二甲双胍和LPS通过转录因子RUNX2调控NFATc2基因的转录
2022-04-13薛晓阳李忠豪
T细胞活化的核因子(NFAT)是一类转录因子,在T细胞受体激活后负责调节淋巴细胞中细胞因子的表达。NFATc2是NFAT家族中的一员。NFATC2约占静止期T细胞中总NFAT的80%~90%,主要参与调节细胞因子如干扰素γ、白细胞介素4、IL-5 和IL-13的表达。NFAT蛋白在静止期T细胞中以磷酸化蛋白的形式存在于细胞质中,一旦T细胞通过T细胞受体被激活,T细胞内的钙离子水平持续升高进而使钙离子依赖性的钙调神经磷酸酶激活。活化的钙调神经磷酸酶进一步使NFAT蛋白发生去磷酸化,并移位至细胞核,从而驱动细胞因子和细胞表面受体等靶基因的表达。NFAT蛋白随后被糖原合酶激酶-3重新磷酸化并转移回细胞质。除了在淋巴细胞中起到关键作用外,NFAT蛋白在心肌细胞和骨骼肌细胞中也广泛表达,对心肌和骨骼肌的发育发挥重要的作用。
首先,重视预算编制,在预算编制中,重点关注经费支出数据,确保这类数据编制的合理性和有用性。其次,预算执行过程中,加强对经费开支数据的监控和审核,针对实际经费开支情况与预算数进行对比分析,查找原因,及时提出改进建议。最后,重视预算管理,有条件的事业单位,比如自收自支型事业单位,可以将经费开支情况与绩效考核相挂钩,严格经费管控,对经费超标行为严厉打击,防止资金浪费。
有研究发现,NFATc2抑制T辅助细胞(Th2)分泌Th2细胞因子,减轻Th2介导的过敏性免疫反应,同时抑制NFATC2会削弱Th2的反应,从而缓解化疗药物天冬酰胺酶引起的超敏反应。更有学者指出,NFATc2是人类黑色素瘤的潜在治疗靶点。目前国内外有关NFATC2的研究主要聚焦在NFATC2调节T细胞功能以及NFATC2在肿瘤筛查中诊断价值,然而有关转录因子调控NFATC2转录表达的研究鲜有报道,NFATC2在转录水平的调控仍然不明确。有研究初步证实转录因子RUNX2 是NFATC2 基因潜在的转录因子,但RUNX2如何上调NFATC2基因的表达以及RUNX2在NFATC2基因上的具体结合位点尚不清楚。RUNX2对NFATC2的转录调控仍需进一步明确。
图3为设计线宽为100 μm,方阻数为20的NiCr薄膜电阻的实际线宽。由图3可见,制备的薄膜电阻的线宽精度≤±5%。
因此,为更好地阐明NFATc2 表达的转录调控机制,本研究通过构建人NFATc2基因启动子的荧光素酶报告基因重组质粒pGL3-NFATc2-promoter,并转染到人胚肾上皮细胞293F中验证其启动子的转录活性,进一步运用点突变技术突变转录因子RUNX2潜在结合位点探究二甲双胍和脂多糖(LPS)对NFATc2转录活性的调节作用。
1 材料和方法
1.1 主要材料与试剂
293F细胞株、pGL3-basic质粒、pRL-TK质粒、细菌脂多糖LPS、二甲双胍由本实验室保存。基因组DNA提取试剂盒、感受态细胞DH5α、凝胶回收试剂盒、超保真DNA聚合酶、一步法克隆试剂盒、双荧光素酶报告基因检测试剂盒(南京诺唯赞生物科技有限公司)。DMEM高糖培养基、胎牛血清、Lipofectamine 3000转染脂质(Gibco)。限制性内切酶Ⅰ和d Ⅲ(NEB)。琼脂粉(Agar)、胰蛋白胨(Tryptone)、酵母浸出粉(Yeast Extract)(Oxoid)。引合成与测序由北京擎科生物科技有限公司完成。所用仪器:PCR 仪(Bio-Rad);电泳仪(北京百晶);凝胶图像分析仪Tanon-2500(上海天能公司);Spectra MAX M5酶标仪(Molecular Device);超微量蛋白核酸分析仪Biodrop(英国柏楉)。
1.2 细胞培养和DNA提取
所有实验结果均重复3次。应用SPSS 20.0统计软件进行统计分析,定量资料用均数±标准差表示。两组间比较采用两独立样本检验,多组间比较采用单因素方差分析,组间两两比较采用LSD检验,<0.05为差异具有统计学意义。
在纳入的10篇研究[2-11]中,均报道了治疗的疾病控制率,样本量共620例:替吉奥组309例,卡培他滨组311例。各研究间具有同质性(P=0.98,I2=0%),采用固定效应模型。结果显示,替吉奥组与卡培他滨组在疾病控制率上无明显差异,无统计学意义[OR=1.18,95%CI:(0.84,1.66),P=0.34],见图 1。
其次,这种思维差异体现在句子的时间顺序和逻辑顺序上。英语的语法中存在时态,可以通过动词形式的变化来表现动作发生的先后顺序,同时还可以使用大量分词和从句,用法灵活,可前可后。而汉语中不存在英语中的那些手段,叙事多靠并列结构,所以更加依赖句中各成分之间的顺序,尤其是汉语中的时间顺序和逻辑顺序。其中,较为典型的逻辑顺序差异就是:英语句中习惯“先果后因”,而汉语却喜欢“先因后果”。因此,在英译汉时,我们就需要在理解句意的基础上,按照事情发生的先后顺序或者根据事件中所包含的逻辑顺序,适当地调整译文的顺序,使其更符合汉语思维,如下例:
1.3 PCR扩增人NFATC2基因启动子片段和回收纯化
在USCS基因数据库中查找人NFATC2基因的启动子序列,选取长度为2170 bp(-2155 bp~+15 bp)的DNA 序列作为本项目研究的启动子片段。应用CE Design V1.04引物设计软件,以Ⅰ和dⅢ为酶切位点分别设计上游引物5'-atttctctatcgataggtaccCCC CAGGCCCCTGCAGTA-3'和下游引物5'-cagtaccggaa tgccaagcttCGCGAGCTTCCTGCTCCG-3'。以基因组DNA为模板扩增目的启动子片段:在PCR反应管中依此加入ddHO,基因组DNA,上下游引物,2×Phanta Max Buffer,dNTP Mix,PhantaMax超保真DNA聚合酶。PCR反应条件如下:95 ℃预变性3 min,95 ℃变性15 s,58 ℃退火15 s,72 ℃延伸150 s,共35个循环,72 ℃彻底延伸5 min。反应结束后,将PCR产物进行1%琼脂糖凝胶电泳,在切胶仪上切下目的产物并利用凝胶回收试剂盒回收PCR产物。同时用相同的下游引物分别构建2077 bp NFATC2 基因启动子片段(上游引物5'-atttct ctatcgataggtaccCTGATGCCTAAGTCCCAACCC-3')、1802 bp NFATC2 基因启动子片段(上游引物5'-attt ctctatcgataggtaccTGGAATGATGGATGGCTTGG-3'),1651 bp NFATC2基因启动子片段(上游引物5'-atttctct atcgataggtaccCCCGTCTAATCTTCTCATCCTCA-3'),1083 bp NFATC2基因启动子片段(上游引物5'-atttctc tatcgataggtaccTTTAGTTACTGGGTCCTTATGCAGT G-3')和323 bp NFATC2基因启动子片段(上游引物5'-atttctctatcgataggtaccCTGATGCCTAAGTCCCAACCC-3')。
1.4 pGL3-basic质粒双酶切及回收纯化
应用限制性内切酶Ⅰ和dⅢ对pGL3-basic质粒进行双酶切,酶切条件为:pGL3-basic 质粒2 μL(2 μg),Ⅰ(10 U/μL)2 μL,dⅢ(20 U/μL)2 μL,10×CutSmart Buffer 5 μL,ddHO 39 μL,共50 μL。37 ℃酶切4 h,反应完成后将酶切产物进行1%琼脂糖凝胶电泳和凝胶回收。
1.5 重组和转化
将293F细胞用含10%胎牛血清的DMEM培养基培养。转染前1 d接种2×10细胞于24孔板中,在37 ℃、5%CO培养箱中培养。直至细胞融合度达到80%时,按照Lipofectamine 3000试剂盒操作说明,分别以0.8 μg/孔pGL3-basic质粒(对照组)和pGL3-NFATC2-promoter质粒(实验组)的剂量瞬时转染293F细胞,同时以0.1 μg/孔pRL-TK海肾荧光素酶表达质粒作为转染效率内参照,每组分别设置3个复孔。
1.2 纳入标准 (1)参考《中国心力衰竭诊断和治疗指南 2014》[4]确诊为慢性心力衰竭;(2)参考《室性心律失常中国专家共识》[5]确诊为室性心律失常;(3)NYHA心功能分级Ⅲ—Ⅳ级;(4)左室射血分数(LVEF)≤40%;(4)用药及随访依从性高;(5)对本研究知情且签署同意书。
1.6 阳性克隆的鉴定和质粒抽提
质粒转染24 h后,吸掉培养基,每孔细胞用PBS洗涤两次。根据Vazyme公司的双荧光素酶报告基因检测试剂盒(Dual Luciferase Reporter Assay Kit)操作说明,加入100 μL/孔细胞裂解液,室温下振摇裂解5 min,吹打并吸取细胞裂解产物至1.5 mL离心管中,12 000 g常温离心2 min。吸取20 μL细胞裂解上清至酶标板中,加入100 μL/孔Luciferase Substrate,迅速混匀后读取萤火虫荧光素酶发光值。然后再加入100 μL Renilla底物工作液(萤火虫荧光素酶终止液和海肾荧光素酶的底物),迅速混匀后读取海肾荧光素酶发光值。每组细胞重复测量3次,以萤火虫荧光素酶发光值与海肾荧光素酶发光值的比值作为衡量NFATC2启动子转录活性的依据。
1.7 NFATC2启动子荧光素酶报告基因质粒转染293F细胞
将回收得到的NFATC2启动子片段与pGL3-basic质粒片段应用重组克隆试剂盒进行重组反应。反应结束后取10 μL 重组产物与50 μL 感受态细胞DH5α混匀,冰上静置30 min,42 ℃热激90 s,冰上放置3 min,加入600 μL LB 培养基(不含抗生素),37 ℃摇菌1 h(220 r/min),5000 r/min离心1 min,加入100 μL培养基重悬,取50 μL 涂于LB 固体培养平板(含Ampicillin)上,37 ℃培养箱中倒置培养过夜。
1.8 NFATC2启动子活性鉴定
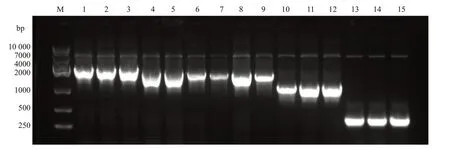
挑取5个菌落接种于800 μL的LB液体培养基(含Ampicillin)中,37 ℃摇菌4 h,取1 μL菌液进行菌液PCR鉴定,挑选菌液PCR鉴定为阳性的菌液送至北京擎科生物科技有限公司测序进一步鉴定。将测序鉴定正确的阳性克隆进行扩大培养并提质粒。
1.9 二甲双胍和LPS对NFATc2的转录调控
293F 细胞转染pGL3-1651 bp NFATC2-promoter质粒24 h,分别给予二甲双胍浓度梯度(2.5、5、10 mmol/L)和LPS浓度梯度(2、5、10、20µg/mL,)处理24 h后收集细胞测定各组荧光素酶活性。接着分别将pGL3-1651 bp NFATC2-promoter 质粒和pGL3-1651bp NFATC2-promoter突变质粒分别转染293F细胞24 h,接着给予二甲双胍(10 mmol/L)和LPS(5µg/mL)处理24 h,收集细胞测定转录因子RUNX2结合位点突变前后各组荧光素酶活性。
1.10 统计学方法
293F细胞用RPMI 1640培养基接种至3.5 cm细胞培养皿,待细胞密度达95%,收集细胞。按照基因组DNA 提取试剂盒操作说明提取293F 细胞的基因组DNA,应用超微量核酸分析仪Biodrop测定提取的基因组DNA产物浓度。
2 结果
2.1 NFATC2启动子序列的扩增
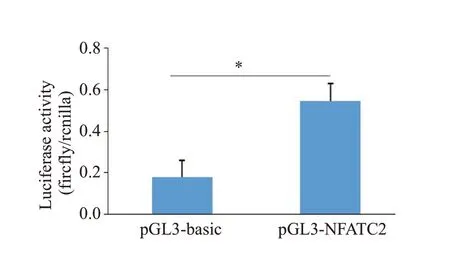
将pGL3-basic 空载质粒和pGL3-NFATC2-promoter 质粒分别转染293F 细胞,24 h 后测得转染pGL3-basic 空载质粒组的荧光数值为0.18±0.18,转染pGL3-NFATC2-promoter 质粒组的荧光数值为0.55±0.09,pGL3-NFATC2-promoter 质粒组比pGL3-basic空载质粒组的荧光素酶活性高约3.04倍(=3.178,df=4,=0.0336,图5)。
2.2 pGL3-NFATC2-promoter载体的构建
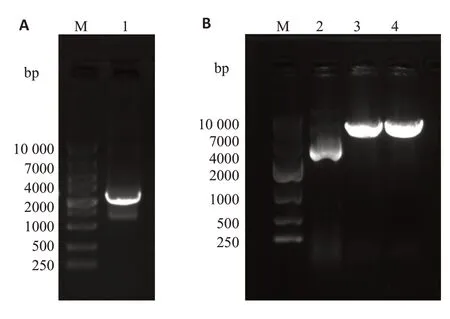
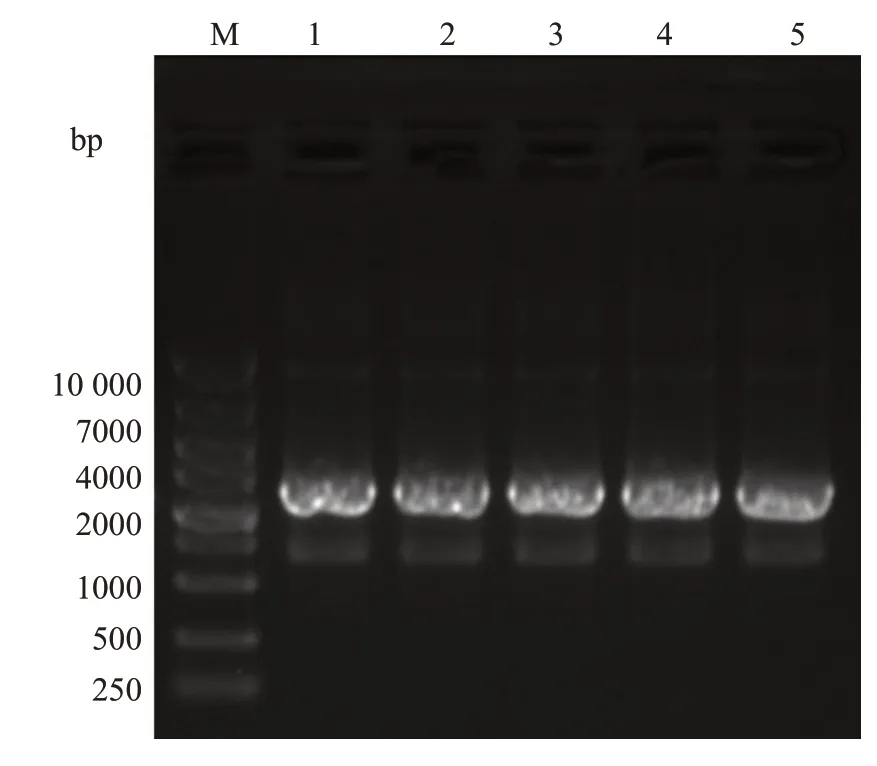
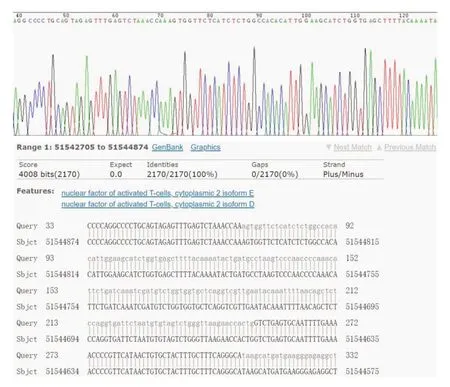
应用限制性内切酶Ⅰ和dⅢ对pGL3-basic质粒进行双酶切,结果见图1B,泳道2为未进行双酶切的pGL3-basic质粒,泳道3-4为双酶切的pGL3-basic质粒,由于双酶切破坏质粒的超螺旋结构,因此酶切的质粒在琼脂糖凝胶中的迁移速度较慢。凝胶回收酶切后的pGL3-basic质粒,与NFATC2启动子片段进行重组连接、转化大肠杆菌DH5α。挑取5个平板克隆进行菌液PCR鉴定(图2),5个克隆均在2000 bp附近可见清晰条带,与目的片段大小基本一致。挑选阳性克隆测序,测序结果显示NFATC2基因启动子成功插入到pGL3-basic 载体中,所构建质粒与设计相符(图3),且未发现碱基突变或缺失。同时,按照相同的方法利用已构建好的pGL3-NFATC2-promoter重组质粒为模板,构建不同片段长度的NFATC2基因启动子报告基因载体,分别为2077 bp(-2062 bp~+15 bp)、1802 bp(-1787 bp~+15 bp)、1651 bp(-1636 bp~+15 bp)、1083 bp(-1068 bp~+15 bp)和323 bp(-308 bp~+15 bp),凝胶电泳结果显示各个片段扩增条带与相应目的片段大小一致(图4)。
2.3 NFATC2启动子报告基因转染细胞后荧光素酶表达
琼脂糖凝胶电泳结果显示(图1A),在2000 bp附近可见清晰条带,与NFATC2基因启动子PCR扩增片段大小相符。对该片段进行凝胶回收,测定浓度为136 ng/μL。
海岛是我国海洋国土的重要组成部分,也是海洋经济发展及国防建设的重要基点。海岛工业污染源较少,生活垃圾和生活污水是海岛的主要污染来源;随着海岛的开发和建设大力推进,生活垃圾和污水产量日益增加[1-3]。海岛生活垃圾处理方式和设施的选择,必须根据海岛生活垃圾产生现状和理化特征等基本资料和实际情况进行设计。国内对生活污染源的研究主要集中于内陆农村地区,海岛地区生活污染物排放规律、理化特性研究较少。我国海岛大部分面积较小,多为村镇级海岛。笔者以广东省2个村镇级海岛(外伶仃岛、东澳岛)为研究地点,对生活垃圾产生量、理化特性和生活污水排放系数、污染物浓度进行研究。
2.4 NFATC2启动子不同片段长度的荧光素酶表达量
为了探究NFATC2启动子上的转录活跃区域及其潜在转录因子,我们构建了6 个不同片段长度的NFATC2启动子报告基因载体,分别是pGL3-2170 bp、pGL3-2077 bp、pGL3-1802 bp、pGL3-1651 bp、pGL3-1083 bp、pGL3-323 bp,并将其分别转染293F细胞,结果显示6组差异有统计学意义(=121.6,<0.001)。进一步运用Dunnett's多重比较检验,显示pGL3-1802 bp组与pGL3-2170 bp组差异没有统计学意义(>0.05),其余各组与pGL3-2170 bp组之间差异均有统计学意义(<0.05),而且转染pGL3-1651 bp质粒达到最大的荧光素酶表达量(图6)。

2.5 LPS和二甲双胍对NFATC2的转录调控作用
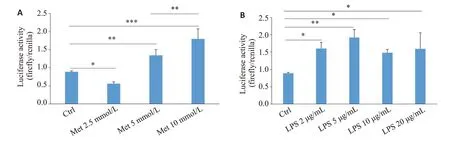
将pGL3-1651bp报告基因质粒转染到293F细胞,分别使用LPS和二甲双胍刺激细胞。LPS组4个不同浓度之间差异有统计学意义(=6.811,=0.0065),而且均对NFATC2启动子的转录活性有激活作用,其中以5 µg/mL 的效果最显著(图7B)。然而二甲双胍对NFATC2 启动子的转录具有双向调节作用(=33.10,<0.0001),低浓度的二甲双胍(2.5 mmol/L)对NFATC2的启动子起抑制作用,而中高浓度的二甲双胍(5、10 mmol/L)则对NFATC2启动子的转录具有激活作用。
为进一步探究LPS和二甲双胍调控NFATC2转录的潜在机制,将pGL3-1651 bp质粒上转录因子RUNX2的结合位点进行点突变(图8A),构建pGL3-1651 bp突变质粒(pGL3-1651bp-Mutation),并将转染到293F细胞。结果显示,RUNX2的结合位点突变后,LPS和二甲双胍对NFATC2的转录激活作用下调(=0.0179,图8B;<0.0001,图8C)。

3 讨论
NFATc2可以抑制成年动物软骨细胞的生长和分化,小鼠敲除NFATc2基因后导致关节外软组织中的细胞自分化形成软骨,而老年小鼠中一旦NFATc2蛋白丧乏功能将导致软骨细胞不受控制地增殖。此外,胶质母细胞瘤中NFATc2的表达明显上调,NFATc2敲低后显著抑制胶质母细胞瘤的增殖。由此可见NFATC2 在各种病理生理过程中扮演极其重要的作用。近年来越来越多的研究表明NFATC2与黑色素瘤的生长和转移密切相关[,提示NFATC2很可能是黑色素瘤的潜在治疗靶点。因此,本研究旨在通过构建NFATC2荧光素酶报告基因探讨NFATC2在转录水平上的表达调控机制,为临床筛查抗肿瘤药物的分子生物学机制奠定基础。
本研究成功构建了人NFATc2基因启动子的荧光素酶报告基因,并在293F细胞中验证了其转录活性。与此同时,本研究构建了6个不同片段长度的NFATc2基因启动子报告基因载体,发现不同片段长度的NFATc2 基因启动子的转录活性不尽相同,与长度为2170 bp 的NFATc2 基因启动子相比,长度为1651、1083、323 bp的NFATc2基因启动子片段的转录活性显著增加,而长度为2077 bp和1802 bp的NFATc2基因启动子片段的转录活性或降低或不变,这与Kannan等研究结果基本一致。令人惊喜的是,我们证实片段长度为1651 bp的NFATc2基因启动子的转录活性最高,提示NFATc2基因启动子-1636 bp~+15 bp的区域可能为NFATc2基因的转录活跃区域。有研究表明骨髓来源的树突状细胞暴露于高剂量LPS可上调NFATc2的表达以及核移位,增加细胞凋亡从而阻止实验性自身免疫性葡萄膜视网膜炎进展。本研究实验结果也发现在293F细胞转染了NFATc2报告基因24 h后再给予不同浓度的LPS刺激24 h均能增加NFATc2报告基因的活性,在LPS 5µg/mL的浓度下NFATc2报告基因的活性最高,当LPS浓度提高到10或20µg/mL,NFATc2报告基因的活性有所下降,说明LPS对NFATc2转录的调节的最适剂量是5µg/mL。二甲双胍是2型糖尿病患者使用最广泛的降糖药物,目前国内外还没有研究阐明二甲双胍对NFATc2的调控作用,我们发现二甲双胍对NFATc2报告基因的转录活性具有双向调控作用,2.5 mmol/L的二甲双胍显著抑制NFATc2报告基因的转录活性,而5 mmol/L和10 mmol/L的二甲双胍明显增加NFATc2报告基因的转录活性,本研究首次证实二甲双胍可从基因转录层面调控NFATc2的表达。
RUNX2是成骨细胞分化必不可少的最上游转录因子,序列分析揭示NFATC2基因启动子上存在多个转录因子RUNX2的潜在结合位点,成纤维细胞、间充质细胞和软骨细胞中过表达RUNX2 显著增加NFATc2 基因的转录活性,提示NFATc2 基因可能是RUNX2的潜在下游调控基因,但RUNX2与NFATC2基因启动子上哪个位点结合并发挥转录激活作用尚未证实。因此,我们将NFATc2基因启动子-1636 bp~+15 bp区域的一个潜在RUNX2结合位点(-1632 bp~-1626 bp)进行点突变,发现突变前后NFATc2报告基因活性并没有明显变化,然而突变后再给予LPS或二甲双胍刺激,NFATc2 报告基因的活性均显著受到削弱,由此证实LPS 和二甲双胍诱导NFATC2 的转录激活依赖于RUNX2 在NFATC2 启动子上的结合位点(-1632 bp~-1626 bp),提示NFATC2 基因启动子上-1632 bp~-1626 bp的RUNX2结合位点很可能是RUNX2的一个主要调控位点,这在既往研究中未见报道。
综上所述,本研究成功构建人NFATC2基因启动子的荧光素酶报告基因,并证实NFATc2 基因启动子-1636 bp~+15 bp的区域为NFATc2基因的转录活跃区域。我们首次证实LPS和二甲双胍对NFATc2的转录调节作用,LPS和二甲双胍对NFATc2的转录调节依赖于转录因子RUNX2,且依赖于NFATC2基因启动子上-1632 bp~-1626 bp的RUNX2结合位点。后续我们将在此报告基因质粒的基础之上围绕NFATC2的转录调控,深入探讨LPS和二甲双胍在相关的人类疾病模型中的具体作用及其分子机制。
