海洋酸化与升温对浮游植物种群的影响研究综述
2022-04-21冯媛媛王建才
冯媛媛,王建才,蔡 婷
(天津科技大学海洋与环境学院,天津 300457)
浮游植物是海洋生态系统汇总物质循环和能量流动的基础,贡献了地球上约50%的初级生产力,在全球氧生产、能量传递过程中发挥重要作用[1-2].近年来,在全球气候变化和人类活动的共同作用下,海洋浮游植物赖以生存的海洋理化环境发生了显著的改变,浮游植物对这些环境变化也将产生系列生理生态学响应,并将进一步对海洋上层食物网、海洋生态系统结构和服务功能以及海洋碳汇产生深远的影响.因此,研究全球气候变化的生理生态学效应已成为海洋学中的热点方向[3–5].
1 海洋酸化和海洋升温
自工业革命以来,随着化石燃料的大量使用,大气二氧化碳(CO2)浓度平均值持续快速上升[6].海洋是地球表面最大的碳库,吸收了人类排放CO2总量的三分之一,其吸收速率每天可达2500多万吨[7].然而,随着海洋吸收CO2量的增加,海水pH也随之降低,引起海洋酸化[8-9].与19世纪70年代前水平对比,上层海洋中H+浓度已增加了32%,pH随之下降了0.1.根据IPCC预测模型推测,到21世纪末表层海水pH将进一步下降0.3~0.4[10].
海洋酸化所引起的海水化学(碳酸盐系统及物质形态)、物理(声波吸收)及生物过程的变化[11–12],影响到多种海洋生物的生理过程,并将影响海洋生态系统的平衡.近海海洋环境是陆海交界地带,同时也是水产养殖的重要区域,更易受到人类活动影响.近岸海域复杂的生态结构决定了其对环境变化的特殊敏感性[13],相较其他海洋区域,近岸海域最先容易受到海洋酸化的影响,其生态环境健康也易受到全球气候变化的影响和破坏[14].
二氧化碳是主要的温室气体,其过度排放导致了温室效应,进而引起表层海水温度也随之升高[15].自20世纪50年代以来,全球水深700m以下的海水温度每10年平均上升约0.02℃,全球表层海水温度则每10年升温0.07℃[16].据模型预测,到21世纪末全球表层海水平均温度将会持续升高1~4℃[17].近百年间,中国近海海域水体增温较为明显,尤以20世纪90年代以来升高最快,模型预测升温速度为每百年升高0.8~2℃[18-19].温度是影响生物代谢途径中酶活性的重要环境因子,对生物有机体的生长和新陈代谢等过程起着至关重要的作用[20–22].海洋升温可引起多种生物生态位、优势浮游生物种群的季节性变迁、浮游生物群落结构以及全球海洋生产力格局等发生变化[23–24].
2 海洋酸化对浮游植物的生理生态学效应
2.1 海洋酸化对不同浮游植物种群的生理学效应
海洋酸化引起海水碳酸盐体系发生显著变化,海水中的总溶解无机碳(DIC)和浓度升高.无机碳是浮游植物光合作用的主要碳源,浮游植物对CO2浓度变化的响应等相关研究在过去十几年受到科学界的重视,尤其是针对海洋酸化对海洋浮游植物生理生态学的影响进行大量研究[25-27].CO2浓度的变化不仅可以影响浮游植物的生长速率、光合固碳速率,也影响海洋初级生产力.海水中CO2浓度的增加,除了能给浮游植物细胞提供更多溶解无机碳外,还能有效防止胞内CO2逸出,抑制光呼吸,下调其无机碳浓缩机制,从而节约能量[28]供给于细胞生长和光合作用,因此在某种程度上促进浮游植物生长和光合固碳作用[29–30].
CO2浓度升高对光合固碳的促进作用在硅藻、甲藻、颗石藻中均有发现[31],且不同种类的浮游植物对海洋酸化的响应具有明显的种间差异性.尽管硅藻细胞普遍具有较为高效的碳浓缩机制(carbon concentration mechanism,CCM),但研究发现CO2浓度升高可以降低硅藻细胞中生物硅的含量[32],改变产毒硅藻毒素的生产[33].不同粒径大小[34]、近岸与大洋[35]、羽纹纲与中心纲硅藻物种[36]应对CO2浓度变化的生理响应也存在明显的种间差异.高CO2浓度可促进小皮舟形藻(Navicula pelliculosa)[36]的生长,而抑制直舟形藻(Navicula directa)[37]和威氏海链藻(Thalassiosira weissflogii)[38]的生长.有相关研究表明海洋酸化对海洋原甲藻(Prorocentrum micans)、亚历山大藻(Alexandrium sp.)的生长无明显影响[39],而对东海原甲藻(Prorocentrum donghaiense)和米氏凯伦藻(Karenia mikimotoi)的生长起到抑制作用[40].研究也发现CO2浓度升高明显促进了颗石藻优势物种赫氏艾密里藻(Emiliania huxleyi)和大洋桥石藻(Geophyracapsa oceanica)的生长[41–42],而对高度钙化的海洋颗石藻(Coccolithus pelagicus)的生长影响并不显著[43].
海洋酸化对浮游植物光合作用的影响也会受控于其他环境因子的影响.例如,在较低光照强度下中肋骨条藻光合能力随CO2浓度升高而增强[44],而在高光照强度条件下中肋骨条藻和三角褐指藻光合速率则随CO2浓度升高而下降[45].CO2浓度升高通常会降低钙化浮游植物颗石藻的钙化速率[46],但存在着种间及株间特异性[47–48],该响应同时也受到碳酸盐体系中不同无机碳组分的浓度[49]以及其他环境因子如营养盐水平、光照和温度条件的影响[50].
2.2 海洋酸化对浮游植物种群组成及演替的影响
目前对海洋酸化的生理生态学效应研究已经由对单一物种的响应机制上升到种群、群落甚至于生态系统角度.在不同CO2浓度下对自然群落进行模拟培养实验,测定其种群组成及演替变化和相关的生物地球化学参数,并由此判断其对上层食物网及真光层碳输出的影响.Tortell等[51]研究表明不同CO2浓度(0.015%和0.075%)对赤道太平洋海域浮游植物自然种群演替有显著影响,其中水体中优势度较高的硅藻相对丰度在低CO2浓度条件下降低了50%,而棕囊藻丰度升高了60%.王为民等[52]研究发现海洋酸化显著促进了黄海近岸海域叶绿素浓度(Chl-a)和浮游植物总丰度的上升,同时,浮游植物群落多样性在高CO2浓度条件下出现明显下降.在南极罗斯海进行的现场模拟培养实验则表明,CO2浓度的升高显著促进了浮游植物的生物量,并使容易成链的中心纲硅藻成为群落中的优势物种[53].在高CO2浓度条件下,秘鲁沿岸和北大西洋海域硅藻占比明显上升[51–54],而鄂霍次克海和白令海硅藻所占比例显著减少、微型浮游植物(nanophytoplankton)占比上升[55–56].近年来科学家开展的中尺度围隔实验进一步为我们在浮游植物种群及食物链的层面提供了认知,例如在德国基尔近岸区域开展的系列中尺度围隔实验KMSMOS[57].Schulz等[58]通过围隔实验发现,在CO2浓度升高的条件下,颗石藻优势种赫氏艾密里藻的丰度在整个培养阶段明显降低,而个体较小的聚球藻(Synechococcus)和超微型真核浮游植物的生长则受益于较高的CO2水平.总体上,自然浮游植物群落对海洋酸化的响应表现出了复杂的区域性.在过去的10年间,针对浮游植物自然群落的相应研究逐渐增多,不同研究的实验设计存在着较大的差异,如采样区域、CO2浓度、温度及营养盐添加情况等(其结果汇总见表1).
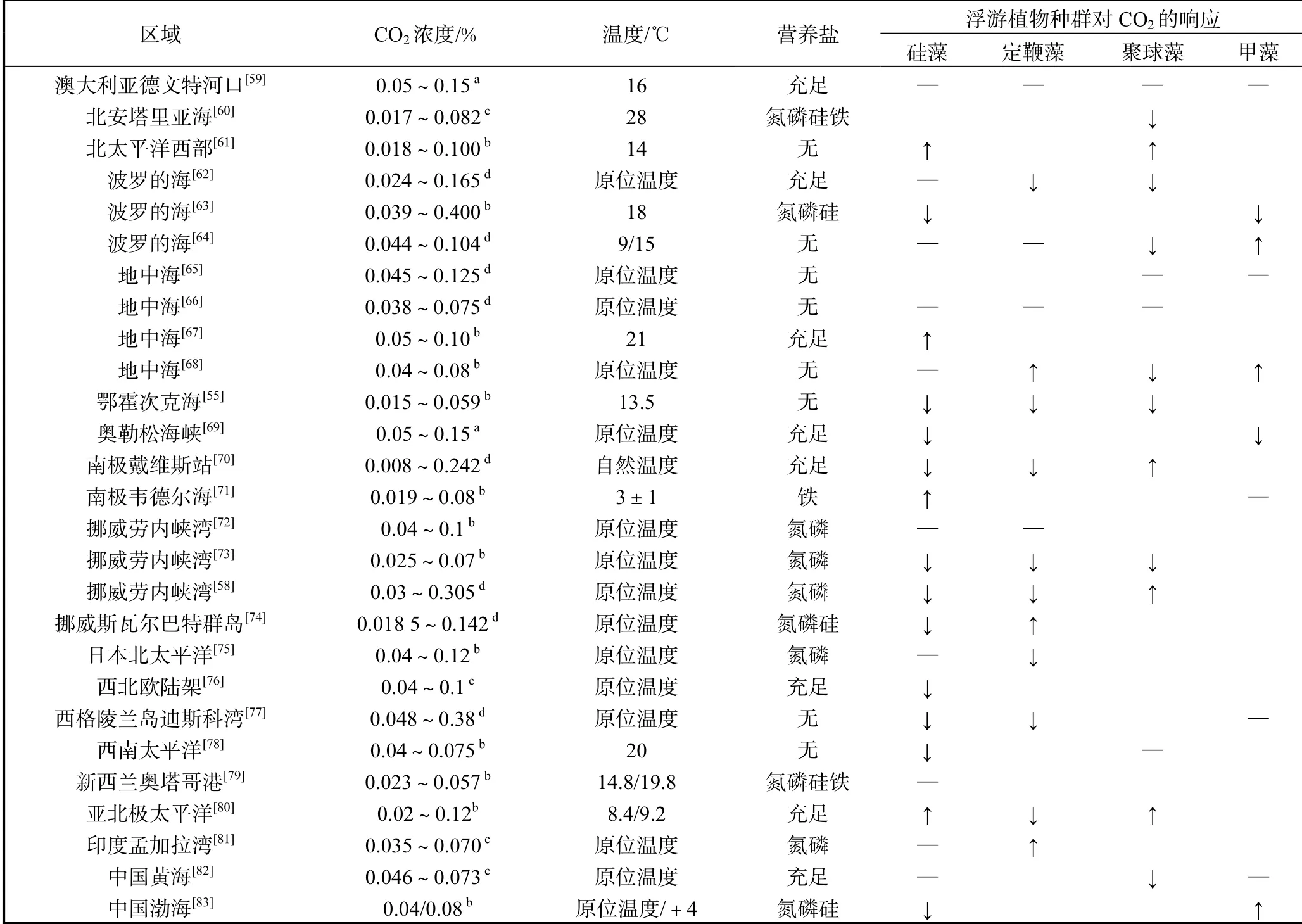
表1 近年来对自然浮游植物群落二氧化碳模拟培养实验结果汇总Tab. 1 Summary of published results from CO2 manipulation experiments on natural phytoplankton community in recent years
3 海洋升温对浮游植物的生理生态学效应
3.1 海洋升温对浮游植物不同物种的生理学效应
海水温度是海洋水文最重要的因子之一[84].海水温度对浮游植物的生命活动有重要的影响,一方面通过控制光合作用或呼吸作用强度直接影响藻类的生长繁殖;另一方面,可通过控制水体中各类营养物质的溶解度、离解度或分解率等理化过程间接影响藻类的生长[85].任何生物的生长和繁殖都离不开适宜的温度,大多数浮游植物生长和繁殖的适宜温度在20~30℃[86].
在一定温度范围内,升温通常可以导致浮游植物细胞生长和同化速率显著升高,影响藻类基因型和表型变异以及胞内化学转化和营养盐输运等过程[85].藻类的光合作用对温度变化尤为敏感,极端温度也会导致藻类光合中心细胞器受损[87].藻类细胞通常可以通过调节其核心生理过程和蛋白质合成来应对温度变化的压力[88].在适温条件下,升温对浮游植物生长的促进作用也存在种间差异.研究表明小球藻(Chlorella)和铜绿微囊藻(Microcystisaeruginosa)的生长受温度影响较大,随着温度的上升,其分裂速率显示出快速的升高,而变异直链藻(Melosira varians)生长速率的增长较慢[89].浮游植物不同物种对温度的生理响应也同样会受到其他环境因子的调节,例如在高的光照强度下,三角褐指藻的生长随着温度的升高而降低,而威氏海链藻的生长却受到了升温的促进,但该趋势在低的光照强度下则不明显[90].
3.2 海洋升温对浮游植物种群组成及演替的影响
浮游植物对于温度适应能力的差异将会导致浮游植物群落发生改变.由于不同浮游植物物种的最优生长温度存在差异[91],因此某些浮游植物物种将会更受益于海水升温;同时,另一部分将失去竞争优势,因而引起物种组成变化和群落演替[92].温度变化也可改变自然浮游植物物种的粒径大小[93],通常升温更有利于小粒径的初级生产者[94],从而影响海洋初级生产力、碳沉降及碳输出等重要过程.现场受控培养实验表明,随着温度升高,白令海浮游植物种群由硅藻向着微型浮游植物占优的趋势发展[95],而北大西洋春季水华浮游植物群落中的颗石藻相对占比则在升温4℃的条件下显著升高[35].近年来的相关模型研究也表明,海水温度的持续升高也会引发浮游植物物种向极地的迁移,并降低热带海域的浮游植物的物种多样性[96].赵瀛[97]通过研究升温对琼氏圆筛藻(Coscinodiscus jonesianus)生物量的影响发现,水温升高范围在0.2~0.79℃,能促进琼氏圆筛藻生物量的增加;温度升高在0.8~2.0℃范围内,则能抑制琼氏圆筛藻的生长繁殖过程.李婷等[98]在象山港国华电厂附近海域的研究发现:温排水对于浮游植物种类的影响因季节差异而具有明显区别.春季浮游植物的种类最少、多样性最低,秋季浮游植物种类和多样性最高,温排水对于夏季浮游植物群落结构也存在影响,能促进角毛藻更倾向于聚集到排水口邻近海域,导致了高温期角毛藻赤潮的概率增加.模型研究表明,全球变暖导致硅藻在北大西洋、南太平洋的丰度下降60%,在全球范围内下降10%[99],而海洋中原绿球藻(Prochlorococcus)和聚球藻(Synechococcus)的生物量分别增加29%和14%[100].值得注意的是,近海赤潮的产生和发展往往受到温度的控制[101],升温作用可能在特定海域促进了赤潮物种的短期快速增殖,进而影响近海海洋生态环境健康.一般认为,20~30℃是赤潮形成的适宜海水温度,而当海水温度急剧升高,如一周内升高大于2℃,可增加赤潮形成的可能性[102].
4 酸化与升温耦合对浮游植物种群组成及演替的影响
全球变暖和海洋酸化通常被称为气候变化的“邪恶双胞胎”[103],共同影响生物地球化学过程及生态系统.处于混合层内的海洋浮游植物还通常面临着多重环境因子的耦合作用[104],多重环境因子对生物的生理过程的交互作用通常有3种:叠加(additive)、协同(synergistic)和拮抗(antagonistic)作用[105].温度与CO2浓度的变化的耦合作用对浮游植物固碳作用等重要生理过程的影响不容忽视[7,106].升温和酸化能提高浮游植物群落的光合作用速率[34],将颗粒有机碳(POC)向溶解有机碳(DOC)转变,直接影响表层海水向深海的碳输出[107],进一步对某些浮游植物物种产生交互影响[108].
对海洋颗石藻而言,前期受控培养实验发现,在光照不限制的条件下,酸化和升温的交互作用使赫氏艾密里藻的颗粒无机碳的含量迅速降低[109],即降低了其钙化速率[110].然而,其他研究表明酸化和升温却协同升高了大洋桥石藻的无机碳与有机碳的生产速率[111].对海洋硅藻的研究表明,在酸化和升温的共同作用下,诺氏海链藻(Thalassiosira nordenskioeldii)、柔弱角毛藻(Chaetoceros debilis)和双突角毛藻(Chaetoceros didymus)的生长并没有受到显著的影响[40],而对中肋骨条藻(Skeletonema costatum)的生长却起到了协同促进的正效应[112–113].升温与酸化协同促进了超微型(0.2~2µm)浮游植物聚球藻的生长,而对原绿球藻的生长没有影响[85].
由于种间差异的存在,酸化和升温的耦合作用也将会对浮游植物群落的演替等产生影响.对北大西洋春季水华浮游植物群落的船载连续培养实验结果显示,尽管赫氏艾密里藻的丰度在高CO2分压和升温条件下均有所升高,但只有在二者同时升高的“温室”组才呈现出了最为显著的增高;并伴随着显著的钙化速率的降低,但在单一酸化或升温组别均未发现明显削弱的钙化速率[34].这与对北大西洋海盆区自然浮游植物群落的长期观测结果相似,即在过去的几十年间随着气候变化颗石藻的丰度呈现了显著的升高[114].对渤海湾近岸浮游植物群落的模拟培养实验则表明,温度和酸化对甲藻的丰度、硅藻和甲藻比值、硅藻组成及颗粒有机碳和颗粒有机磷(POP)的比值(C∶P)均产生了显著的交互效应,尽管酸化和升温均降低了硅藻和甲藻比值,但浮游植物种群的沉降速率只在二者同时变化的耦合作用下呈现出显著降低[81].相关的模型研究尽管还处于起步阶段,但也发现二者的耦合作用可能会改变浮游植物的生长速率达到20%~300%之多,进而对海洋生物地球化学循环产生深远影响[115].
5 展 望
CO2浓度升高引起的海洋酸化和海水升温现象在海洋环境中持续发生,影响浮游植物的生长和代谢过程,产生深远的海洋生物地球化学效应.浮游植物是生态系统的初级生产者,贡献约一半的全球初级生产力,在生物地球化学循环过程中发挥着重要作用.海洋酸化和升温对浮游植物的生理生态学效应可进一步通过食物网的传递影响到整个海洋生态系统,以及主要生源要素的生物地球化学循环,尤其是由真光层向深层海洋的碳输出,进而影响海洋的储碳能力,并对地球气候演变产生反馈效应.尽管目前针对海洋浮游植物对酸化和升温的生理学响应已有大量的研究积累,但二者的交互作用机制及长期进化学上的效应尚未厘清.此外,还要对酸化和升温及二者的耦合作用下不同区域的自然浮游植物群落的响应模式进行进一步认知,并总结出区域性响应规律.而且,相关的预测模型也要将气候变化中多重环境因子共同变化的复杂性纳入考量,以期对未来快速变化的海洋中浮游植物的变化趋势及其生物地球化学效应有更准确的预估.
