千穗谷TCP基因家族生物信息学分析
2022-04-25喻望晨刘宁芳胡龙兴
周 涛, 彭 辉, 喻望晨, 刘宁芳, 胡龙兴, 徐 倩
(湖南农业大学农学院草业科学系, 湖南 长沙 410128)
籽粒苋是苋科苋属一年生草本粒用植物的统称,包括千穗谷(Amaranthushypochondriacus)、老鸦谷(A.cruentus)、尾穗苋(A.caudatus)、绿穗苋(A.hybridus)等。千穗谷为籽粒苋的一种,起源于美洲,目前主要分布于温带和热带地区,在平原和高海拔地区均有分布。千穗谷具有根系发达、吸肥力强、生长快、产量高、营养丰富等特点,是一种粮饲兼用型作物,其茎叶可作蔬菜,种子可食用或酿酒,也可栽培观赏[1-3]。千穗谷耐酸、耐盐碱,具有较强的抗旱性和耐受重金属的能力,可作为土壤修复和生态恢复类型植被[4-6]。
TCP(Teosinte branched1/cycloidea/proliferating cell factor)是一类植物特有的转录因子家族,参与植物多种器官的生长发育与形态建成。目前已在拟南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、蒺藜苜蓿(Medicagotruncatula)、番茄(Solanumlycopersicum)、胡杨(Populuseuphratica)和白菜(Brassicarapa)中分别鉴定到24,30,21,30,36和39个TCP成员[7-11]。TCP的命名来自于三个蛋白的首字母缩写——玉米(Zeamays)中参与顶端优势调节的TEOSINTE BRANCHED1(TB1)、金鱼草(Antirrhinummajus)中控制花不对称性的CYCLOIDEA(CYC)以及水稻中参与分生组织表达的PROLIFERATING CELL FACTOR1(PCF1)和PCF2[12-14]。TCP蛋白的N端均包含一个高度保守的TCP结构域,由59个氨基酸构成,属于非典型的bHLH结构[13]。根据该结构域氨基酸序列的差异,该家族可分为两类:ClassⅠ(PCF)和ClassⅡ[15],其中,ClassⅡ可以进一步分为CIN和CYC/TB1两个分支[16]。与ClassⅡ亚家族相比,ClassⅠ在bHLH结构的Basic区域有一个四氨基酸的缺失[17]。
TCP转录因子在植物的不同生长发育阶段均有重要作用[18-19],包括叶片形态建成[20]、花器官发育[21]、根的生长和分蘖形成[22]等。白菜中的BrTCP7通过促进茉莉酸生物合成基因BrOPR3和叶绿素分解代谢基因BrRCCR的表达,从而促进叶片衰老[23]。毛竹(Phyllostachysedulis)中PeTCP10基因可通过MeJA途径负调控侧根的生长发育[24]。在拟南芥花中,AtTCP15通过调控IAA合成基因YUC1和YUC4影响IAA的合成[25]。近年来,越来越多的研究发现,TCP转录因子可以提高植物对多种生物胁迫和非生物胁迫的抗性。比如,香蕉(Musanana)TCP基因家族大部分基因响应低氮胁迫的诱导,并且调节香蕉缺氮时的逆境响应[26]。马铃薯(Solanumtuberosum)TCP转录因子StTCP13能提高大肠杆菌对高盐胁迫的耐受性[27]。拟南芥CYC分支成员AtTCP18蛋白能和植原体效应因子SWP1互作,参与植物防御进程[28]。水稻中过表达OsTCP21基因可增强水稻对RRSV病毒的抵抗力[29]。
microRNAs(miRNAs)是一类内源性的小分子非编码RNA[30],在基因表达过程中起负调控作用[31]。前人研究发现miR319的主要靶基因为TCP基因,且以ClassⅡ亚家族中的CIN分支基因成员为主,通过抑制它们的表达调控植物的生长发育以及对逆境的响应[32]。比如,水稻中miR319通过调控OsTCP21和OsGAmyb基因的表达使水稻分蘖减少、籽粒产量降低[33];梅(Prunusmume)中发现的Pm-miR319a可以通过调控靶基因PmTCP4的表达间接调控梅花雌蕊的发育[34]。
千穗谷最新基因组于2017年发布[35],但关于千穗谷TCP基因家族的系统研究未见报道。本研究拟采用生物信息学手段,对千穗谷TCP基因家族进行鉴定和特征分析,包括基因序列特征、结构域比较、蛋白理化性质预测、近源物种系统进化分析、染色体定位、启动子顺式作用元件预测、组织特异性表达等多个方面,为千穗谷TCP基因的功能研究和千穗谷分子育种提供重要的理论依据。
1 材料与方法
1.1 千穗谷TCP基因家族成员的鉴定
从Plant TFDB v5.0(http://planttfdb.cbi.pku.edu.cn)数据库中下载千穗谷TCP蛋白序列,并在Phytozome v13(https://phytozome-next.jgi.doe.gov/)数据库中进行blast搜索,得到基因序列全长。通过在线工具HMMER(https://www.ebi.ac.uk/Tools/hmmer/)分析所有蛋白的结构域,保留所有含有典型TCP结构域的基因。
1.2 千穗谷TCP 基因及蛋白结构分析
利用ExPASy网站(http://web.expasy.org/compute_pi/)预测千穗谷TCP蛋白的分子量(Molecular weight,MWs)、等电点(Isoelectric point,pIs)、平均亲水性(Grand average of hydropathicity)等信息。利用GSDS2.0(http://gsds.gao-lab.org/index.php)在线网站绘制千穗谷TCP的外显子和内含子结构。利用软件MEME[36]预测蛋白保守Motif,使用TBtools[37]软件绘制千穗谷TCP基因家族成员的基因结构图。
1.3 多序列比对和系统发育树构建
利用Clustal W[38]软件对千穗谷、拟南芥、水稻的蛋白序列进行多重序列比对。千穗谷多重序列比对结果使用GeneDoc软件进行展示,分析不同家族的特征结构域。系统发育树采用Mega7[39]软件构建,算法采用邻近法,使用Evolview(https://www.evolgenius.info/evolview/)展示构建的系统发育树。拟南芥以及水稻TCP基因家族数据下载于Plant TFDB v5.0(http://planttfdb.cbi.pku.edu.cn)。
1.4 基因染色体定位和蛋白亚细胞定位
通过BioEdit软件对千穗谷每一个TCP家族成员进行本地BLAST搜索比对,以确定其在染色体中的定位,通过MG2C v2.1(http://mg2c.iask.in/mg2c_v2.1/)在线网站对TCP家族成员染色体定位结果进行图形化处理。千穗谷全基因组数据下载于Phytozome数据库(Amaranthushypochondriacusv2.1,DOE-JGI,http://phytozome.jgi.doe.gov/)。通过在线分析工具WoLF PSORT(https://wolfpsort.hgc.jp/)对千穗谷TCP蛋白的亚细胞定位进行预测。
1.5 千穗谷TCP基因启动子顺式作用元件预测
利用PlantCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线网站对千穗谷TCP基因的启动子进行顺式作用元件预测,使用TBtools软件展示预测结果[37]。
1.6 千穗谷TCP基因组织特异性表达分析
从Phytozome数据库获取千穗谷TCP基因在不同发育时期不同组织器官的表达情况[40],通过Heml软件对不同基因的表达量进行分层聚类分析并绘制热图。
1.7 调控千穗谷TCP基因的miRNA预测
利用psRNATarget(https://www.zhaolab.org/psRNATarget/home)在线网站对可能调控千穗谷TCP基因编码区的miRNA进行预测。由于目前并没有已报道的千穗谷miRNA,因此选择双子叶模式植物拟南芥miRNA数据。
2 结果与分析
2.1 千穗谷TCP基因家族的鉴定及特性分析
从Plant TFDB v5.0数据库共检索得到16个千穗谷TCP基因,分别命名为AhTCP1~AhTCP16(表1)。AhTCP基因长度介于270~1 410 bp之间,氨基酸序列长度在90~470之间,最长为AhTCP12,最短为AhTCP10。蛋白分子量介于9 790.9~50 986.9kD之间;理论等电点介于6.26~10之间,且仅有AhTCP2,AhTCP4,AhTCP8,AhTCP9,AhTCP11的理论等电点小于7,其他蛋白等电点均在碱性范围内;平均亲水性指标均为负值,表明AhTCP成员均是亲水性蛋白。亚细胞定位预测的结果显示,所有千穗谷TCP蛋白在细胞核中定位的概率最高,这与其转录因子的功能相符合。
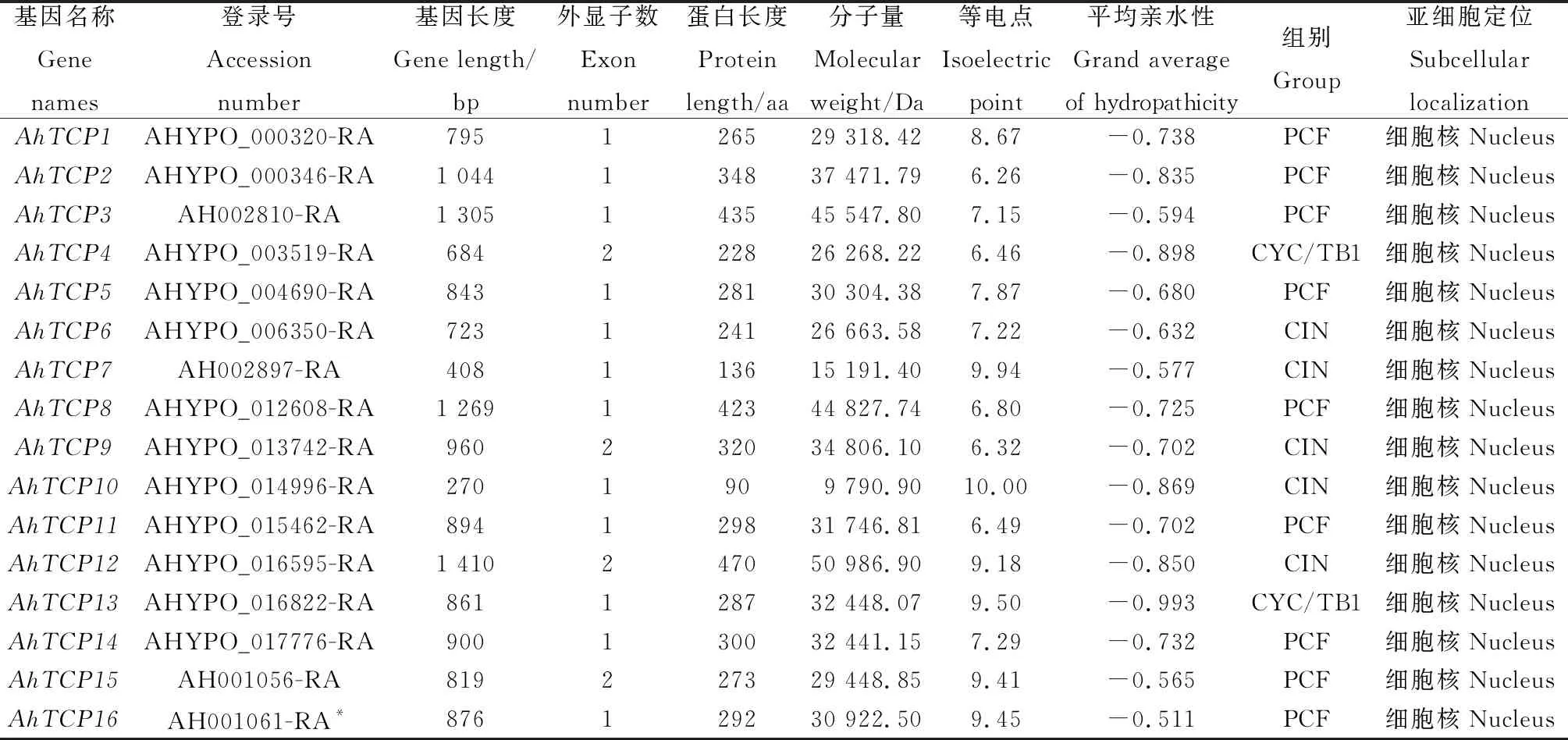
表1 千穗谷TCP 家族成员基本信息
2.2 千穗谷TCP蛋白多重序列比对与系统进化树构建
对千穗谷TCP蛋白序列进行多重序列比对(图1),发现16个成员的TCP结构域呈现不同的特点。典型的TCP结构域属于螺旋-环-螺旋(bHLH)结构,其中AhTCP10在helixII区域有部分缺失,与拟南芥的AtTCP6相似。千穗谷TCP家族中有9个成员在Basic区域缺失了4个氨基酸,与其他物种TCP家族中的PCF亚家族类似,根据这一特征可以将千穗谷16个TCP基因分为两个亚家族:ClassⅠ和ClassⅡ。根据Helix和Loop区域的序列差异(图2),可以进一步将ClassⅡ分为CYC/TB1和CIN两个亚类。在千穗谷中,CYC/TB1亚类特有的保守氨基酸包括Basic区域和HelixI区域之间的亮氨酸,HelixI区域的异亮氨酸和精氨酸,以及Loop区域的苯丙氨酸、赖氨酸和丙氨酸等;而CIN亚类中特有的保守氨基酸包括Basic区域的赖氨酸,HelixⅠ区域的苏氨酸、异亮氨酸、谷氨酰胺和精氨酸,以及Loop区域的脯氨酸等。不同的亚类在HelixII区域的序列差异较大,多态性较高。
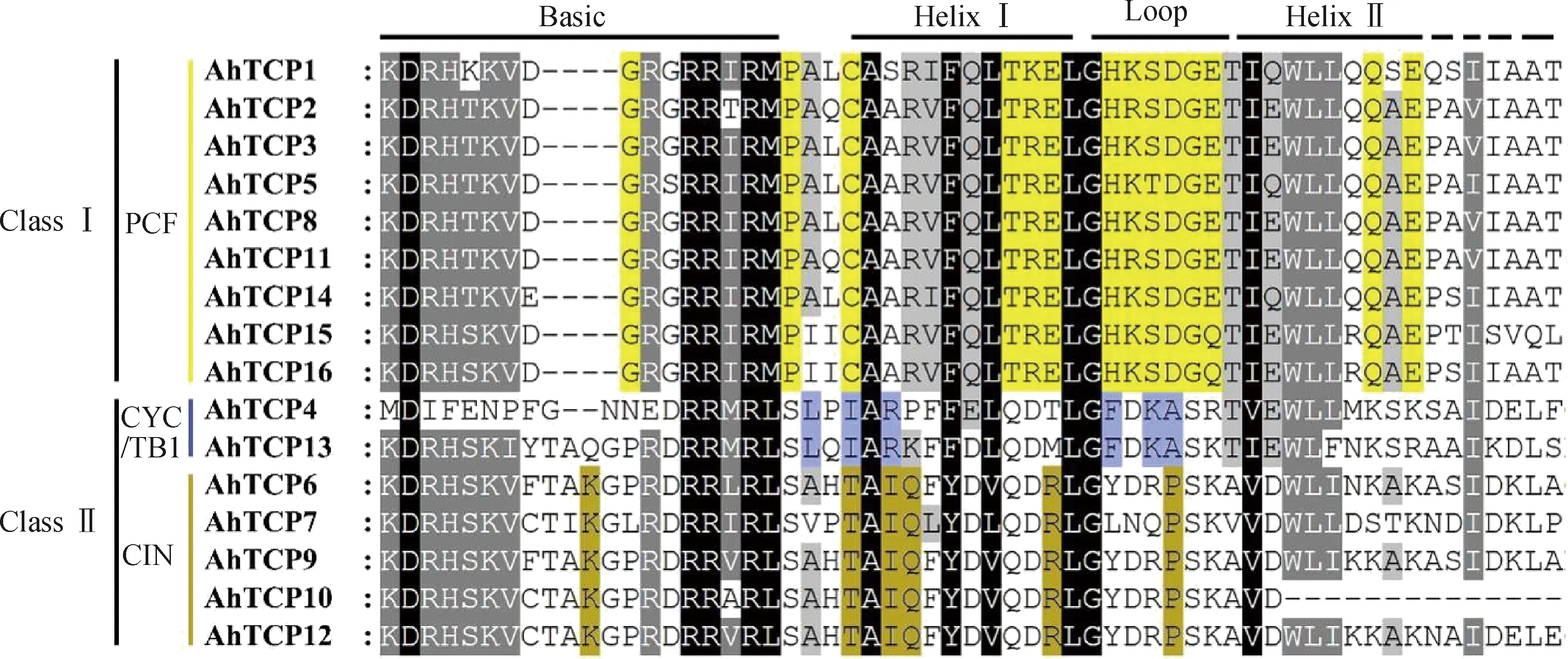
图1 千穗谷TCP蛋白保守结构域序列对比
对千穗谷、拟南芥、水稻三个物种的TCP转录因子构建系统发育树(图2),发现TCP家族成员可分为三大类。根据序列特征,将三大类分别命名为PCF,CYC/TB1和CIN,如图2所示。其中,PCF亚类包含了16个拟南芥基因、10个水稻基因和9个千穗谷基因;CYC/TB1亚类包含了5个拟南芥基因、3个水稻基因和2个千穗谷基因;CIN亚类包含了12个拟南芥基因、10个水稻基因和5个千穗谷基因。千穗谷的TCP家族中一半以上成员为PCF亚类,而拟南芥和水稻中ClassⅠ和ClassⅡ两个亚类的成员数目几乎相同。
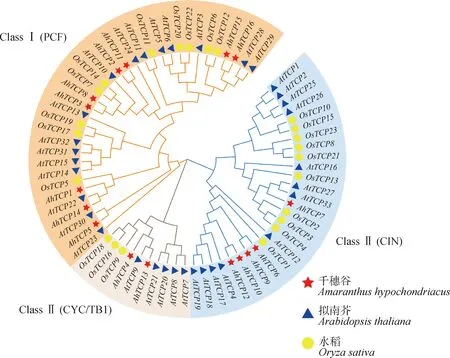
图2 千穗谷、拟南芥和水稻的系统发育树
2.3 千穗谷TCP基因结构和保守基序分析
千穗谷TCP家族成员的基因结构比较简单,大部分AhTCP基因不含内含子,只有4个成员(AhTCP4,AhTCP9,AhTCP12,AhTCP15)含有1个内含子,占总数的25%。不同基因的内含子序列长度差异很大,AhTCP15的内含子长度仅为80 bp,而AhTCP9内含子序列长达791 bp。结合系统进化树的结果可以看出,千穗谷TCP基因的内含子数目在不同亚家族之间具有一定的特异性:ClassⅠ亚家族中,除AhTCP15外的其他成员均不含内含子,而ClassⅡ亚家族中将近一半的成员含有1个内含子和2个外显子(图3)。

图3 千穗谷TCP家族成员基因结构分析图
千穗谷TCP蛋白的保守基序分析显示,在进化树中同一分支的AhTCP蛋白通常具有相似的基序组成。除了TCP结构域所在的motif1外,TCP家族不同成员之间还存在多个保守的DNA区段。比如,除AhTCP7外,Class Ⅱ的CIN亚类中都存在Motif3,Class Ⅰ的PCF分支中的AhTCP蛋白都具有Motif2和Motif5;有的Motif只在个别成员中存在,比如Motif6和Motif10仅存在于AhTCP3和AhTCP8中。
2.4 染色体定位分析
如图4所示,千穗谷的16个TCP基因仅分布在Chr1,Chr2,Chr3,Chr4,Chr6和Chr11等6条染色体上,其他10条染色体无TCP基因分布。其中,Chr2染色体上共分布有5个TCP基因,且包含三个不同亚类;而Chr4和Chr6染色体上仅分布有1个TCP基因,均位于染色体端粒的位置。部分TCP基因在染色体上的分布呈现出成簇的现象,比如Chr1染色体上的AhTCP15,AhTCP16,相距仅0.17Mb;AhTCP1和AhTCP2两个基因,均分布在Chr2染色体端粒部分34.27~34.54 Mb的区域内。
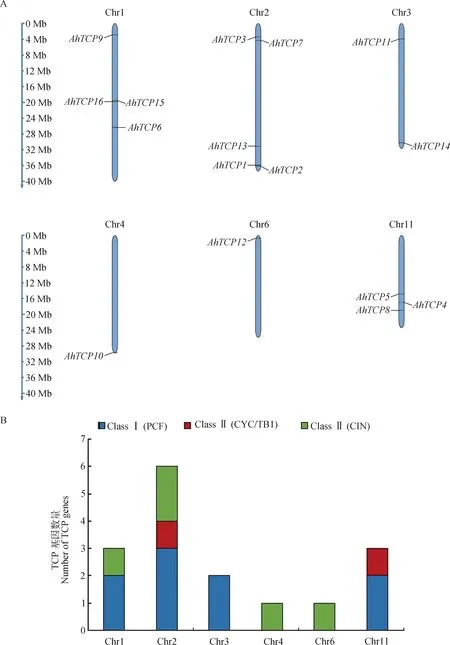
图4 千穗谷TCP家族成员染色体定位信息
2.5 顺式作用元件预测
对千穗谷TCP基因上游1 000 bp序列进行顺式作用元件预测,发现千穗谷TCP基因的启动子区域包含多个与植物生长发育和逆境响应相关的调控元件(图6)。比如,G-Box,Box 4,GT1-motif,MRE等光响应元件,在所有AhTCPs启动子区域均有发现;而响应无氧环境的ARE也存在于大部分的AhTCPs中。此外,部分AhTCP基因上游区域含有响应低温的元件LTR,比如AhTCP7,AhTCP13和AhTCP14等;部分基因包含响应干旱胁迫的顺式作用元件MBS,比如AhTCP5,AhTCP6和AhTCP10等。很多AhTCPs启动子含有响应激素的顺式作用元件,比如AhTCP5,AhTCP7,AhTCP15,AhTCP16中包含响应生长素的AuxRR-core或TGA-element元件,AhTCP4,AhTCP5,AhTCP6,AhTCP7中包含响应赤霉素的P-box或GARE-motif等元件。部分顺式作用元件仅在少数AhTCP基因中存在,例如响应昼夜节律的Circadian仅在AhTCP5中存在,参与胚乳特异性表达的GCN4-motif仅在AhTCP10和AhTCP15中存在。
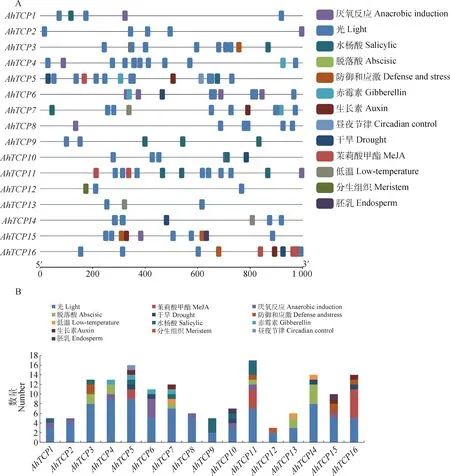
图5 千穗谷TCP基因启动子顺式作用元件预测
2.6 组织特异性表达
基因的组织特异性表达通常与其功能密切相关。根据Phytozome数据库中的信息可知,16个千穗谷TCP基因分别在根、茎、子叶、叶、花、未成熟种子、成熟种子和混合组织中检测到不同水平的表达量(图6)。不同亚家族的千穗谷TCP基因表达模式相似,比如PCF亚家族中的AhTCP2,AhTCP3,AhTCP5,AhTCP8,AhTCP14,AhTCP15,AhTCP16的表达模式被聚为一类;CIN亚家族中的AhTCP10和AhTCP12的表达模式被聚为一类且在大部分组织中未表达。其中,AhTCP4仅在花中表达;AhTCP13仅在根和茎中表达。AhTCPs在不同组织和器官中的表达模式不同,暗示不同成员在千穗谷的生长发育不同阶段具有不同的功能。
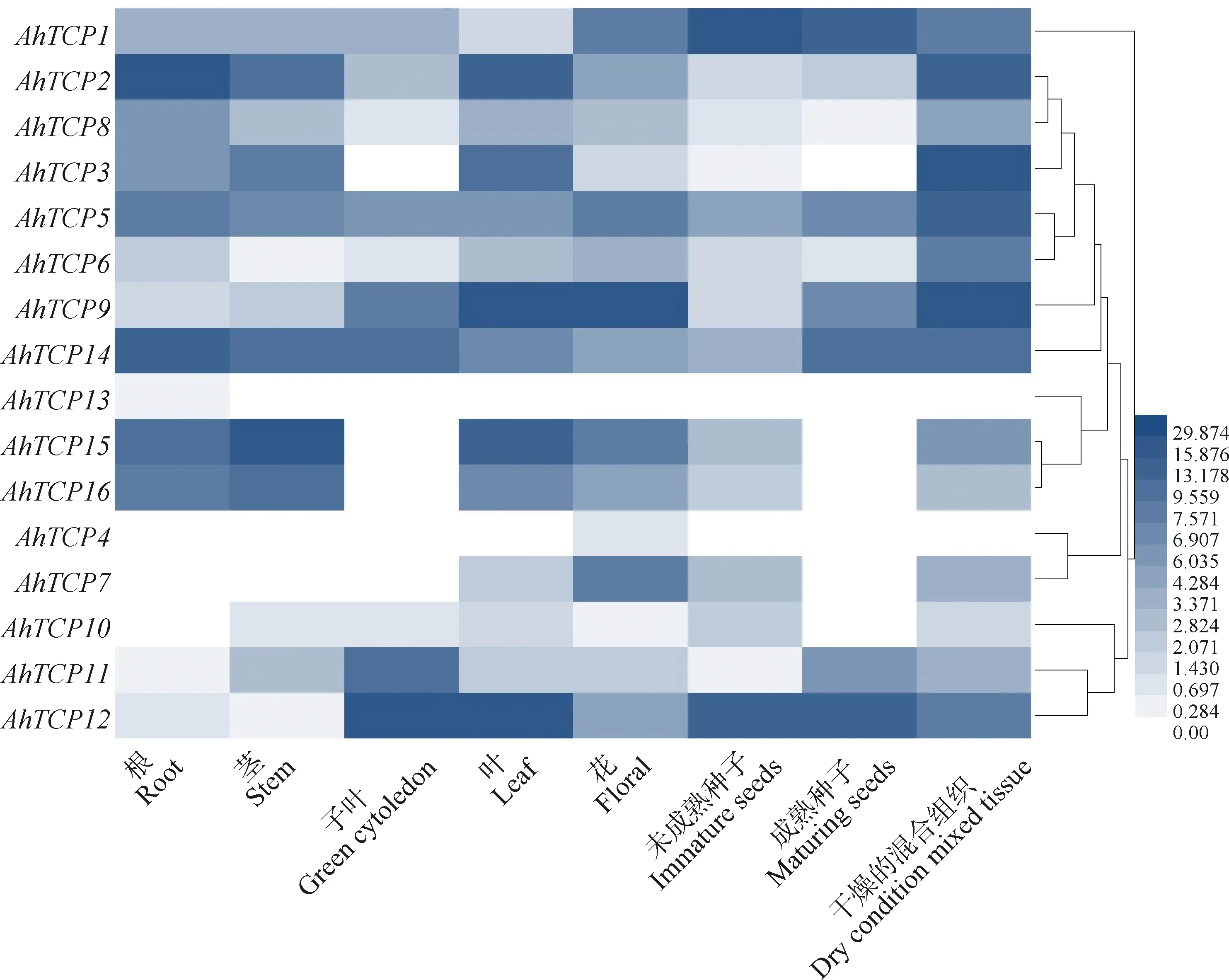
图6 不同组织千穗谷TCP基因表达量分析
2.7 千穗谷TCP基因的miRNA预测
以AhTCP基因的编码区为靶序列预测能够与其互作的miRNA,发现多达47个类型的miRNA与AhTCP基因存在结合位点。其中,AhTCP12编码区存在的结合位点最多,高达16个,AhTCP2存在的结合位点最少,只有1个(图7A)。大部分的miRNA只跟1个AhTCP基因互作,但部分miRNA可以同时靶向不同的AhTCPs。比如,miR159,miR5658和miR834在千穗谷TCP基因中存在4个靶基因,miR472,miR838和miR319存在3个靶基因,另有11个miRNA存在2个靶基因(图7B)。
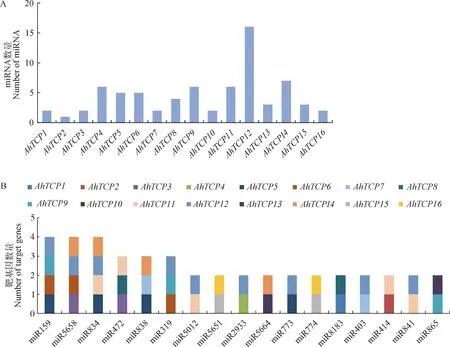
图7 AhTCP基因的互作miRNA预测
3 讨论
千穗谷TCP基因家族不同成员的基因长度、氨基酸大小、分子量和理论等电点差异较大,均编码碱性氨基酸且定位于细胞核;根据氨基酸序列的差异,TCP基因家族可分为Class Ⅰ和Class Ⅱ两大类,其中Class Ⅱ又分为CYC/TB1和CIN两个分支[16]。千穗谷TCP基因家族也同样包含三个亚类。序列比对发现,AhTCPs两个螺旋区的保守氨基酸残基中非极性氨基酸含量更多,例如丙氨酸(A)、亮氨酸(L)、异亮氨酸(I)、苯丙氨酸(F);环状区域则包含许多亲水性氨基酸残基,例如赖氨酸(K)、精氨酸(R)、丝氨酸(S)、天冬氨酸(D)等,这与前人的报道相似[41]。氨基酸序列对比和保守基序分析均显示不同的亚家族具有一定的保守性,例如在PCF,CYC/TB1,CIN亚类中均含有特定的保守的氨基酸(图1)、Class Ⅰ亚家族中均含有Motif2和Motif5。基因结构分析显示千穗谷TCP基因结构比较简单,仅AhTCP4,AhTCP9,AhTCP12和AhTCP15含有一个内含子,大多数成员均没有内含子。这与其他物种中的研究结果一致[41-42],表明TCP基因家族序列具有较强的保守性。染色体定位分析表明AhTCP基因仅定位在千穗谷的6条染色体上,基因在染色体上随机分布且存在成簇的现象,这可能和基因的复制有关。
生物信息学分析可以为预测家族成员功能提供多方面依据。很多研究表明,直系同源基因通常具有相似的功能[43]。Danisman研究发现,AtTCP9通过改变茉莉酸代谢从而促进根的伸长[44],本研究显示,千穗谷的AhTCP13基因和AtTCP9在系统进化树中聚在同一个分支,且AhTCP13主要在根和茎中表达,因此AhTCP13很可能也具有与AtTCP9相似的功能和调控机制,在千穗谷的根茎发育中起重要作用。Sun等人发现拟南芥CIN亚类的AtTCP4具有激活次级细胞壁生物合成和细胞凋亡的功能,在花器官发育过程中起关键作用[21]。千穗谷的AhTCP9与AtTCP4聚在同一个分支,遗传距离较近且组织特异性表达显示其在花中表达量较高,推测AhTCP9基因很可能参与了千穗谷花的发育。前人研究表明,莲藕(Nelumbonucifera)和豌豆(Pisumsativum)中CYC/TB1分支的TCP蛋白参与调控花的不对称性[45-46],同样属于CYC/TB1分支的AhTCP4仅在花中表达,推测其很可能参与了千穗谷花的形态建成。
转录因子通过与下游靶基因启动子上顺式作用元件的相互作用,在植物发育过程和非生物胁迫反应中发挥重要作用[47]。本研究对千穗谷TCP启动子顺式作用元件进行预测,发现其启动子区域含有大量与激素调控和胁迫反应有关的顺式作用元件,暗示千穗谷的TCP基因可能参与了多种发育事件和胁迫反应。不同的千穗谷TCP基因启动子区域的顺式作用元件类型不同,显示了家族基因间功能上的分化。比如,拟南芥中PCF分支的TCP14和TCP15通过促进幼嫩节间的细胞分裂来影响植株高度,还可调节叶片发育和花组织的细胞增殖[48],而千穗谷中与该基因同属PCF分支的AhTCP15和AhTCP16在茎中表达水平较高,且启动子中包含响应生长素的顺式作用元件,推测AhTCP15和AhTCP16基因可能通过响应生长素进而调控千穗谷茎的生长(图6)。此外,ClassⅠ亚家族的AhTCP3,AhTCP11,AhTCP15,AhTCP16在其启动子区域包含多个与防御和应激相关的顺式作用元件(TC-rich repeats),暗示其可能参与了植物的防御机制,这与拟南芥ClassⅠ亚家族中的AtTCP14,AtTCP15,AtTCP19等基因的功能相似[48]。
对AhTCP基因的编码区进行miRNA作用位点预测,结果显示有47个miRNA在AhTCP基因上存在结合位点,其中,miR319和miR5658的预测准确度最高。多个物种中的研究均表明,miR319主要靶向作用于TCP基因,且主要为ClassⅡ亚家族中CIN分支成员[32,49]。比如,水稻中的Osa-miR319bc通过调控OsPCF6和OsTCP21的表达调节水稻对冷的应激反应[50];鸡爪槭(Acerpalmatum)中的miR319通过靶向ApTCP2影响植株开花、叶片形态和衰老[20]。本研究中,AhTCP6,AhTCP9和AhTCP12基因中均含有miR319的结合位点,且三者都属于ClassⅡ亚家族中CIN分支成员,这与其他物种类似[51]。进一步分析显示,AhTCP12与ath-miR319在互补配对区有4个碱基的差异,AhTCP6和AhTCP9与ath-miR319在互补配对区仅有2个碱基的差异(图8)。

图8 miR319靶向AhTCP基因的结合位点
4 结论
根据TCP结构域特点,千穗谷的16个TCP基因分为PCF,CYC/TB1和CIN三个分支,与其他物种的亚家族分类相似。染色体定位结果显示,AhTCPs不均匀的分布在6条染色体上,部分基因呈簇状分布。顺式作用元件预测和组织特异性表达分析均显示千穗谷TCP基因家族不同成员存在功能上的分化:不同AhTCPs的启动子所包含的顺式作用元件存在较大差异,暗示其响应不同的植物激素或者环境信号;16个AhTCPs基因在不同组织和器官中的表达模式不同,系统进化树中同一分支的成员表达模式相似,不同分支的成员存在较大差异。miRNA预测结果显示,miR319在千穗谷TCP家族中的靶基因也主要为CIN亚类成员。
