定。从Pielou均匀度指数看,丘间低地和固定盐碱沙地差异显著(p<0.05),盐碱地和退耕湿地物种差异不显著(p>0.05)。
2.2 不同湿地类型柽柳群落土壤含水率和全盐量变化规律
从不同生境柽柳群落土壤平均水分含量来看(图4),固定盐碱沙地水分含量最高,为13.23%,退耕湿地次之,为11.58%,盐碱地和丘间低地最低分别,为9.33%和9.15%。从不同土层深度来看,变化差异明显,盐减地以表层深度0—10 cm水分含量高,为21.21%, 40—80 cm最低,为3.08%;固定盐碱沙地次之,以土层深度30—40 cm水分含量最高,为17.22%, 0—10 cm最低,为8.4%;退耕湿地以土层深度30—40 cm水分含量高,为14.01%,0—10 cm最低,为5.35%;丘间低地以土层深度20—30 cm水分含量高,为12.22%,0—10 cm最低,为4.02%。总的变化规律从高到低排列为固定盐碱沙地>盐减地>退耕湿地>丘间低地。
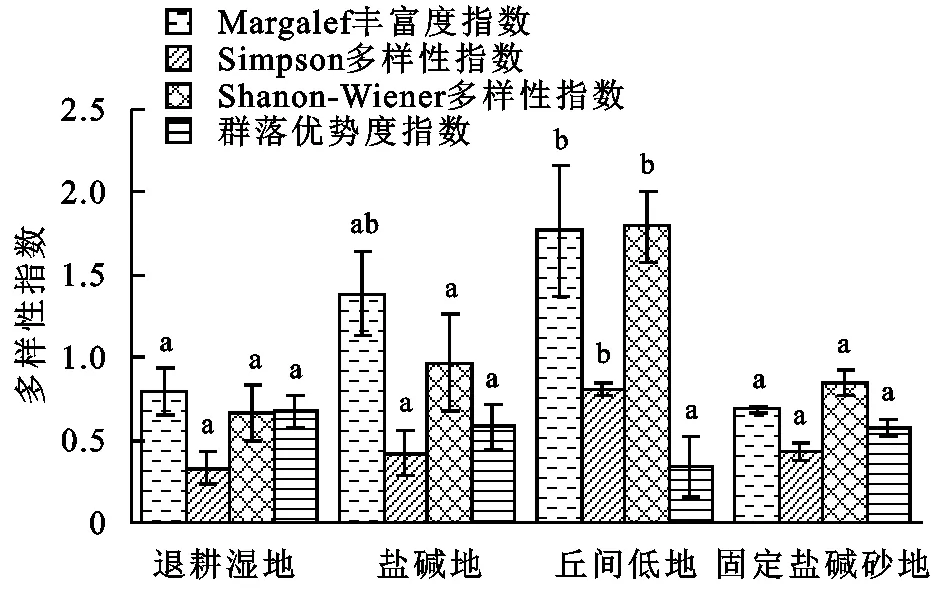
图2 不同湿地类型柽柳灌丛群落多样性指数
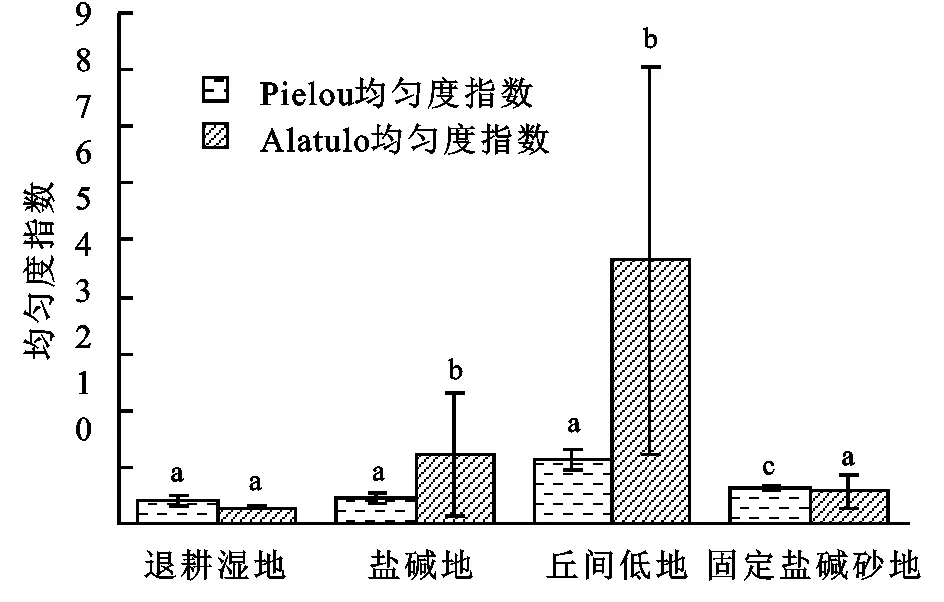
图3 不同湿地类型柽柳灌丛群落均匀度指数
盐碱地平均含盐量最高,为4.01%,固定盐碱沙地次之为3.52%,退耕湿地和丘间低地最低,分别为0.11%和0.11%。从不同土层深度来看,变化差异较大,固定盐碱沙地和盐减地表层0—5 cm盐分含量较高,分别为7.59%和4.66%,丘间低地是中间0—40 cm含量较高,为0.16%,退耕湿地下层40—80 cm含量较高,为0.14%。随着土层深度的不断增加盐分含量呈逐渐下降的趋势。
2.3 不同湿地类型柽柳群落物种多样性指数与土壤水分的相关性
土壤水分在植被生长过程中起主要作用。通过石羊河中下游不同生境类型柽柳群落物种多样性指数与土壤水分的相关性分析(表4),研究结果表明:在不同湿地演替期间各指数与土壤水分间存在不同程度的差异。Simpson指数、Shanon-Wiener指数、群落优势度指数、Pielou指数、Alatulo指数与Margalef指数显著性不明显;Shannon-Wiener指数与Simpson指数呈极显著正相关(p<0.01);群落优势度指数与Simpson指数呈极显著负相关(p>0.01);群落优势度指数与Shannon-Wiener指数呈显著负相关(p>0.05);Pielou指数与Simpson指数和Shannon-Wiener指数分别呈显著正相关(p<0.05);Pielou指数与群落优势度指数呈显著负相关(p>0.05);Alatulo指数与Simpson指数呈显著正相关(p<0.05);Alatulo指数与Shannon-Wiener指数呈极显著正相关(p<0.01);Alatulo指数与群落优势度指数呈显著负相关(p>0.05);Alatulo指数与Pielou指数显著性不明显;土壤平均含水率与Margalef丰富度指数 、Simpson多样性指数、Shanon-Wiener多样性指数、群落优势度指数、Pielou均匀度指数、Alatulo均匀度指数间均呈正相关且均未达到显著水平。这或许是由于研究尺度的差异所引起的,这说明土壤水分有效性的改变,从而影响土壤水分与物种多样性相关关系的强弱,甚至会发生逆转[18]。所以物种组成或物种多样性对生态系统功能的发挥具有十分重要的作用。

图4 不同湿地类型柽柳群落土壤水盐的垂直变化
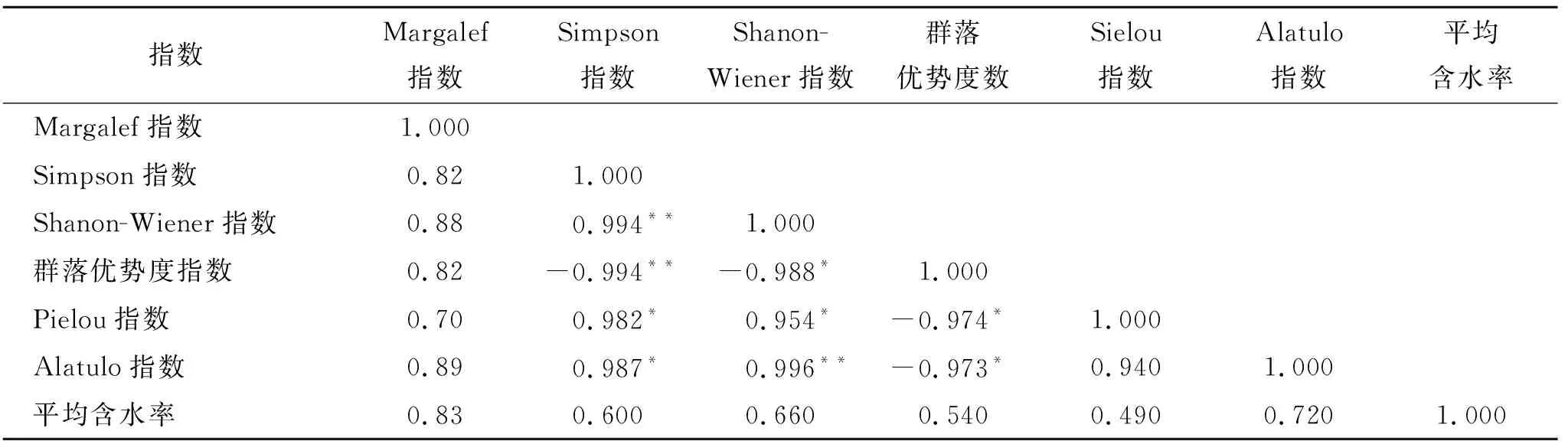
表4 物种多样性与土壤水分的关系
3 讨 论
石羊河中下游不同生境类型,柽柳灌丛在退耕湿地、盐碱地、丘间低地和固定盐碱沙地不同生境类型中,因受地下水埋深度及环境条件的影响,柽柳群落组成物种较少,各群落中共出现物种22种,分属9科21属,各立地条件下,柽柳群落物种主要集中在藜科、禾本科、蒺藜科及柽柳科,其余科物种为单科单属单种,无乔木层和高大灌木层、物种间相比只有较偏高的柽柳灌木层占为优势[19]。在不同生境类型的柽柳群落中,灌木主要局限在柽柳、黑果枸杞、白刺、枸杞、盐爪爪几个单一的物种之间。在不同生境类型柽柳群落中,丘间低地物种最多,退耕湿地原是多年前的弃耕地,随着青土湖水量的增加,现已演变为湿地,物种较少,基本被灌木层所占据,由于种间竞争加剧,草本植物难以存活,退耕湿地和丘间低地的灌木物种种类相同,均为柽柳、黑果枸杞、白刺、枸杞、盐爪爪;盐碱地和固定盐碱沙地灌木物种数目最少,地表有干盐湖沉积,含水率最高。草本植物主要以禾草类和水草类为主。
从不同生境类型柽柳生长指标看,退耕湿地,物种密度、新梢生长和盖度明显低于其他3种类型;丘间低地为人工多年栽植的柽柳,土壤条件相对最优,物种出现最多,新梢生长、相对株高和冠幅显著高于其他3种类型;盐碱沙地中物种次之,柽柳株数最多,密度和盖度较大,存在种内竞争,使得株高和冠幅均不突出,长势一般;固定盐碱沙地中物种较少基本以灌木层为主,表层形成盐分的移动和积聚,水分蒸发快,水分基本集中在中层,长势较好,且冠幅、盖度最高,明显高于其他3种类型。这与刘发民[20]、王玉辉[21]、常兆丰[22]等的研究结果相似,因此,在干旱荒漠地区土壤水分是影响群落结构的主导因子,在不同土壤类型上分布的植物群落,其群落结构和物种多样性存在明显差异,而湿地周围虽土壤含水较多,但盐碱含量也较高,导致群落物种多样性和优势度逐渐减小,为了提高植被覆盖率,要通过人工辅助恢复更新来还原植物群落。
不同湿地类型柽柳群落的物种多样性特征表明:植物群落物种丰富度指数、多样性指数、Simpson多样性指数、 Alatalo均匀度指数在植被恢复过程中呈增大趋势,Pielou均匀度指数、群落优势度指数呈减小的趋势,丘间低地在4种湿地类型中Margalef丰富度指数、Shanon-Wiener多样性指数和Alatalo均匀度指数最高,其次为盐碱沙地和退耕湿地、固定盐碱沙地最低,总体呈先逐步增加然后再缓慢减小趋势。在演变过程中,退耕湿地和固定盐碱沙地群落优势种逐步趋于集中,多年生与一年生草本逐渐消退,基本被灌木层所占据,物种多样性呈下降趋势最后趋于不稳定,盐碱地和丘间低地中,被多年生灌木和草本层占据,物种多样性开始恢复和增加最后趋于稳定,因此,不同生境伴生种草本群落多样性有明显的差异,其中,总盖度、物种数、均匀度指数、多样性指数、丰富度指数随着土壤水分的增加呈逐渐增加趋势,表现出一定的“表聚性”,但在荒漠区有限的降水限制下,植被的恢复是一个滞后的过程,各项多样性指标仍在不断变化,需要经过一段时间才能充分提现出来。
在4种不同湿地类型群落中,随着土层深度的增加,土壤紧实度加大,土壤含水量是固定盐碱沙地和退耕湿地最高,其次为盐碱地和丘间低地,土壤含水量的活跃土层在30 cm或40 cm(土壤相对活跃层),相关性分析表明:柽柳群落物种多样性与土壤含水量均呈正相关性但均未达到显著水平,这与鲁延芳[23]、孟明[24]等的研究结果一致,而与濮阳雪华[25]等的研究结果不同,这或许是由于各研究区域柽柳群落的人为干预和研究尺度的差异引起的结果。物种多样性和土壤水分梯度间关系很复杂,多数研究表明,植物群落物种多样性与土壤含水量间表现出正相关关系[26-28];还有研究表明,植物多样性与年降水量具有正相关关系[29]。但也有研究指出,水分对植物群落物种多样性无显著影响[30],同样,植物生长特性与土壤水分间的关系也存在争议[31],说明在不同水盐养分环境下的显著变化可能对物种多样性、功能多样性二者间关系起决定性作用[32]。在今后的研究中,应加强关于降水梯度与盐分养分协同变化下对植物群落多样性特征的响应研究,深入了解全球变化背景下生态脆弱区域植被恢复规律。
4 结 论
石羊河中下游不同生境带柽柳群落因受环境条件的限制,共有22种,9科,20属,灌木8种主要有柽柳科、茄科和蒺藜科等物种,多年生草本与一年生草本物种数相同,均为7种,主要有藜科和禾本科等物种,多为单科多属单种。不同湿地类型柽柳群落物种多样性指数从高到低依次排列为丘间低地>盐碱地>固定盐碱沙地>退耕湿地,且差异均具有统计学意义(p<0.05),从Pielou均匀度指数看,丘间低地和固定盐碱沙地差异显著(p<0.05),盐碱地和退耕湿地物种差异不显著(p>0.05)。土壤平均含水率与植被群落物种多样性均呈正相关但均未达到显著水平。
