苗期白肋烟和烤烟主要碳水化合物含量差异及形成机理
2022-04-29刘德水冯雨晴李亚飞史素娟马雁军周骏史宏志
刘德水,冯雨晴,李亚飞,史素娟,马雁军,周骏,史宏志*
苗期白肋烟和烤烟主要碳水化合物含量差异及形成机理
刘德水1,冯雨晴2,李亚飞2,史素娟1,马雁军1,周骏1,史宏志2*
1上海烟草集团有限公司技术中心北京工作站,北京 101121;2河南农业大学烟草学院,郑州 450002
为探讨白肋烟和烤烟烟叶中碳水化合物含量差异,揭示其形成机理。以红花大金元(HD)、中烟 100(Z100)、TN90、TN86为材料,通过水培试验,研究烤烟和白肋烟碳水化合物含量、色素含量、光合作用参数及其碳代谢关键基因表达量差异。白肋烟烟叶中色素、总糖和还原糖含量明显低于烤烟,在同一施氮水平条件下,白肋烟烟叶还原糖和总糖含量分别较烤烟低42.89%和43.89%;在相近叶片生物量条件下,白肋烟烟叶还原糖和总糖含量分别较烤烟低64.40%和36.46%。白肋烟色素含量和光合作用效率比烤烟低,且碳代谢关键基因和表达水平均低于烤烟,碳固定和同化能力弱,不利于光合作用产物的形成。白肋烟碳水化合物含量与色素含量和光合作用效率呈现出正相关,相关系数达到极显著水平。色素含量低,光合作用弱,碳固定同化能力弱可能是引起白肋烟主要碳水化合物含量低的关键原因。
烤烟;白肋烟;色素;光合作用;碳同化;碳水化合物
碳水化合物是植物通过光合作用合成的一类重要的有机化合物,是碳代谢过程中的重要产物。烟叶碳水化合物占干物质总量的25%~50%,是烟叶所有化合物中含量最多的一类,在烟草生物体中起重要作 用[1]。其中,水溶性总糖和还原糖是最主要的碳水化合物,也是形成香气物质的重要前体物,它们的含量对烟叶质量及卷烟品质的影响尤为突出[2]。张转勇等[3]研究发现日照时数、降雨量、气温等生态因素影响烟叶中总糖和还原糖的含量。王晶等[4]研究发现,碳代谢旺盛的“豫烟10号”烟叶中总糖和还原糖含量较高。方明等[5]研究表明,磷、钾营养供应不足的烟叶碳代谢过程受到抑制,从而限制光合作用的最初产物-糖的合成。
白肋烟在混合型卷烟叶组配方中的比例高达30%,在调控烟叶感官质量和吸食品质中起重要作用[6]。白肋烟主脉乳白色,叶色黄绿,是一种叶绿素缺乏的缺绿突变体[7]。叶绿素是绿色植物吸收光能和光能转化的重要物质,其中叶绿素a和叶绿素b直接影响作物的光合同化过程[8]。白肋烟烟叶因缺乏叶绿素,且水溶性总糖和还原糖较低(与烤烟相比),因此探究白肋烟烟叶碳水化合物含量低的原因是提高混合型卷烟质量的有效途径。
目前国内外对影响烤烟碳水化合物含量以及提高糖含量的措施进行了广泛的研究,但对苗期不同类型烟草糖含量形成差异及影响机理研究较少。因此,本研究对烤烟和白肋烟烟叶色素含量、光合作用、最大光量子产量等差异因素进行分析,并结合关键基因表达量分析,旨在明确造成白肋烟碳水化合物含量低的主要原因,为提升混合型卷烟的质量提供参考。
1 材料与方法
1.1 实验材料和培养条件
气候室条件设置:温度22~28℃,湿度75%~80%,实行昼夜交叉光照12 h /12 h(光照时间段为8:00—20:00),光照强度为 400 μmol·m-2·s-1。以烤烟品种红花大金元(HD)和中烟100(Z100)、白肋烟品种TN90和TN86为试验材料。用2%的次氯酸钠溶液消毒种子,连续消毒两次,每次消毒5 min,然后播种在育苗盘中。当烟苗长出4~5片真叶(播种40 d后)时,移栽入规格大小为25 cm×30 cm(直径×深度)的塑料盆中,用蛭石固定烟苗开展水培试验。前期研究表明,白肋烟24 mmol/L的供氮条件与烤烟在4 mmol/L时的供氮条件烟叶生物量相近。因此,本研究共设置6个实验处理,分别是4 mmol/L供氮条件下的HD、Z100、TN90、TN86和24 mmol/L供氮条件下的TN86和TN90。每个处理进行3次实验重复,营养液配制采用霍格兰氏法进行。
1.2 测定项目
选择长势均匀一致的烟叶叶片(4 mmol/L和24 mmol/L的供氮条件下培养10 d),每个烟苗选取同一部位的1片烟叶(从上往下数第4片叶),测定净光合速率和叶绿素荧光参数。然后将各处理烟叶平均分成2份,新鲜样品用于色素含量和转录组测定,杀青样品用于烟叶总糖和还原糖测定。
1.2.1 色素含量,光合速率和叶绿素荧光的测定
采用95%乙醇提取测定色素含量[9]。使用便携式光合作用测定系统(LI-COR Biotechnology,6400XT,Lincoln,NE,USA)测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Cd)和胞间二氧化碳浓度(Ci),CO2浓度和光量子通量密度设定为400 μmol·m-2·s-1和1200 μmol·m-2·s-1。将叶片在黑暗条件下处理20 min,然后用Mini-PAM荧光光度计测定叶绿素a荧光。参考Schreiber的方法测定和计算最大光化学量子产量(Fv/Fm)[10]。
1.2.2 总糖和还原糖含量的测量
取样品(0.25 g粉末混合物和25 mL,5%乙酸在50 mL锥形瓶中溶解)震荡30 min后过滤,根据中国烟草行业标准YC/T 161,159—2002用流动分析仪测定总糖和还原糖含量[11]。
1.3 碳代谢关键基因的转录分析
取液氮保存的材料进行总RNA提取。使用Thermo Fisher Scientific M-MLV反转录试剂盒将提取的RNA反转录合成cDNA,并以cDNA为模板,进行qRT-PCR实验。、和的引物序列具体如表1,、、、(烟草)的引物序列同之前报道[12]。反应扩增程序为:95℃ 10 s,95℃ 10 s,60℃ 30 s,40个循环。每个处理设置3次重复。

表1 qRT-PCR 引物序列
1.4 数据分析
采用 Origin pro 9.0 和Excel 2019进行数据分析和图表制作;使用 SPSS 24.0 软件进行方差及相关性分析。将烤烟和白肋烟2个品种的平均值为两个独立样本,用独立样本 T 检验法对烟株干物质积累量、总糖、还原糖、色素、光合速率等指标进行方差齐性检验和 T 检验,对烟叶干物质积累量、总糖和还原糖含量与色素含量和光合速率进行相关性分析。
2 结果与分析
2.1 白肋烟和烤烟烟叶还原糖和总糖含量差异
白肋烟和烤烟中还原糖和总糖含量的测定结果如图1所示,在同一施氮水平条件下,白肋烟(TN90和TN86)烟叶还原糖和总糖含量的平均值分别较烤烟(HD和Z100)低42.89%和43.89%;在相近叶片生物量条件下,白肋烟(TN90和TN86)烟叶还原糖和总糖含量的平均值分别较烤烟(HD和Z100)低64.40%和36.46%。由此可见,无论是同一施氮水平还是相近生物量条件,白肋烟烟叶碳水化合物的含量明显低于烤烟。
2.2 白肋烟和烤烟烟叶色素含量差异
白肋烟和烤烟叶片叶绿素含量的测定结果显示(图2),在相同施氮水平下,烤烟两品种烟叶叶绿素a含量、叶绿素b含量和类胡萝卜素平均值分别较白肋烟高70.83%、82.76%和50.00%,且差异达极显著水平(<0.01)。在叶片生物量相近条件下,白肋烟烟叶的类胡萝卜素含量明显高于烤烟(类胡萝卜素含量受施氮量影响较大),而其烟叶叶绿素a和叶绿素b含量与烤烟无显著差异。由此说明,相同施氮条件下,白肋烟烟叶色素含量显著低于烤烟,随着施氮量增加,白肋烟烟叶色素总量、叶绿素a、叶绿素b和胡萝卜素含量均呈增加趋势。

注: *1或**1代表在相同的施氮量下,白肋烟两品种(TN90和TN86)与烤烟品种(HD和Z100)进行比较,差异已达到显著或极显著水平(P < 0.05或P < 0.01)。*2或**2代表在相近叶片生物量条件下,白肋烟两品种(TN90和TN86)与烤烟品种(HD和Z100)进行比较,差异已达到显著或极显著水平(P <0.05或P <0.01)。下同。
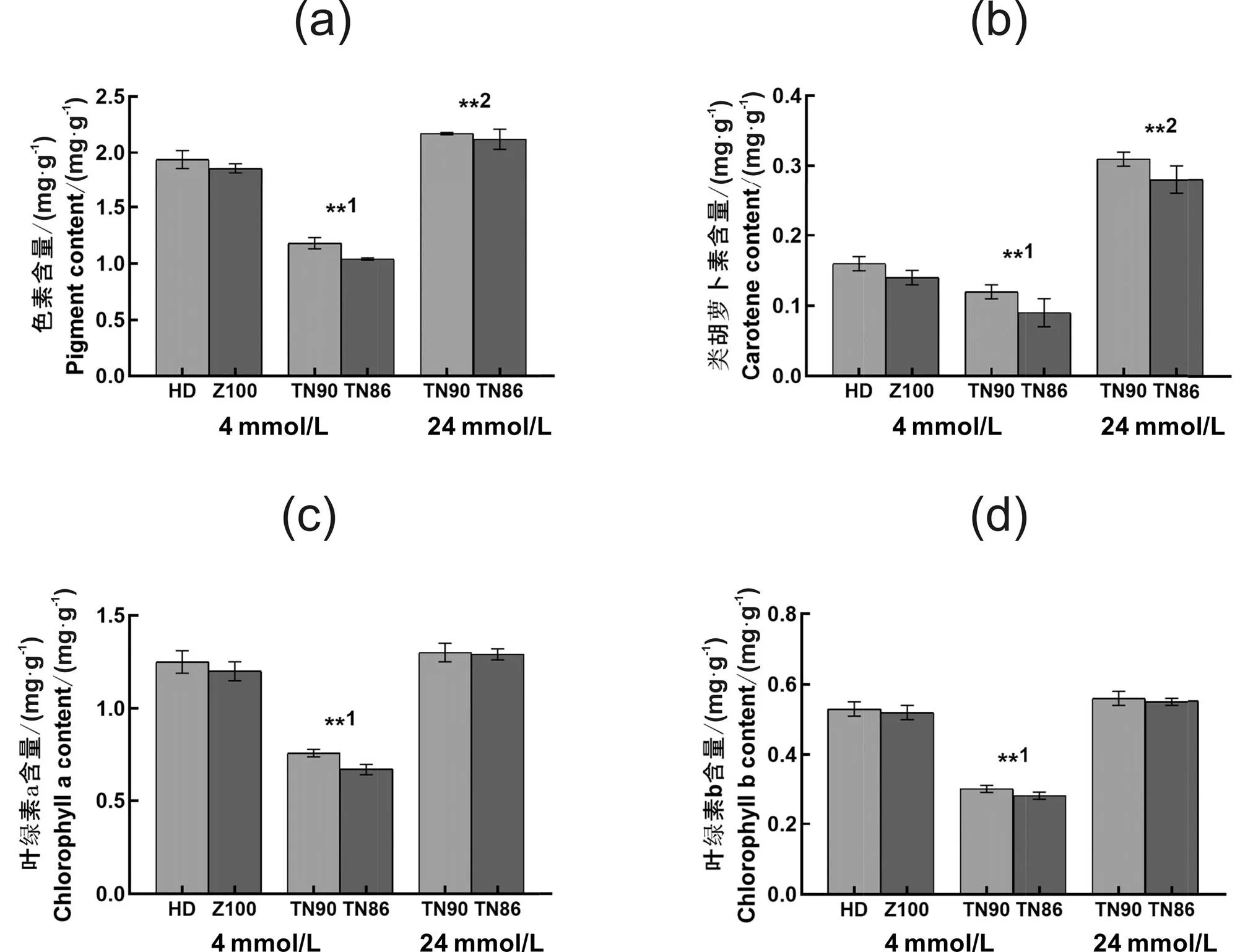
图2 白肋烟和烤烟烟叶色素含量差异
2.3 白肋烟和烤烟烟叶最大光量子产量差异
最大光量子产量(Fv/Fm)反映植物潜在最大光合能力,可代表叶绿体中的光系统II(PSⅡ)的最大光化学效率,较高的光化学效率有利于光合色素捕获光能并以最大的效率转化为化学能,从而获得更多的能量。白肋烟和烤烟叶片Fv/Fm的测定结果显示(图3),在同一施氮水平和相近叶片生物量条件下,白肋烟烟叶最大光量子产量与烤烟烟叶相差不大。由此说明,白肋烟碳水化合物含量与烟叶最大光量子产量无关。

注: ns代表处理间无显著性差异。
2.4 白肋烟和烤烟光合参数差异
白肋烟和烤烟光合参数的测定结果如图4所示,在相同施氮条件下,烤烟两品种净光合速率(Pn)、气孔导度(Cd)、CO2胞间浓度(Ci)、蒸腾效率(Tr)的平均值分别是白肋烟的1.31倍、3.00倍、2.24倍和3.65倍。叶片生物量积累相近条件下,烤烟烟叶Pn、Ci与白肋烟无显著差异,但Tr仍显著高于白肋烟,其平均值为白肋烟的1.70倍。由此可见,白肋烟和烤烟之间光合速率存在明显差异,且差异达到显著或极显著性水平。
2.5 相关性分析
在相同施氮条件下,白肋烟和烤烟烟叶总糖和还原糖含量与叶绿素a含量、色素含量和净光合速率呈正相关,且相关系数达到极显著水平。由此可知,烟叶中碳水化合物含量与色素含量和光合作用关系密切,提高白肋烟中色素含量和光合速率有助于提高烟叶中碳水化合物含量。

图4 白肋烟和烤烟光合参数差异

表2 碳水化合物含量与色素含量、叶绿素a和净光合速率的相关性分析
2.6 白肋烟和烤烟烟叶碳代谢差异基因分析
白肋烟和烤烟碳代谢关键基因表达量的分析结果显示(图5),在同一施氮量和相近叶片生物量积累条件下,白肋烟烟叶中光合碳固定和糖类物质合成相关基因()表达水平均比烤烟低。其中,相同施氮量,烤烟和基因表达量分别是白肋烟的6.22倍、4.47倍,叶片生物量相近条件下,烤烟和基因表达量分别是白肋烟的3.76倍和3.66倍。由此说明,白肋烟烟叶光合碳固定能力弱,蔗糖和淀粉合成能力低,从而导致白肋烟烟叶糖含量低。

注:(a),基因EGY1 (Gene Probable zinc metalloprotease);(b),基因CP12-2 (Gene Calvin cycle protein);(c),基因SUS2(Gene Sucrose synthase);(d),基因gpmA(Gene 2,3-bisphosphoglycerate-dependent phosphoglycerate mutase);(e),基因AGPS1(Gene Glucose-1-phosphate adenylyltransferase large subunit 1);(f),基因GDHA(Gene Glutamate dehydrogenase A)。
3 讨论
不同烟草品种烟叶碳水化合物含量差异显著,其中白肋烟烟叶中总糖和还原糖含量与烤烟有较大差异[5]。本研究结果显示,在相同施氮量和相近生物量条件下,白肋烟两品种中烟叶总糖和还原糖含量明显低于烤烟品种,且受碳固定同化过程的关键因子影响。
光合作用是绿色植物重要的生化反应过程,其中,PSII作为光合作用的主要场所,可以直接捕获光能并完成水的光解和质体醌的还原,对于光合作用的开启具有重要作用[13]。编码一个定位于类囊体膜上的金属蛋白酶,突变体表现为叶色黄绿,叶绿体和光合系统II稳定性下降,叶绿素含量降低,从而影响植物生长[14-15]。编码叶绿体基质中的一种小肽,且CP12蛋白可与甘油醛-3-磷酸脱氢酶(GAPDH)和磷酸核酮糖激酶(PRK)(卡尔文循环的两种关键酶)形成一种超分子复合物,从而调节光合作用[16]。研究表明,的转录物在决定叶片光合作用能力方面起重要作用[17]。Marri等也发现,CP12蛋白氧化对植物不同环境条件下光合过程的整体平衡至关重要[18]。叶绿素含量降低在一定程度上能抑制叶绿体发育,降低光合速率,影响植物体的有机物质积累、生长发育和产量形成[19]。拓阳阳等[20]研究发现,叶绿素a与总糖含量呈显著正相关。本研究表明,在相同施氮水平下,白肋烟烟叶色素总含量明显低于烤烟,且差异达极显著水平,与其光合速率低于烤烟的表现一致。基因表达量结果表明,同一施氮量和相同生物量条件下,白肋烟和基因表达量显著低于烤烟,导致其色素含量和光合作用效率低,进一步影响白肋烟烟叶中总糖和还原糖含量。
蔗糖与淀粉代谢途径中关键酶基因的表达,与烟叶中糖类物质的含量密切相关。本研究结果发现,白肋烟烟叶蔗糖和淀粉代谢途径关键基因表达量低于烤烟,可能是其糖含量低的重要原因。蔗糖合成酶(SUS)是植物中广泛存在的一种UDP-糖基转移酶,也是催化蔗糖代谢的关键酶,在种子和果实的发育中发挥作用[21]。研究发现,过量表达基因的转基因烟草中可溶性总糖含量显著高于野生植株[22]。由基因编码的磷酸甘油酸变位酶是糖异生和糖酵解过程中的重要产物,在光合碳固定过程发挥重要作用[23]。焦磷酸化酶(AGPase)是淀粉合成过程中的限速酶,赵凯琴等[24]研究表明过表达外源AGPase基因,可增加油菜胚淀粉的合成。本试验中,相同施氮量和相同生物量条件下,白肋烟可溶性总糖和还原糖含量显著低于烤烟,与蔗糖和淀粉合成途径中关键基因和表达量低密切相关。
谷氨酸脱氢酶(GDH)是烟叶氮代谢的关键酶,通过提高氮同化能力来增加酶及叶绿素的积累量,从而间接影响植物的碳同化。氮同化过程利用碳代谢合成的碳架合成蛋白质和氨基酸,GDH可在碳架不足时通过脱氨作用提供碳骨架[25-26]。申丽霞等[27]研究认为较高的GDH活性有利于催化和加速合成谷氨酸,维持叶片较强的碳代谢。Egami等[28]发现转基因马铃薯植株的叶片在开花期的可溶性蛋白质浓度和光合作用效率均增加。本研究中,相同施氮量和相近生物量下白肋烟烟叶总糖和还原糖含量和基因表达量均低于烤烟,推测白肋烟烟叶糖含量低以及氮同化过程中碳架不足是导致烟叶硝酸盐含量和氮素利用效率低于烤烟的原因,这对提高烟叶安全性和减肥增效不利,还有待进一步研究。
4 结论
在相同施氮水平和相近叶片生物量积累条件下,白肋烟烟叶中还原糖和总糖含量明显低于烤烟,并且色素含量、光合作用均低于烤烟。白肋烟碳固定和糖类物质合成相关基因(、、、、、)表达量显著降低;相关性分析结果显示,总糖和还原糖含量与色素含量和净光合速率呈显著正相关。色素含量低,光合作用弱,碳固定同化能力弱可能是引起白肋烟碳水化合物含量低的关键原因。
[1] 韩锦峰. 烟草栽培生理[M]. 中国农业出版社,1996.
HAN Jinfeng. Physiology of tobacco cultivation [M]. China Agriculture Press,1996.
[2] 曹景林,程君奇,李亚培,等. 烤烟常规化学成分与吸食品质关系的研究进展[J]. 湖北农业科学,2020,59(S1): 253-258+262.
CAO Jinglin, CHENG Junqi, LI Yapei, et al. Research progress on the relationship between routine chemical composition and smoking quality of flue-cured tobacco[J]. Hubei Agricultural Sciences, 2020, 59(S1): 253-258+262.
[3] 张转勇,冯圭如,王婷,等. 烤烟糖质量分数与气象因子的相关性评价——以昭通烟区为例[J]. 西南师范大学学报(自然科学版),2019,44(12): 54-58.
ZHANG Zhuanyong, FENG Guiru, WANG Ting, et al. On correlation analysis between sugar content of flue-cured tobacco and meteorological factors: A case study of Zhaotong tobacco- growing areas[J]. Journal of Southwest China Normal University (Natural Science Edition),2019,44(12):54-58..
[4] 王晶. 不同生态区烟叶碳水化合物合成及积累规律的比较研究[D]. 郑州:河南农业大学,2013.
WANG Jing. Research on the dynamic synthesis and accumulation regulation of carbohydrate on tobacco leaves of different ecological zone[D]. Henan Agricultural University, 2013.
[5] 方明,符云鹏,刘国顺,等. 磷钾配施对晒红烟碳氮代谢和光合效率的影响[J]. 中国烟草科学,2007(02): 27-30.
FANG Ming, FU Yunpeng, LIU Guoshun, et al. Effects of P and K fertilizers on the carbon and nitrogen metabolism and photosynthetic efficiency of dark sun-cured tobacco[J]. Chinese Tobacco Science, 2007(02):27-30.
[6] 刘开楠. 白肋烟烘焙工艺研究[D]. 长沙:湖南农业大学,2014.
LIU Kainan. Study on the baking techniques of burley tobacco[D]. Hunan Agricultural University, 2014.
[7] 何其芳. 94份烟草材料的遗传多样性和主要品质成分的分析[D]. 广州:华南理工大学,2012.
HE Qifang. Analysis of genetic diversity for 94 tobacco cultivars and the main quality components of these[D]. South China University of Technology, 2012.
[8] 彭方仁,朱振贤,谭鹏鹏,等. NaCl胁迫对5个树种幼苗叶片叶绿素荧光参数的影响[J]. 植物资源与环境学报,2010, 19(03): 42-47.
PENG Fangren, ZHU Zhenxian, TAN Pengpeng, et al. Effect of NaCl stress on chlorophyll fluorescence parameter in leaf of five tree species seedling[J]. Journal of Plant Resources and Environment, 2010, 19(03): 42-47.
[9] 邹琦. 植物生理学实验指导[M]. 北京:中国农业出版社,2000: 56-59.
ZOU Qi. Experimental guidance for plant physiology[M]. Beijing: China Agricultural Press, 2000:56-59.
[10] Schreiber U, Bilger W, Neubauer C.Chlorophyll fluorescence as a non-intrusive indicator for rapid assessment ofphotosynthesis[J].,1995, 100: 49-70.
[11] YC/T 159—2002烟草及烟草制品水溶性糖的测定连续流动法[S].
YC/T 159-2002 Tobacco and tobacco products-determination of water-soluble sugars-continuous flow method[S].
[12] 冯雨晴,李亚飞,史宏志. 叶面喷施丙三醇对烟叶碳氮代谢及硝酸盐积累的影响[J]. 中国烟草学报,2019, 25(05): 53-62.
FENG Yuqing,LI Yafei,SHI Hongzhi. Effect of foliar application of glycerol on carbon and nitrogen metabolism and nitrate accumulation in tobacco leaves [J].Acta Tatacaria Sinica, 2019,25(05): 53-62.
[13] Li X, Wang H B, Jin H L. Light signaling-dependent regulation of PSII biogenesis and functional maintenance[J]. Plant Physiology, 2020, 183(4): 1855-1868.
[14] Qi Y, Wang X, Lei P, et al. The chloroplast metalloproteases VAR2 and EGY1 act synergistically to regulate chloroplast development in Arabidopsis[J]. Journal of Biological Chemistry, 2020, 295(4): 1036-1046.
[15] ADAMIEC M, MISZTAL L, CIESIELSKA M, et al. The changes of PSII supercomplex stoichiometry inmutants are related to chlorophyll b deficiency[J]., 2021, 59 (2): 294-302.
[16] Gurrieri L, Fermani S, Zaffagnini M, et al. Calvin-Benson cycle regulation is getting complex[J]. Trends in Plant Science, 2021, 26(9).
[17] López C, Abuzaid O, Lawson T, et al.CP12 mutants have reduced levels of phosphoribulokinase and impaired function of the calvin-benson cycle. Journal of Experimental Botany, 2017, 68: 2285-2298.
[18] Marri L, Thieulin P G, Lebrun R, et al. CP12-mediated protection of Calvin-Benson cycle enzymes from oxidative stress., 2014, 97: 228-237.
[19] 彭门路,赵小珍,王晓东,等. 甘蓝型油菜叶绿素缺失突变体1的表型鉴定与遗传分析[J]. 中国油料作物学报,2021, 43(3): 443-451.
PENG Menlu, ZHAO Xiaozhen, WANG Xiaodong, et al. Phenotypic identification and genetic analysis of a chlorophyll deficient muantin Brassicanapus[J]. Chinese Journal of oil Crop Sciences, 2021, 43(3):443-451.
[20] 拓阳阳,赵铭钦,金洪石,等. 烤烟不同基因型间质体色素含量差异与烟叶化学成分的关系[J]. 中国烟草科学,2011, 32(06): 32-36.
TUO Yangyang, ZHAO Mingqin, JIN Hongshi, et al. The Relationship between the difference of plastid pigment contents and chemical components in different flue-cured tobacco varieties[J]. Chinese Tobacco Science, 2011, 32(06): 32-36
[21] Xu S M, Brill E, Llewellyn D J, et al. Overexpression of a potato sucrose synthase gene in cotton accelerates leaf expansion, reduces seed abortion, and enhances fiber production[J]. Molecular Plant, 2012, 5(2): 430-441.
[22] Nguyen Q A, Luan S, Wi S G, et al. Pronounced phenotypic changes in transgenic tobacco plants overexpressing sucrose synthase may reveal a novel sugar signaling pathway[J]. Frontiers in plant science, 2016, 6: 1216.
[23] Bond C S, White M F, Hunter W N. Mechanistic implications forcofactor-dependent phosphoglycerate mutase based on the high-resolution crystal structure of a vanadate complex[J]. Journal of Molecular Biology, 2002, 316(5): 1071- 1081.
[24] 赵凯琴,杨清辉,张云云,等. 过表达AGPase编码基因对甘蓝型油菜种子淀粉含量及油脂合成的影响[J]. 中国油料作物学报,2021,43(03): 426-434.
ZHAO Kaiqin, YANG Qinghui, ZHANG Yunyun, et al. Effects of AGPase gene Glgc overexpression on starch and oil synthesis in rapeseed ()[J]. Chinese Journal of oil Crop Sciences, 2021, 43(03):426-434.
[25] MIYASHITA Y, GOOD A G. NAD(H)-dependent glutamate dehydrogenase is essential for the survival ofduring dark-induced carbon starvation[J]. Journal of Experimental Botany, 2008, 59(3): 667-680
[26] 龚茵茵,燕璐,林建中,等. 低等生物谷氨酸脱氢酶基因用于作物遗传改良的研究进展[J]. 生命科学研究,2021,25(01): 31- 38.
GONG Yinyin, YAN Lu, LIN Jianzhong, et al. Research advances on glutamate dehydrogenase genes of lower organisms for genetic improvement[J]. Life Science Research, 2021, 25(01): 31-38.
[27] 申丽霞,王璞. 玉米穗位叶碳氮代谢的关键指标测定[J]. 中国农学通报,2009, 25(24): 155-157.
SHEN Lixia, WANG Pu. Determination of C-N indices in ear-leaf of maize() [J]. Chinese Agricultural Science Bulletin, 2009, 25(24): 155-157.
[28] EGAMI T, WAKAYAMA M, AOKI N, et al. The effects of introduction of a fungal glutamate dehydrogenase gene () on the photosynthetic rates, biomass, carbon and nitrogen contents in transgenic potato[J]. Plant Biotechnology, 2012, 29(1): 57-64.
Difference in carbohydrate content between burley tobacco and flue-cured tobacco and the related formation mechanism
LIU Deshui1, FENG Yuqing2, LI Yafei2, SHI Sujuan1, MA Yanjun1, ZHOU Jun1, SHI Hongzhi2*
1 Technology Center, Shanghai Tobacco Group Beijing Cigarette Factory Co., Ltd., Beijing 101121, China;2 College of Tobacco,Henan Agricultural University,Zhengzhou 450002,China
To investigate the difference in carbohydrate content between burley tobacco and flue-cured tobacco leaves, pots experiments were carried out to reveal the formation mechanism within.By using flue-cured tobacco (HD, Z100) and burley tobacco (TN90, TN86) as research objects, the hydroponic experiments were conducted to study differences in pigments, photosynthesis, carbohydrates and transcriptome.The results showed that the contents of pigment, reducing sugar and total sugar in flue-cured tobacco was significantly higher than that in burley tobacco. Under same nitrogen level, the content of reducing sugar and total sugar of burley tobacco were lower than that of flue-cured tobacco by 42.89% and 43.89% respectively. Under the same leaf biomass, the contents of reducing sugar and total sugar of the burley tobacco were 36.46% and 64.40% lower than that of flue-cured tobacco, respectively. Moreover, compared with flue-cured tobacco, burley tobacco had lower pigment content and photosynthetic efficiency, and weaker carbon fixation and assimilation ability. Also, genes () expression level were all significantly lower in burley tobacco than in flue-cured tobacco. Besides, correlation coefficient between total sugar content, reducing sugar content and pigment content, photosynthesis were significantly positive.Low pigment content, weak photosynthesis, weak carbon fixation assimilation ability were the key reasons for low content of carbohydrate in burley tobacco.
burley tobacco; flue-cured tobacco; pigment; photosythesis; carbon assimilation; carbohydrate
. Email:shihongzhi88@163.com
刘德水,冯雨晴,李亚飞,等. 苗期白肋烟和烤烟主要碳水化合物含量差异及形成机理[J]. 中国烟草学报,2022,28(2).LIU Deshui, FENG Yuqing, LI Yafei, et al. Difference in carbohydrate content between burley tobacco and flue-cured tobacco and the related formation mechanism [J]. Acta Tabacaria Sinica, 2022,28(2).doi:10.16472/j.chinatobacco.2021.158
上海烟草集团北京卷烟厂有限公司科技项目“烟草硝酸盐积累机理及降低硝酸盐含量调节技术研究”(TP2019-C4)
刘德水(1988—),博士研究生,Email:deshui@yeah.net
史宏志(1963—),博士,教授,主要从事烟草栽培生理研究,Email:shihongzhi88@163.com
2021-08-17;
2021-12-17
