外源性神经肽CART对大鼠海马神经元氧化应激的抵抗作用及机制研究
2022-04-24杨成迎林海烂黄子晴王乃秀
杨成迎,林海烂,黄子晴,王乃秀,汪 锴,甘 玲,2*
(1.西南大学动物医学院,重庆 402460;2.重庆市兽医工程研究中心,重庆 402460)
在动物生长过程中,由于环境、营养及病原感染等因素引起的机体氧化与抗氧化系统的失衡,易导致机体出现氧化应激,从而损伤组织细胞[1],极大地增加了动物疾病的发生率。哺乳动物中枢神经系统的氧化代谢非常活跃,是遭受氧化应激损伤较严重的部位,而脑海马作为脑部结构重要的区域之一,不仅是应激损伤的重要靶点,同时,也通过表达大量的神经肽参与对应激损伤的调控[2-3]。研究表明,当仔猪发生氧化应激时,海马组织中神经肽可卡因-苯丙胺调节转录因子(cocaine-and amphetamine-regulated transcript, CART)表达显著下调[4]。
研究发现,CART对中风、脑缺血等神经元损伤有保护作用[5-7]。CART可通过促进脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)的表达来增加神经元的存活、增殖、迁移和分化[8-10]。同时,CART通过上调BDNF基因的表达,抑制了大鼠神经元内质网应激和神经元的凋亡,但trkB免疫抗体可抑制该作用[11]。trkB是定位于线粒体内膜上的BDNF受体[12]。由此推测,氧化应激引起仔猪脑海马中CART的表达下调,可能会抑制BDNF/trkB信号通路,从而引起神经元的凋亡,而补充外源性的神经肽CART则可能通过上调BDNF/trkB通路抵抗氧化应激引起的海马神经元凋亡。为了验证该假设,本研究拟采用过氧化氢诱导原代培养的海马神经元,以构建海马神经元氧化应激模型,并添加不同浓度的外源性CART处理海马神经元,通过检测神经元活力和ATP含量,筛选抵抗海马神经元氧化损伤的最佳CART处理浓度。随后采用JC-1染色法检测神经元线粒体的膜电位,荧光定量PCR和Western blot技术检测神经元凋亡相关因子和trkB基因的表达变化,并分析其相关性,以探索外源性神经肽抵抗海马神经元氧化应激的作用效果及分子机制,为神经肽类动物应激调节剂的研发奠定基础。
1 材料与方法
1.1 实验动物
出生24 h以内的SD大鼠(SPF级)购自重庆医科大学实验动物中心。
1.2 主要试剂
bcl-2一抗、bax一抗、caspase-3一抗均购自Proteintech公司(中国);trkB Antibody(一抗)购自于美国R&D SYSTEMS 公司;Goat Anti-Rabbit IgG(H+L)(二抗)购自Affinity公司;FITC Affinipure Goat Anti-mouse IgG(H+L)购自美国Earthox 公司(中国);DAPI染色液购自北京四正柏公司;MTT试剂盒购自上海生工公司(中国);ATP试剂盒、JC-1试剂盒购自碧云天公司(中国);冰乙酸购自重庆川东化工有限公司。CART肽由合肥博肽生物科技有限公司合成。
1.3 试验方法
1.3.1 原代大鼠海马神经元的培养 剥取24 h以内的SD新生大鼠脑组织,分离其海马并浸润在100 mL·L-1FBS-DMEM-F12 培养基中剪碎、反复多次吹打及消化后,采用筛网(200目孔径)过滤后,用适量 DMEM/F12(含100 mL·L-1FBS 胎牛血清)将单细胞悬液稀释后,均匀接种在细胞瓶或 6 孔板中,进行海马神经元原代培养,待大多数细胞开始贴壁后(2~4 h),采用含 20 g·L-1B27、10 g·L-1L-谷氨酰胺及 10 mL·L-1双抗(100 U·mL-1青霉素、100 g·mL-1链霉素)的 neurobasal TM-a培养基进行维持培养,每天在倒置显微镜下观察细胞变化,根据细胞生长情况,每隔2 d更换1/3~1/2的培养液。神经元培养2 d 后加入阿糖胞苷(终浓度: 2 μg·mL-1)抑制胶质细胞增殖。采用 MAP2-GFAP 免疫荧光法鉴定神经元的纯度至少大于95%。
1.3.2 试验分组和处理 在海马神经元培养7 d后,将神经元分为5组,即对照组(Control)、过氧化氢组(H2O2)、H2O2+10 nmol·L-1CART组、H2O2+100 nmol·L-1CART组、H2O2+1 000 nmol·L-1CART组。每组3个重复。对照组不做处理,过氧化氢组用200 μmol·mL-1H2O2处理神经元,孵育24 h。各CART组在H2O2处理的基础上分别添加终浓度为10、100、1 000 nmol·L-1CART的培养液,孵育4 h。
1.3.3 神经元活率、ATP含量及线粒体膜电位(ΔΨm)的检测 将成功培养并处理好的神经元,分别严格按照试剂盒说明书进行样品收集并进行下一步检测。采用MTT法检测神经元活率:将神经元置于96孔板上培养,按试验设计处理神经元后,每孔加入10 μmol·L-1的MTT培养基,于37 ℃孵育24 h。移除培养基,每孔加入100 μL Farmazan solubilization solution 37 ℃孵育10 min,于紫外可见分光光度计570 nm处检测吸光度并计算神经元活率。
采用化学发光仪检测ATP含量:将处理好的样品用ATP裂解液裂解神经元,充分裂解后进行离心5 min(12 000 r·min-1,4 ℃),收集上清液。将200 μL ATP检测工作液加入检测孔中消耗孔内ATP,3~5 min后每孔加入20 μL待测样品或稀释过的ATP标准液,充分混匀并常温孵育2 min。
采用化学发光仪测量相对光单位(RLU)的亮度:用荧光显微镜观察JC-1荧光变化以确定ΔΨm:处理好的神经元中加入JC-1染色工作,培养箱37 ℃ 孵育20 min。孵育结束后,用JC-1染色缓冲液洗涤2次,并加入2 mL细胞培养液,在荧光显微镜下观察线粒体膜电位的变化。
1.3.4 荧光定量PCR技术检测凋亡通路相关基因的转录表达水平 采用 TRIzol法提取海马神经元的总RNA,并反转录制备cDNA。以GAPDH作为内参基因,采用实时荧光定量 PCR(RT-qPCR)技术检测神经元凋亡通路相关基因的转录表达水平,引物序列见表1。Real-Time PCR 的反应体系(20 μL)如下:SYBR Premix Ex Taq 10 μL;ROX Reference Dye 0.4 μL;PCR Forward Primer 0.4 μL;PCR Reverse Primer 0.4 μL;DNA 模板2 μL;ddH2O 6.8 μL。反应程序(两步法)如下:95 ℃,30 s;95 ℃,5 s,60 ℃,30 s,40个循环;95 ℃,15 s,60 ℃,1 min,95 ℃,15 s。采用 2-ΔΔCt相对定量法计算基因相对表达量。

表1 引物序列
1.3.5 Western blot技术检测线粒体凋亡通路相关基因的翻译表达水平 按照1 mL RIRP加入10 μL混合型蛋白抑制剂配制成裂解液,混匀后裂解神经元,在冰上充分研磨并静置30 min,吸取混合液进行离心30 min(12 000 r·min-1,4 ℃),吸取上清置于1.5 mL EP管,-80 ℃冻存备用;配制BCA工作液,在OD562 nm下检测吸光度值,制作标准曲线并计算各个样品的浓度;加入Loading Buffer于每个待测蛋白样品中,混合均匀后插在煮样泡沫板上,于98 ℃热处理10 min 使蛋白变性;后进行SDS-PAGE电泳(80 V,20 min,120 V,1 h),然后转膜至PVDF膜(200 mA,90 min)。采用5%脱脂奶粉室温慢摇封闭1~2 h后,一抗室温慢摇2 h或4 ℃孵育过夜,TBST漂洗3次,每次10 min。在二抗室温慢摇孵育1 h后,TBST漂洗3次,每次10 min,最后采用ECL发光液显影。
1.3.6 数据分析 试验数据采用SPSS 20.0(IBM;New York City,NY,USA)软件进行统计及显著性及相关性分析,单因素方差分析检验进行组间差异比较分析。结果均以“平均值±标准误(Mean ± SEM)”表示,P<0.05为差异显著,P<0.01为差异极显著。
2 结 果
2.1 CART对氧化应激海马神经元活率和ATP含量的影响
如图1,与对照组相比,H2O2组海马神经元活率和ATP含量均极显著下降(P<0.01);与H2O2组相比,添加10 nmol·L-1CART可以显著提高海马神经元活率和ATP含量(P<0.05),添加100和1 000 nmol·L-1CART组中神经元活率和ATP含量均显著高于H2O2组(P<0.05或P<0.01),且显著高于10 nmol·L-1CART添加组(P<0.05)。该试验结果表明,与H2O2组比较,添加10 nmol·L-1CART能显著抵抗氧化应激造成的海马神经元损伤,因此后续试验添加10 nmol·L-1CART进行处理。

与Control比较,*P<0.05,**P<0.01;与H2O2比较,# P<0.05,## P<0.01;与H2O2+10 nmol·L-1 CART组比较,& P<0.05,&& P<0.01。下同
2.2 CART对氧化应激海马神经元线粒体膜电位的影响
与对照相比,H2O2组中线粒体膜电位显著降低(P<0.05),在添加CART之后,线粒体膜电位显著高于H2O2组(P<0.05,图2),表明氧化应激引发神经元线粒体损伤,从而降低线粒体膜电位,而CART能稳定线粒体膜电位,从而防止线粒体损伤引起神经元凋亡。
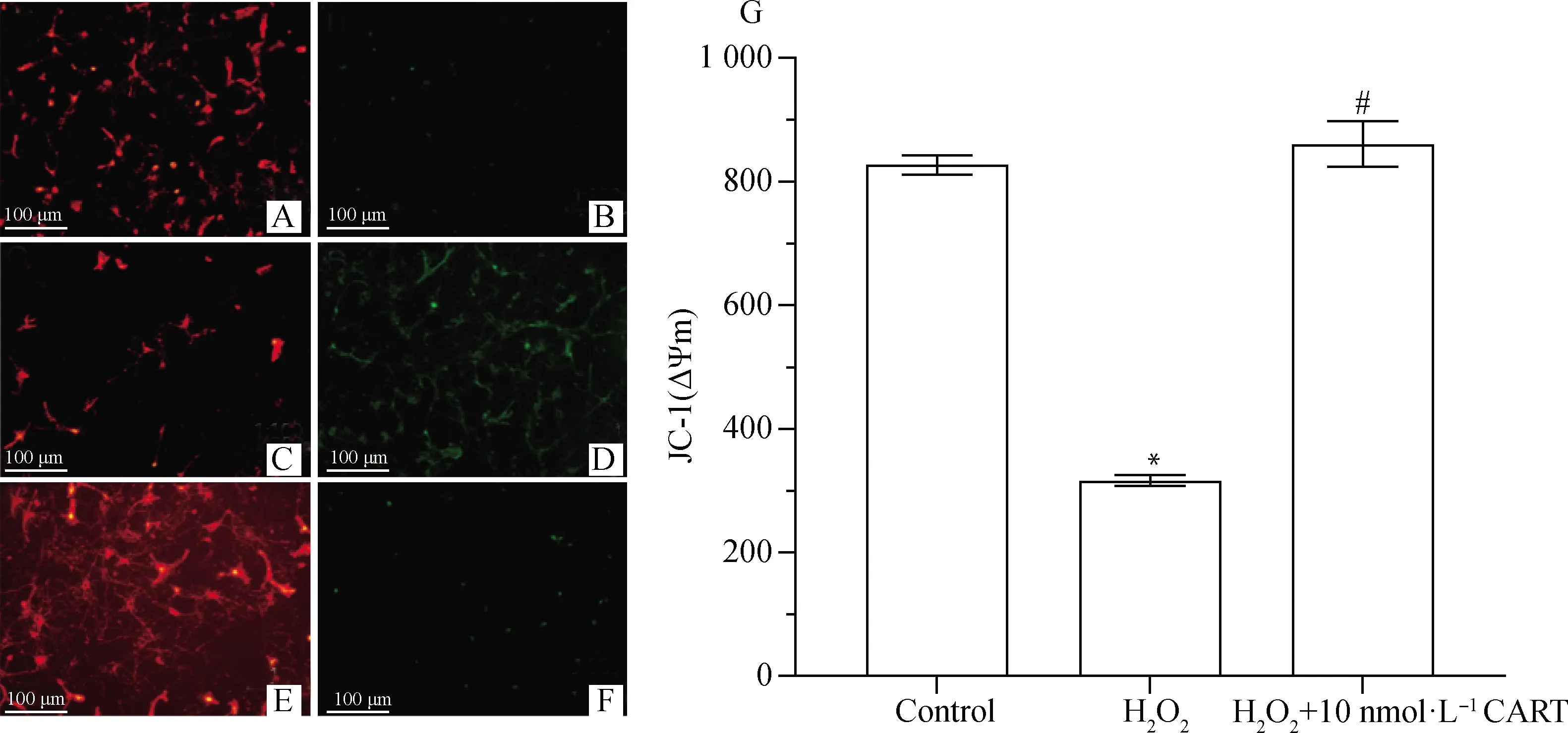
A、B.对照组;C、D.H2O2组;E、F.H2O2+10nmol·L-1 CART组;G.线粒体膜电位柱形图
2.3 CART对氧化应激海马神经元线粒体凋亡基因表达水平的影响
为进一步考察线粒体凋亡信号通路在CART抵抗氧化应激损伤中的介导作用,本研究采用荧光定量PCR技术检测海马神经元中线粒体凋亡基因mRNA水平的变化。结果显示,与对照组相比,H2O2组中抑凋亡基因bcl-2的mRNA水平极显著降低(P<0.01),促凋亡基因bax和caspase-3的mRNA 水平均极显著增加(P<0.01);与H2O2组比较,添加CART组中bax基因的mRNA水平显著降低(P<0.05),caspase-3基因的mRNA水平极显著降低(P<0.01),bcl-2基因的mRNA水平极显著高于H2O2组(P<0.01)(图3A)。采用Western blot技术进一步考察CART对氧化应激海马神经元凋亡基因蛋白质水平的影响。结果显示,与对照组相比,H2O2组中caspase-3和bax的蛋白质水平显著升高(P<0.05),bcl-2的蛋白质水平显著降低(P<0.05);与H2O2组比较,CART添加组中caspase-3和bax的蛋白质水平显著降低(P<0.05),bcl-2的蛋白质水平显著升高(P<0.05)(图3B、3C、3D、3E),与基因转录表达水平一致,表明CART可以抑制H2O2诱发的神经元凋亡。

A.荧光定量PCR技术检测结果;B、C、D、E.Western blot技术检测结果
2.4 CART对氧化应激海马神经元trkB表达水平的影响
为考察BDNF/trkB信号通路是否参与了CART抗海马神经元氧化应激损伤的作用,本研究采用荧光定量PCR和Western blot技术检测了海马神经元trkB的mRNA和蛋白质水平。结果显示,与对照组相比,H2O2组中trkB的mRNA水平显著降低(P<0.05),H2O2+CART组中trkB的mRNA水平显著高于H2O2组(P<0.05,图4A)。对trkB的蛋白水平的检测结果显示,与对照组相比,H2O2组中trkB的蛋白表达量显著降低(P<0.05);与H2O2组比较,H2O2+CART组中trkB蛋白水平显著升高(P<0.05),与mRNA表达水平变化一致。由此表明,CART可能通过trkB基因的表达调控凋亡蛋白基因的表达,从而抑制细胞凋亡,缓解氧化应激。

A.荧光定量PCR技术检测结果;B、C.Western blot技术检测结果
2.5 海马神经元trkB与凋亡相关蛋白、基因表达水平相关性分析
为了更好地显示BNDF/trkB信号通路与凋亡信号通路之间的关系,本研究考察了trkB与凋亡信号通路相关的蛋白和基因表达水平之间的相关性。结果如图5所示,在过氧化氢组,trkB与bax的mRNA及蛋白质水平呈显著负相关(Pearson系数=-0.999 7;P=0.015 5),与bcl-2基因的mRNA 水平呈显著正相关(Pearson系数=0.996 9;P=0.049 8),并与caspase-3的蛋白质水平呈显著负相关(Pearson系数=-0.999 1,P=0.026 3);在H2O2+CART组,trkB与bax的蛋白质水平呈显著负相关(Pearson系数=-0.998 1;P=0.039 5),与trkB与bcl-2的蛋白质水平呈正相关的趋势(Pearson系数=0.995 1;P=0.063 3)。以上结果表明,trkB与凋亡相关蛋白和基因的表达呈显著相关性,进一步提示BNDF/trkB信号通路在CART调节凋亡信号路径抵抗氧化应激损伤中的介导作用。

A~C.在H2O2组,trkB与caspase-3、bax及bcl-2在mRNA水平的相关性; D~F.在H2O2+CART组,trkB与caspase-3、bax及bcl-2在mRNA水平的相关性; G~I.在H2O2组,trkB与caspase-3、bax及bcl-2在蛋白质水平的相关性; J~L.在H2O2+CART组,trkB与caspase-3、bax及bcl-2在蛋白质水平的相关性
3 讨 论
氧化应激是引起机体细胞凋亡的重要原因,而线粒体损伤是其中的关键因素。线粒体是细胞的能量加工厂,其主要作用是为细胞的各种生命活动提供能量ATP[13]。线粒体膜是否完整对细胞正常发挥功能至关重要[14]。膜损伤后功能失调的线粒体产生ATP的效率较低,但产生ROS的效率较高[15-18]。由此可见,ATP是反映机体活细胞新陈代谢的一个重要指标,通过测定培养细胞ATP的含量可反映出细胞的活力和线粒体的完整性[19-20]。因此,本研究中过氧化氢诱导神经元后降低的ATP含量及神经元活率证实神经元的线粒体受到了损伤。众多研究表明,CART肽在抵抗脑组织的氧化损伤中发挥了重要的作用[21-23]。体外的研究也显示,CART肽可降低缺氧和缺糖神经元中的ROS水平,增加神经元抗氧化性和线粒体活性[24]。与此相似,本研究添加不同剂量外源性神经肽CART显著提高了氧化应激大鼠海马神经元的活力及ATP水平,暗示神经肽CART可能通过抵抗氧化应激对神经元线粒体造成的氧化损伤来发挥作用。
线粒体膜电位由线粒体内膜和外膜之间的不同离子电化学梯度产生。研究发现,当细胞出现凋亡的第一个信号便是线粒体膜电位值(MMP)显著降低[25-26]。JC-1作为一种常用的检测MMP(ΔΨm)的理想荧光探针,可以检测组织或细胞的线粒体膜电位。当线粒体膜电位较高时,JC-1呈聚合物状态,聚集在线粒体基质中,可以产生红色荧光;而在线粒体膜电位较低时,JC-1呈单体状态,便不在线粒体基质中聚集,而发出绿色荧光。通过荧光颜色的转变便可检测线粒体膜电位的变化。因此,本研究中过氧化氢诱导的神经元MMP显著降低,表明线粒体膜的通透性变小,进一步证实过氧化氢诱导使神经元的线粒体遭到损伤。Wang等[27]的研究结果显示,与对照组相比,氧化应激状态下的大鼠海马神经元MMP显著下降,而他克莫司可以显著抑制MMP的下降,从而保护神经元免于凋亡。1-甲基-4-苯基吡啶(1-methyl-4-phenylpyridinium,MPP +)在诱导SH-SY5Y细胞凋亡的同时,显著降低了MMP,而经过磷酸二酯酶4治疗可阻碍MMP的下降,同时抑制细胞凋亡[28],由此表明,MMP与神经元凋亡之间呈正相关。研究发现,位于线粒体外膜的调节因子bcl-2家族蛋白、caspase系列蛋白、细胞色素C与细胞凋亡有密切的关系[29]。氧化应激将伴随线粒体凋亡途径关键蛋白caspase-3、bax及抗凋亡蛋白bcl-2基因的表达变化[30-32]。番茄红素(lycopene,LYC)能使细胞中bax和caspase-3基因的表达水平显著降低,而bcl-2基因的表达水平显著升高[33]。因此,为了进一步考察神经肽CART抵抗神经元线粒体氧化损伤的分子机制,本研究考察了凋亡相关基因的表达变化。结果显示,CART在显著降低氧化应激海马神经元bax和caspase-3基因表达水平的同时,增加了bcl-2基因的表达,表明CART能显著抑制神经元凋亡,从而缓解海马神经元的氧化应激。
有文献表明,trkB是脑源性营养因子(BDNF)的受体,位于线粒体内膜[12,34]。trkB能通过调节细胞线粒体内钙离子的释放,影响线粒体活性[35],并且介导apelin-13、N-乙酰基5-羟色胺(NAS)及丙咪嗪等多种药物或抗氧化物质发挥抗凋亡的调节作用[36-38]。因此,为了考察trkB是否也介导了CART抵抗神经元氧化损伤的作用过程,本研究检测了trkB基因的表达变化,并与凋亡相关基因的表达变化做进一步相关性分析。结果发现,trkB基因与凋亡蛋白基因的表达具有显著相关性,这相似于前人的研究报道NAS通过调节BNDF/trkB信号通路,抑制了bax和caspase-3基因的表达,并增强了bcl-2基因的表达,从而防止谷氨酸诱导HT-22细胞凋亡[38],因此,进一步提示了trkB在CART调节凋亡信号路径抵抗氧化应激损伤中的介导作用。
4 结 论
在本研究中,添加CART能显著提高氧化应激海马神经元trkB基因的mRNA和蛋白质水平,且trkB与凋亡/抗凋亡相关基因的表达显著相关,提示CART可能通过激活BDNF/trkB信号通路来改变膜电位,防止海马神经元凋亡,从而缓减氧化应激。
致谢:本研究感谢西南大学何宁佳和罗义维对qRT-PCR测定的平台支持
