Selaginella orientali-chinensis,a new resurrection spikemoss species from southeastern China based on morphological and molecular evidences*
2022-04-12,,,,
,,,,
1. State Key Laboratory of Biocontrol/ Guangdong Provincial Key Laboratory of Plant Resources,School of Life Sciences,Sun Yat-sen University,Guangzhou 510275,China
2. Administrative Commission of Danxiashan National Park,Shaoguan 512300,China
Abstract:Selaginella tamariscina(Beauv.)Spring and S. pulvinata(Hook. et Grev.)Maxim. are two closely related rosette forming and resurrection spikemoss species widespread in Asia. In this paper,Selaginella orientali-chinensis Ching et C. F. Zhang ex H. W. Wang et W. B. Liao was described and illustrated. This species was closely related to S. tamariscina and S. pulvinata and previously misidentified as S. tamariscina. By the gross morphological and molecular evidences,we found that this species is easily distinguished from S. tamariscina and S. pulvinata by the absolutely dichotomously branched main stems. Our field works confirmed that this species is widely distributed in southeastern China and mainly inhabits Danxia landforms areas. In some cases,this species shares the same habitats with S. tamariscina. Illustrating the morphological,phylogenetic and distribution characters of this new species would provide new data for the taxonomic studies on the genus Selaginella in the future.
Key words:Selaginella;Danxia landforms;morphology;phylogeny;distribution
Among the three extant lycophyte families,the largest one(Selaginellaceae,or spikemoss)contains only one genusSelaginellabut more than 700 living species,which occupy a variety of habitats in every continent except Antarctic[1-4]. Some spikemoss species are well-known for their capability of desiccation tolerance(DT)and resurrection. For example,members of theRosulataeandS. lepidophyllaclades that newly validated by Zhou et al[1]can survive under complete dehydration for as long as a decade and have therefore gained attention by researchers who focus on the mechanism of plant DT[5-7]. In Asia,the extensive seasonally arid environments are ideal habitats of resurrection spike mosses , such asS. tamariscinaandS. pulvinata[8-9]. These two species resemble each other in morphology and are widely distributed across Siberia,Mongolia,East Asia,Qinghai-Tibet Plateau,Indian Subcontinent,Indo-China Peninsula,and Philippines,but occurred in different habitats.S. pulvinatais usually in alpine areas,whileS. tamariscinaprefers lower elevation and warmer environment[10].
Among specimens ofS. tamariscinacollected from Danxiashan National Geological Park(Guangdong Province,China)in 2016,several distinct specimens were selected out for further analysis,and confirmed to be an unrecognized species closely related toS. tamariscinaandS. pulvinatabased on morphological and molecular phylogenetic evidences. To obtain more distribution information of this new species and its two closely related species ,we attempted to examine specimens ofS. tamariscinaandS. pulvinatadeposited in several herbaria(CSFI,CSH,CZH,GXMG,FJFC,IBSC,JIU,LBG,PE,SYS). To our surprise,dozens of specimens of this new species which were ever misidentified asS.tamariscina,were screened out. Interestingly,two specimens in PE labeled with“typus”indicated that this species had ever been informally denominated as“Selaginella orientali-chinensis”by R. C. Ching and C. F. Zhang in 1982(H. H. Chung 979(PE)),but was treated asS. tamariscinaagain in 1991 by X. C.Zhang. Here,we describe and illustrate this new species,Selaginella orientali-chinensisChingetC. F. ZhangexH. W. WangetW. B. Liao.
1 Method
1.1 Samples collection and morphological analysis
A total 34 samples,including 14 ofS. orientalichinensis,14 ofS. tamariscinaand 6 ofS. pulvinata,were collected from six locations for molecular and morphology analysis(Table 1). Morphological features were examined from fresh samples and dried specimens. Voucher specimens were deposited at Sun Yat-sen University Herbarium(SYS).
1.2 DNA extraction, sequencing and sequence treatment
A green portion of each individual was cleaned with tap water and sterilized with commercial bleach.Genomic DNA was extracted with the CTAB protocol[11]. ITS andrbcL fragments were amplified with primer pairs ITS 1 and ITS 4[12]andrbcL 192 F andrbcL 1324 R[3],respectively,in the 20 μL reactions containing 0.2 mmol/L of each dNTP,1.0 μmol/L of each primer,1× EasyTaq Buffer,1 U EasyTaq and 1 μL of DNA extract.PCR conditions are as described by Wu et al[3]. Amplicons were resolved by 1% agarose gel and those with the expected sizes(ca.650 bp for ITS and ca. 1 kb forrbcL)were purified and sequenced with ABI 3730XL analyzer (Applied Biosystems, Foster City, California, USA).Sequences ofrbcL and ITS were deposited in GenBank (accession numbers and voucher see Table 1).
1.3 Phylogenetic analysis
Using the ITS andrbcL sequences of the new species as query,BLASTn searches were performed on the NCBI website,the top hits were obtained fromS. tamariscinaandS. pulvinata. Previous research on the phylogeny ofSelaginella[1,3]showed that in the clade Rosulatae,S. tamariscinaclustered first withS. pulvinata,thenS. stauntoniana,and then a monogroup includingS. imbricata,S. pilifera,S. heliocladaandS. digitata). So ITS andrbcL sequences of the above mentioned species were downloaded from NCBI website(accession number see Table 1). Together with those obtained in this study,the sequences were aligned with SeqMan,DNAStar[13]and checked manually. Phylogeny trees based on ITS andrbcL sequences were constructed with the maximum parsimony(MP),maximum likelihood(ML)and Bayesian inference(BI)methods. MP analysis was performed in PAUP* v. 4.0b10[14]and all characters were weighted equally. The most parsi-monious trees were obtained with heuristic searches of 1 000 replicates random stepwise sequence addition(RAS)replicates,and MP bootstrap values(BS)were calculated with 1 000 replicates. ML analysis was performed in IQ-TREE[15],and the appropriate nucleotides substitution model was searched by ModleFinder implemented in it[15]. ML Bootstrap percentages(BP)were calculated based on 1 000 fast bootstrap replicates(bb=1 000). BI analyses was carried out in MrBayes v 3.2.7[16]. Among the three supported models,F81 was selected for its best perform,and substitution rate was set as Gamma,while other parameters were set as default,and sampled the tree every 1 000 generations of 1 100 000 generations. The first 25% of samples were discarded as burn in,and the remaining trees were used to calculate a 50% majority-rule consensus topology and posterior probability(PP)values.
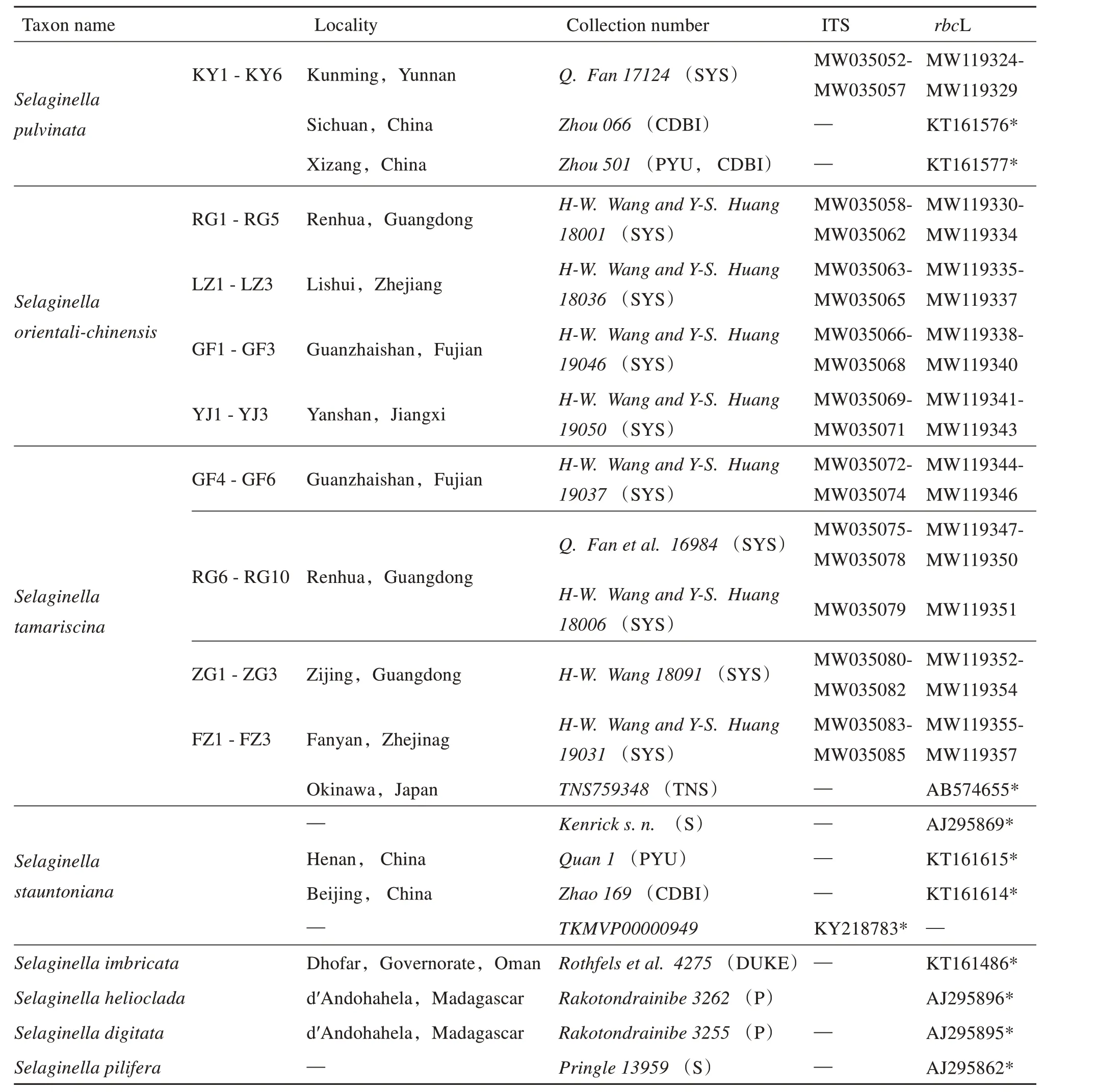
Table 1 Materials and the GenBank accession number of ITS and rbcL sequences obtained in this study
Further,an ILD test[17](incongruence length difference)was performed on ITS andrbcL databases and no conflict was detected between them. So ITS andrbcL sequences were concatenated to perform phylogenetic analysis to discriminateS. tamariscina,S. pulvinata,S. orientali-chinensis,usingS. stautonianaas outgroup.
2 Results and discussion
The aligned ITS andrbcL sequences were 531 bp and 1 018 bp in length,and no heterozygous sites were detected. The phylogenetic tree constructed from ITS orrbcL sequences showed thatS. orientalichinensis,S. tamariscinaandS. pulvinataformed a monogroup as expected,though species relationship among the three species was not strongly supported(Fig. 1). As ITS andrbcL sequences were concatenated,the phylogenetic tree showed that individuals of each species formed monogroups with high supports,S. orientali-chinensisclustered first withS. tamariscina,and thenS. pulvinata,though with low supports(Fig.2).
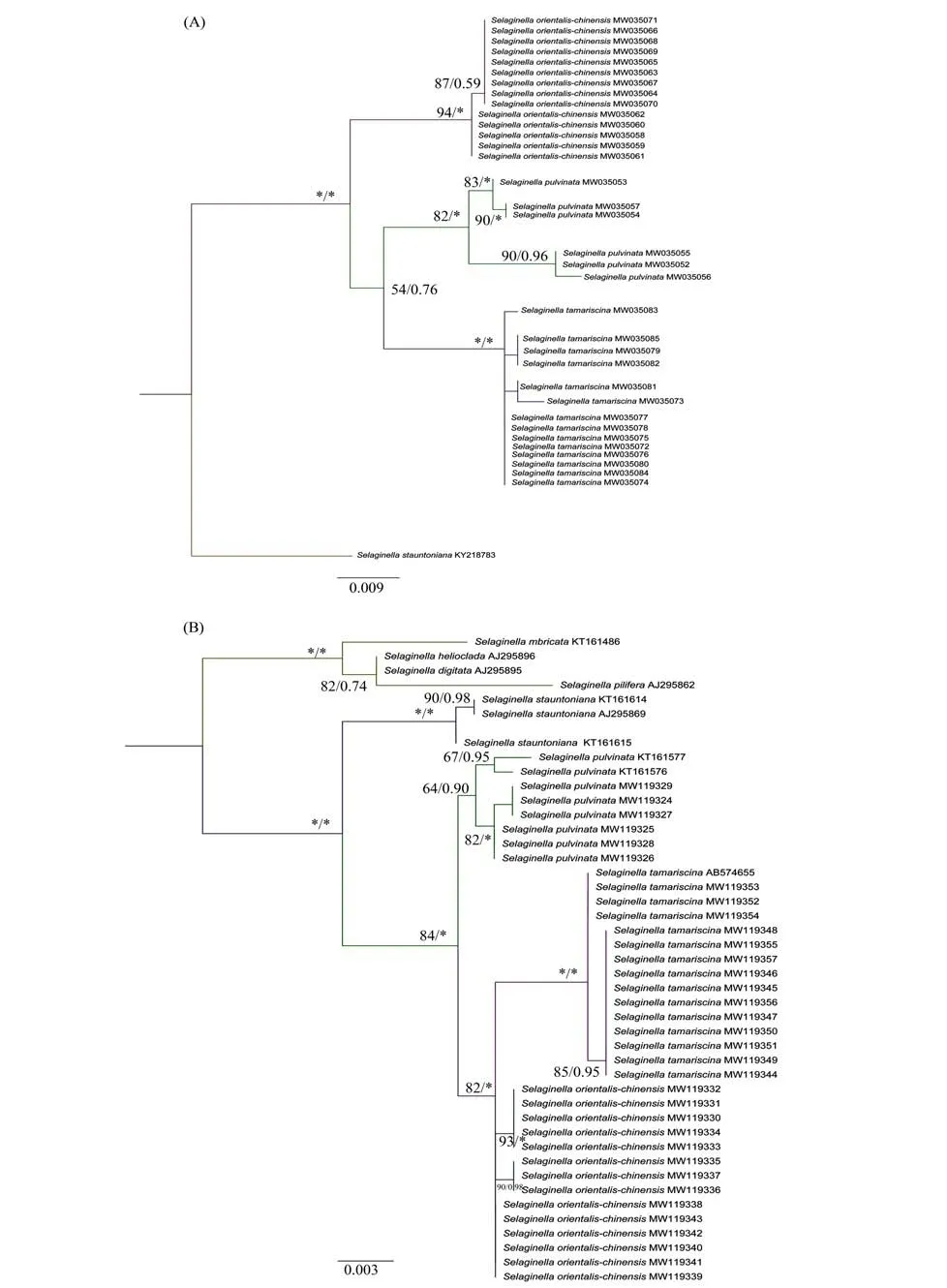
Fig.1 The ML phylogenetic tree constructed from ITS sequences(A)and rbcL sequences(B)of Selaginella orientali-chinensis and other closely related species

Fig.2 The 50%majority rule consensus tree derived from Bayes inference presenting the relationship between Selaginella orientali-chinensis,S. tamariscina and S. pulvinata
Although the plant form, sporangium and spore morphology are similar in the three species,S. orientali-chinensiscan be distinguished fromS. tamariscinaandS. pulvinataeasily. The branches ofS. orientali-chinensiswere absolutely dichotomously branched,while the branches ofS. tamariscinaandS. pulvinatawere pinnately branched(Figs. 3-5).Additionally,the differences amongS. orientalichinensis,S. tamariscinaandS. pulvinataare also obvious. The former two species usually form a slender tree-like trunk mainly composed of rhizophores and roots,whileS. pulvinataalways forms compact cushion due to the persistent withered branches.According to our own field investigations and other specimens and photography records,S. orientalichinensisis not as widespread asS. tamariscina,but is still widely distributed in Guangdong,Guangxi,Fujian,Hunan,Jiangxi,and Zhejiang provinces of China.S. pulvinatamainly inhabits cold and alpine environment,including the Himalayan,the Yunnan-Guizhou Plateau,West Sichuan,Qinling Mountains,Taihang Mountains,Northeast China and Siberia.While,Selaginella orientali-chinensisandS. tamariscinacommonly prefer seasonally arid habitats,and in some cases,co-occurs (Fig. 5). However,S. orientali-chinensisis mainly distributed in Danxia landforms,whileS. tamariscinacan be observed in multiple landscapes.

Fig.5 Morphology comparison between Selaginella orientali-chinensis and S. tamariscina
3 Taxonomic treatment
Selaginella orientali-chinensisChingetC. F.ZhangexH.W. WangetW.B. Liao,sp.nov.
1)Type. China. Guangdong:Renhua County,Danxiashan National Nature Reserve,24°56'N,113°45'E,290 m a. s. l.,23 May 2020,Q. Fan 17870(holotype, SYS! isotypes IBSC! PE! SYS!)(Fig.3-5).
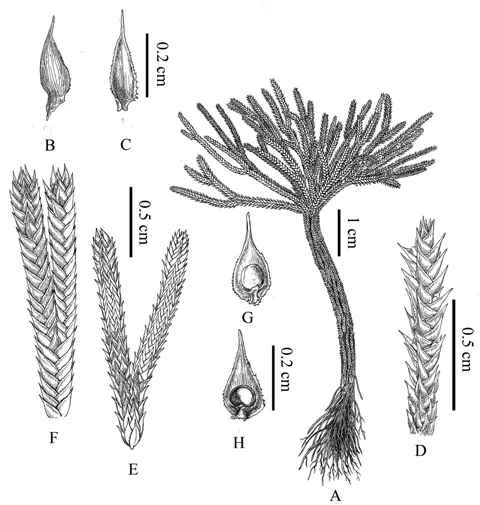
Fig.3 Selaginella orientali-chinensis Ching et C. F. Zhang ex H. W. Wang et W. B. Liao(Drawn by Yun-Xiao Liu)
2)Diagnosis. The new species is similar toS.tamariscinaandS. pulvinatain plant form and the morphology of dorsal leaves,ventral leaves,and sporophylls. However,the main stems ofS. orientalichinensisare absolutely dichotomously branched,while those ofS. tarmariscinaandS. pulvinataare pinnately branched. Additionally,S. pulvinatausually inhabits alpine,plateau,and temperate region,and the tree-like trunks usually form compact cushions,butS. orientali-chinensisandS. tamariscinaare distributed in the mountains or lowland of subtropical regions,and have tree-like trunks that are slender and do not form compact cushions.
3)Description. Plants terrestrial or epilithic,resurrectional;stems forming rosette,evergreen or seasonally green;stems,rhizophores and roots entangled forming treelike trunk,1-20 cm in length and ca.0.5 cm in diam. Rhizophores restricted to base of stem,0.5-20 cm,slender and adhering to the treelike trunk,forked and forming rootstock near the ground.Stems straminous or brown,terete,not sulcate,dichotomously branched,not crooked under dehydration. Branches sparse and regular,dorsiventrally flattened,2-3 mm wide including leaves. Axillary leaves symmetrical,ovate,ovate-triangular,or elliptic,(1.5-2.5)mm×(0.5-1.0)mm,base exauriculate,margins denticulate and obviously white-margined. Dorsal leaves imbricate,asymmetrical,elliptic,(1.5-3.0)mm×(0.5-1.5)mm,not carinate,base obtuse,not peltate,margin denticulate,obviously white-margined,apex aristate,spreading or parallel to axis. Ventral leaves slightly ascending,overlapping,asymmetrical,ovate to triangular or oblong-ovate,(1.5-2.5)mm×(0.5-1.2)mm,apex aristate;basiscopic margin subentire,serrate or ciliolate (at base),revolute ;acroscopic base enlarged,broader,overlapping stem and branches,margin lacerate or denticulate. Strobili solitary,terminal,compact,tetragonal,(12-15)mm×(1.2-2.6)mm;sporophylls uniform,white-margined and hyaline, ovate-triangular, margin denticulate,membranous,apex acuminate or aristate;megasporophylls ± randomly distributed on both sides of strobilus;microsporangia transversely elliptic,relatively thick;microspores yellowish orange,megaspores pale yellow.
4)Distribution and ecology.Selaginella orientali-chinensisis widely distributed in southern and eastern China (Guangdong, Guangxi, Fujian,Hunan,Jiangxi and Zhejiang provinces),growing on bare rock and grassy slope at 100 to 1 000 m a.s.l.,and mainly in Danxia landforms.

Fig.4 Selaginella orientali-chinensis Ching et C. F. Zhang ex H. W. Wang et W. B. Liao
5) Conservation status. Due to its wide distribution and numerous individuals in each investigation site,Selaginella orientali-chinensisshould be classified as the Least Concern(LC)category according to IUCN Categories[18].
6)Etymology. The specific epithet "orientalichinensis" was designated by the famous fern botanists Qin Renchang and Zhang Chaofang.
7)Key toSelaginella orientali-chinensisand related species.

8)Paratypes. China. Fujian:Putian,17 Feb 1923,H. H. Chung 979(PE);Hunan:Youxian,22 Sept 2015,G.X. Chen et al. LXP-06-5369(JIU).
9)Additional specimens examined. China.Fujian:s.n.,11 Oct 1974,Anonymous 0284(PE);
Zhanghu,31 July 1984,ⅡNan 84434(FJFC);Dehua,1 Aug 1958,Nanjing University 23387(FJFC);Cong′an,1 Aug 1958,East China Normal University 48230(FJFC);Ningde,1 Sept 1958,Fujian Agriculture and Forestry University 30180(FJFC);
Guangdong:Fengshun,6 Aug 2009,C.F. Zeng and Q.Y. Zeng ZXF 7379(CZH);s.n. ,3 Aug 2014,X. L. Zhou et al. ZXL09943(CSH);Guangxi:
Tengxian,31 Oct 1960,Z. M. Liu 13282(PE);Guiping, 28 Oct 2017,Guiping survey team450881171028062LY(GXMG);Hunan:Liuyang,3 Aug 1959,J. Y. Xiao 00018(CSFI);Jiangxi:Guixi,4 Nov 1979,S.J.Sheng and D.F.Huang 399(LBG);Zixi,4 Oct 1985,S.S. Lai and D.F.Huang 160(LBG);Zixi,26 July 2015,B.C.Guan NCU201507 MTS0085(JXU);Suichuan,1 June 1984,J. L.Wang and D.F. Huang 19(LBG);Jinggangshan,16 Nov 1982,J. L. Wang et S. C. Zhang 8210131;Zhejiang:Suichang,26 June 1980,P.X.Qiu and G.H. Yao 5674(PE);Qinyuan,10 Oct 1977,P. X.Qiu 4300(PE);Qinyuan,24 Feb 1991,C.Z. Zheng and C.F. Zhang 7926(PE).
