广灵大尾羊TP53INP2基因的克隆、生物信息学分析及其在不同脂肪组织中的表达研究
2022-04-15侯文欣潘洋洋车雨彤秦朋云赵祥乔利英刘建华刘文忠
侯文欣,潘洋洋,车雨彤,秦朋云,赵祥,乔利英,刘建华,刘文忠
(山西农业大学动物科学学院,山西太谷 030801)
在自然界中,生物体经常面临能量获取不足的情况,所以需要在能量充足时将多余能量储存起来。脂肪组织能够动态的储存能量,在机体能量消耗过多或摄入不足时供能,但过多的脂肪分解会减少体内脂肪沉积。脂肪沉积分为内脏脂肪沉积和肌内脂肪沉积,多余的内脏脂肪沉积会影响家畜产肉率,而肌内脂肪的沉积会提高肉质口感。羊肉营养价值较高,食之能暖身补身,深入探究羊肉脂肪沉积相关基因的重要性不言而喻。
肿瘤蛋白p53 诱导的核蛋白2(Tumor Protein 53-Induced Nuclear Protein 2,TP53INP2)又称糖尿病和肥胖调控蛋白(DOR),在人的淋巴细胞中首次发现。TP53INP2 蛋白能够作为甲状腺激素受体(TR)的转录辅激活因子参与甲状腺激素介导的成肌分化和成骨分化过程。TP53INP2 蛋白在细胞代谢中发挥双重作用。一方面,TP53INP2 蛋白通过促进rDNA 启动子处RNA 聚合酶I 前起始复合物的组装促进rDNA 转录,参与核糖体生物合成;另一方面,在营养缺乏的条件下,该蛋白在细胞核中与自噬相关蛋白LC3 结合,移至细胞质中参与细胞自噬。许多研究表明,自噬在脂肪形成和脂质代谢中发挥重要作用。近期研究表明,基因与脂肪细胞分化相关,但在小鼠和牛中所起的作用相反。过表达小鼠3T3-L1 前体脂肪细胞中的基因,发现形成的脂滴减少,脂肪细胞分化标志基因的表达量降低,表明基因抑制小鼠前体脂肪细胞的分化。但是过表达秦川牛前体脂肪细胞中的基因,导致形成的脂滴增加,脂肪细胞分化标志基因的表达量升高,表明基因促进牛前体脂肪细胞的分化。造成这种结果的原因可能是物种差异,也可能是基因参与的脂肪细胞分化通路不同。牛和羊均属于反刍动物,通过研究绵羊基因在前体脂肪细胞分化中的作用,与在小鼠和牛中的作用进行比较,可以初步判断是否因为物种差异导致上述结果。
目前有关绵羊基因的报道较少。本课题组前期高通量测序结果表明,基因在广灵大尾羊和湖羊尾部脂肪中表达量差异显著。广灵大尾羊为长脂尾型羊,湖羊为短脂尾型羊。前期高通量测序的差异结果提示该基因可能对脂肪沉积起作用。因此,本实验选用广灵大尾羊进行研究,克隆基因CDS 区,分析基因mRNA 序列及其编码蛋白的性质,并检测5 种脂肪组织中该基因的表达情况,为研究基因在绵羊脂肪代谢中的作用奠定基础。
1 材料与方法
1.1 样品采集 在屠宰场随机挑选4 只8 月龄的广灵大尾羊,屠宰后在无菌条件下迅速采集皮下脂肪、尾部脂肪、腹膜后脂肪、肾周脂肪和肠系膜脂肪组织。将各组织分装于标记好的冻存管后,迅速放入液氮中,后转至-80℃保存。
1.2 主要试剂 RNAiso Plus、PrimeSTARHS(Premix)、DL 2000 DNA Marker、DNA Loading Buffer、PrimeScriptRT reagent Kit with gDNA Eraser(Perfect Real Time)和TB GreenPremix Ex Taq(Tli RNaseH Plus)均购自日本TaKaRa 公司;Poly-Gel DNA Extraction Kit 购自美国Omega 公司;50×TAE 溶液、大肠杆菌DH5感受态细胞、琼脂糖和氨苄青霉素均购自北京索莱宝公司;pHBLV-CMVIE-ZsGreen-T2A-Puro 载体购自汉恒生物科技(上海)有限公司;ClonExpress Ultra One Step Cloning Kit 购自南京诺唯赞生物科技有限公司。
1.3 基因引物 根据NCBI 上绵羊的基因序列(XM_027977111.1),使用Primer Primier 5.0 软件和GenScript Primer Design 在线工具设计特异性引物,选用作为内参基因(所有引物见表1),引物均由Thermo Fisher 公司合成。

表1 引物信息
1.4 RNA 与cDNA 获取 参考Trizol 试剂盒说明书提取5 种脂肪组织的总RNA 后,吸取1 μL 打入核酸蛋白测定仪孔内,检测RNA 的浓度和纯度值(OD/OD)。检测合格后参考反转录试剂盒说明书合成cDNA,测浓度和纯度后放入-20℃保存备用。
1.5基因CDS 区克隆与测序 PCR 反应体系为15.8 μL:2×PrimeSTAR HS(Premix)7.6 μL,引物各0.6 μL,模板cDNA 1 μL,ddHO 6 μL。程序为:95 ℃ 5 min;98 ℃ 10 s,62 ℃ 10 s,72 ℃ 60 s,设为30个循环;72℃ 5 min;4℃终止反应。反应完毕后进行琼脂糖凝胶电泳检测,对正确条带快速切胶,并回收纯化目的片段。参照无缝克隆试剂盒说明书连接目的片段与载体后,转至感受态细胞中,涂抹在固体培养基上并在37℃培养箱中过夜。挑选5个单克隆菌落进行摇菌后,送至Thermo Fisher 公司测序。
1.6 生物信息学分析 使用DNAMAN Version 6(Lynnon Biosoft,USA)软件分析绵羊基因的测序结果;使用DNASTAR.Lasergene.v7.1(DNASTAR Inc.,USA)软件将该基因的mRNA 序列与山羊(XM_01805 7741.1)、黄牛(XM_003586843.5)、野猪(XM_003 359949.4)、小鼠(NM_178111.3)、大鼠(NM_00127 0947.1)、人类(NM_021202.3)和黑猩猩(XM_0011 60082.4)进行比对;使用Mega-X Version 10.1.8(Mega Limited,NZL)软件构建系统进化树;使用ProtParam Tool(https://web.expasy.org/protparam/)分析TP53INP2蛋白的理化性质;使用ProtScale(https://web.expasy.org/protscale/)分析TP53INP2 蛋白的亲疏水性;使用NetPhos 3.0 Sever(http://www.cbs.dtu.dk/services/NetPhos/)预测TP53INP2 蛋白的磷酸化位点;使用PSORT II(https://www.genscript.com/tools/psort)预测TP53INP2 蛋白的亚细胞定位;使用TMHMM Server v.2.0(http://www.cbs.dtu.dk/services/TMHMM/)预 测TP53INP2 蛋白的跨膜结构;使用SignalP 4.1 Server(http://www.detaibio.com/tools/signal-peptide.html)预测TP53INP2 蛋白的信号肽;使用NetNGlyc 1.0 Server(http://www.cbs.dtu.dk/services/NetNGlyc/)预测TP53INP2 蛋白的N-糖基化位点;使用NetOGlyc 4.0 Server(http://www.cbs.dtu.dk/services/NetOGlyc/)预测TP53INP2 蛋白的O-糖基化位点;使用SOPMA(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html)预测TP53INP2 蛋白的二级结构;使用Phyre 2(http://www.sbg.bio.ic.ac.uk/phyre2/phyre2 output/)构建TP53INP2 蛋白的三级结构模型。
1.7 实时荧光定量PCR 以1.4 中获取的cDNA 为模板进行实验。反应体系为10 μL:TB Green Premix Ex Taq II 5 μL;上、下游引物各0.4 μL;cDNA 2 μL;ddHO 2.2 μL。程序为:95℃ 5 min;95℃ 10 s,60℃15 s,72℃ 20 s,设为40个循环;95℃ 5 s,65℃ 1 min。拷贝反应结束后的结果用于数据分析。
1.8 统计分析 以腹膜后脂肪为对照,为内参,采用2法计算拷贝数据;使用GraphPad Prism 7.0(GraphPad Software Inc.,USA)软件作图并分析,差异显著判断标准为<0.05。
2 结果与分析
2.1基因CDS区克隆与测序PCR反应结束后的电泳结果显示,675 bp 处有一条清晰明亮的条带(图1)。测序结果经序列比对发现与绵羊(XM_027977111.2)目的序列完全一致,表明成功克隆出广灵大尾羊基因的CDS 区。
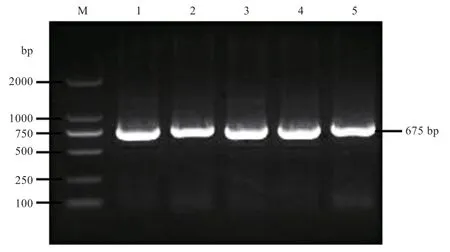
图1 TP53INP2基因PCR 扩增产物电泳图
2.2mRNA 序列比对与进化树构建 绵羊mRNA 序列与山羊、黄牛、大鼠、人类、小鼠、野猪和黑猩猩的相似性分别为99%、96.4%、65.5%、72.4%、65.4%、80.8% 和72.3%(图2)。Mega-X 软件构建的系统进化树中,绵羊与山羊亲缘关系最近,与大鼠和小鼠亲缘关系最远(图3),与序列相似性比对分析结果相一致。

图2 不同物种间TP53INP2mRNA 序列相似性比对
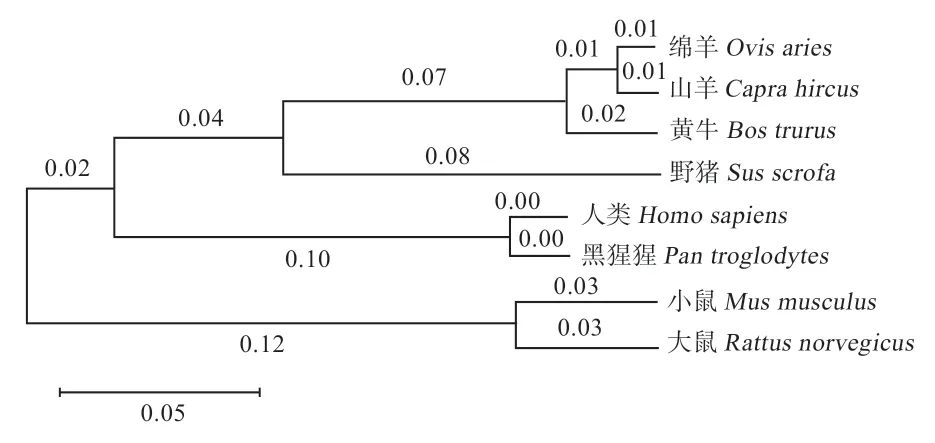
图3 不同物种间TP53INP2mRNA 序列进化树遗传距离关系
2.3 TP53INP2 蛋白理化性质与亲疏水性分析 将TP53INP2 蛋白序列输入ProtParam 软件中,结果显示该基因可编码224个氨基酸,TP53INP2 蛋白分子质量为24 320.28 u,理论等电点(pI)为5.54,分子式为CHNOS,总原子数为3 382;不稳定系数为77.63,是不稳定蛋白;亲水性总平均值为-0.596。利用ProtScale 软件做进一步的亲疏水性分析,结果如图4 所示,纵坐标负值表示亲水区,可见组成TP53INP2蛋白的氨基酸多集中在负值,整体表现为亲水性,与ProtParam 软件预测亲水性的结果相同。
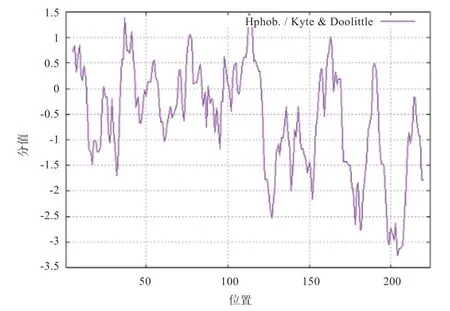
图4 TP53INP2 蛋白亲疏水性分析
2.4 磷酸化位点与糖基化位点预测 NetPhos 3.0 Sever软件预测发现,TP53INP2 蛋白潜在的磷酸化位点如图5 所示,丝氨酸、苏氨酸、酪氨酸位点分别存在14、2、1个。NetNGlyc 1.0 Server 和NetOGlyc 4.0 Server软件预测发现,TP53INP2 蛋白不存在N-糖基化位点,存在7个O-糖基化位点,分别位于第7、11、50、124、140、189、202 位氨基酸处。
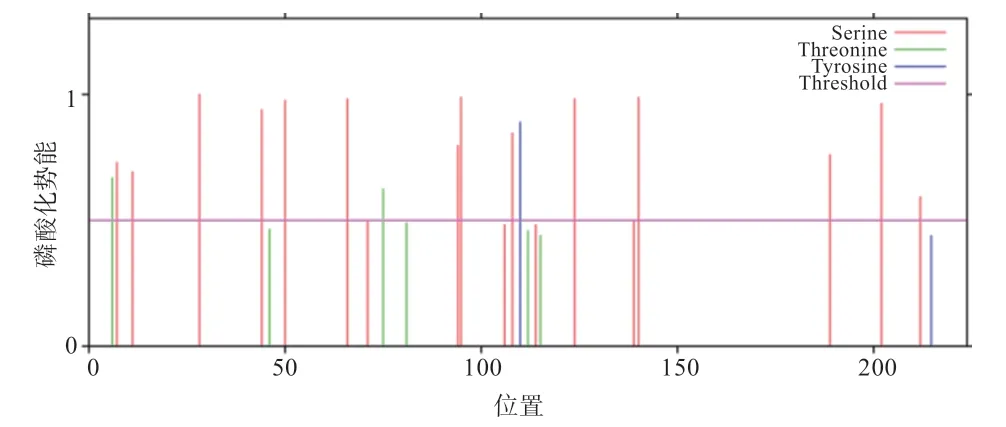
图5 TP53INP2 蛋白磷酸化位点
2.5 信号肽、跨膜结构与亚细胞定位预测 SignalP 4.1 Server 软件预测发现,TP53INP2 蛋白前70个氨基酸的C-score、S-score 和Y-score 均小于0.2,在阈值0.5之下,说明该蛋白不存在信号肽。TMHMM Server v.2.0软件预测表明,TP53INP2 蛋白不存在跨膜区。PSORT II 软件预测发现,TP53INP2 蛋白主要分布在细胞核(47.8%),少量分布在细胞质(8.7%)、液泡(4.3%)和线粒体(4.3%)中。
2.6 蛋白二级结构与三级结构预测 TP53INP2 蛋白二级结构的-螺旋、延伸链、-转角和无规则卷曲分别占23.66%、13.39%、1.79% 和61.16%(图6)。所构建的三级结构如图7 所示。

图6 TP53INP2 蛋白二级结构预测
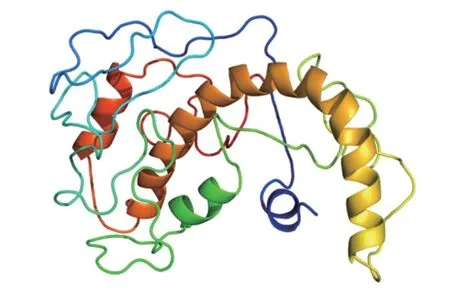
图7 TP53INP2 蛋白三级结构预测
2.7 实时荧光定量PCR 结果分析基因在广灵大尾羊5 种脂肪组织中均有表达,其中在皮下脂肪组织中表达量最高,其次是尾部脂肪组织,腹膜后脂肪组织中表达量最低;在皮下脂肪组织与其余4 种脂肪组织中均有显著差异;在腹膜后脂肪、肾周脂肪和肠系膜脂肪组织中的差异不显著(图8)。
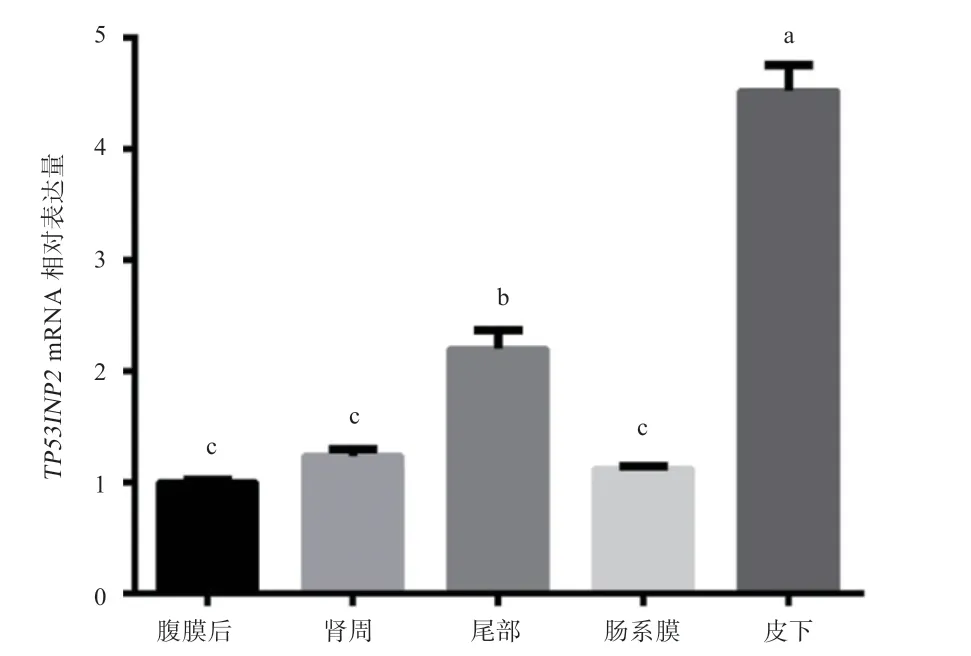
图8 TP53INP2基因在广灵大尾羊各脂肪组织的表达量
3 讨 论
自噬是一种细胞必需的、保守的自我进食过程,可降解可溶性蛋白、聚集蛋白、细胞器、大分子复合物和异物,该过程需要形成自噬体的双膜结构,最终与溶酶体融合。有研究表明,TP53INP2 能够调控哺乳动物和果蝇细胞中自噬体的形成来参与细胞自噬。在小鼠骨骼肌中,敲除基因,骨骼肌自噬水平下降,小鼠肌肉肥大,表明TP53INP2 通过激活自噬负调控小鼠骨骼肌的生成。此外,TP53INP2 还可以促进糖原合成酶激酶3(GSK3)的隔离,通过将其隔离到膜包裹的囊泡结构中激活WNT 通路抑制脂肪前体细胞的分化,而TP53INP2 隔离GSK3的作用依赖于自噬活性。近年来研究表明,在细胞核内TP53INP2能与LC3 结合,之后TP53INP2-LC3 复合物从细胞核移至细胞质,与自噬相关蛋白ATG7 相互作用,从而促进自噬体的生物发生。本研究预测显示,绵羊TP53INP2 蛋白大量定位于细胞核内,少量定位于细胞质,提示该蛋白通过自噬调控绵羊的肌肉和脂肪生成。
基因在进化上高度保守,研究表明,小鼠TP53INP2 氨基酸序列与哺乳动物的相似性均大于70%。在本研究中,绵羊基因的mRNA序列与哺乳动物的相似性均大于64.8%,说明该基因在进化上高度保守。蛋白理化性质预测显示,TP53INP2蛋白等电点为5.54。理论等电点可以在等电聚焦电泳以及使用离子交换层析分离纯化蛋白时作为参考。不稳定系数起判断目的蛋白稳定性的作用,当不稳定系数大于40 为不稳定蛋白。TP53INP2 蛋白不稳定系数为77.63,说明该蛋白是不稳定蛋白。蛋白质磷酸化主要发生在丝氨酸、苏氨酸和酪氨酸残基上,能够使蛋白质在具有电荷后结构发生改变,以此来调节和控制蛋白质活性。本研究预测显示,TP53INP2 蛋白存在17个潜在的磷酸化位点,说明该蛋白的功能可能由氨基酸的磷酸化产生。蛋白质糖基化主要分为N-糖基化和O-糖基化,对蛋白质折叠、可溶性、稳定性和亚细胞定位起重要作用。O-糖基化主要将糖基加至丝氨酸或苏氨酸上,与磷酸化存在竞争关系,这种竞争性修饰对转录很重要。TP53INP2 蛋白存在7个O-糖基化位点,说明该蛋白可能通过磷酸化位点与糖基化位点的竞争参与转录调控。信号肽是蛋白质N-末端的短肽链,靶向真核生物的分泌途径。TP53INP2 蛋白不存在跨膜区和信号肽,说明该蛋白不属于膜蛋白和分泌蛋白。有研究表明,TP53INP2 蛋白在人和大鼠中主要定位于细胞核内,与TR 相互作用,增强其转录活性。在本研究中,预测TP53INP2 蛋白主要定位于细胞核,提示该蛋白在绵羊中发挥同样作用。无规则卷曲容易被侧链相互作用影响,是造成蛋白不稳定的重要原因。预测显示,TP53INP2 蛋白二级结构的无规则卷曲占比高达61.16%,说明该蛋白稳定性差,这与理化性质预测该蛋白是不稳定蛋白的结果相符。
脂肪遍及全身各个地方,可将白色脂肪组织分为皮下脂肪和内脏脂肪。通常,皮下脂肪指所有皮肤下层的脂肪组织,尾脂也属于皮下脂肪,具有保温、储能及参与代谢等功能;内脏脂肪指沉积于内脏周边的脂肪组织,对内脏起保护作用,储能能力比皮下脂肪小。有研究表明,在秦川牛脂肪细胞中,过表达基因能引起基因表达量的升高,促进前体脂肪细胞的分化。在广灵大尾羊中,尾部脂肪基因的表达量高于肾周、小网膜和肠系膜脂肪,表现为皮下脂肪表达量高于内脏脂肪。基因在脂肪细胞增殖和分化过程中起主要调控作用,活化的基因能够增加脂肪细胞数量,促进脂肪细胞分化。在本研究中,基因在广灵大尾羊皮下脂肪的表达量最高,尾部脂肪的表达量高于肾周、肠系膜和腹膜后脂肪,整体表现为皮下脂肪表达量高于内脏脂肪,与基因的表达趋势相同。推测基因可能通过促进基因的表达促进绵羊前体脂肪细胞的分化。
4 结 论
TP53INP2 蛋白是一种不稳定的亲水性蛋白,存在17个潜在的磷酸化位点和7个O-糖基化位点,可能通过磷酸化位点与O-糖基化位点的竞争发挥转录调控作用。基因在广灵大尾羊5 种脂肪组织中均有表达,总体呈皮下脂肪表达量高于内脏脂肪,该结果提示基因可能有促进绵羊脂肪沉积的功能。本实验结果可为研究基因在绵羊脂肪代谢中的作用提供一定参考。
