土壤生化特性在模拟氮沉降条件下对土壤呼吸和N2O排放的影响
2022-04-09张婷周金蓉冯廉洁陈玲蒋静艳
张婷 周金蓉 冯廉洁 陈玲 蒋静艳
0 引言
人为源导致大量活性氮进入大气层,经过一系列转化后,部分以干湿沉降的形式回到地表,对陆地生态系统造成了很大的影响[1].全球氮沉降逐年增加,从1850年至今,陆地生态系统可溶性氮沉降增加了3倍,海洋可溶性氮沉降增加了2倍,未来还将逐渐增加[2].近几十年中国氮沉降也呈急剧增加趋势,从1961年的12.64 kg·hm-2·a-1(以N计,下同)增加到2008年的20.07 kg·hm-2·a-1[3-4].无论是实测结果还是模型模拟均表明氮沉降有巨大的时空变异性,反映在我国就是氮沉降量西低东高[5-7],2013年的定点实测结果表明中国东南部氮湿沉降量均大于 60 kg·hm-2·a-1,而西北部地区则小于9 kg·hm-2·a-1[5].2000—2014年间的实测结果也表明整个扬子江流域无机氮沉降量较高,年均为33.2 kg·hm-2·a-1[7],因干沉降对氮沉降的贡献难以定量,故此结果还有很大的不确定性.
大量氮沉降将对土壤碳氮循环产生影响[1].CO2和N2O作为两大温室气体,也是土壤碳氮循环的中间产物,氮沉降对其有何影响?已有报道认为土壤温室气体排放对氮沉降的响应主要取决于土壤氮的饱和度和氮沉降水平[8-11].在氮饱和的土壤中,氮增加减少了土壤呼吸[11-12]和CH4吸收速率[10],增加了土壤N2O的排放[9].在氮限制的松树林土壤中,氮增加对土壤呼吸和CH4吸收速率没有显著影响,高氮也增加了土壤N2O的排放[8-10].长期氮沉降对温带森林土壤呼吸和N2O排放因季节而异.氮增加对温带森林生长季和春季冻融期CO2排放均无显著影响,但在生长季降低了72.8%~85.7%的N2O排放通量,而在冻融期增加了11.2~17.8倍N2O排放[13].此外也有报道认为短期氮沉降处理对青藏高原高山草甸CO2和 N2O没有显著影响[14].另有研究则认为氮沉降无论是对农业土壤还是森林土壤均增加了N2O 的排放,但CO2排放对氮增加不敏感,CH4和N2O是氮增加的敏感响应因子[15-16].上述结果出现了不一致性,主要源于研究对象的单一性.土壤CO2和N2O均来源于土壤微生物的作用,不同土地利用方式下的土壤生化性质不同[17],其对氮沉降的响应可能不同[18].已有研究也表明土壤呼吸和N2O排放与土壤理化性质有关[18-19],如土壤pH、有效氮、土壤质地、土壤碳氮比(C/N)等为影响N2O排放的重要因子[18,20],土壤CO2排放与土壤碳含量和阳离子交换量呈正相关,与游离铁含量呈负相关[21].但结合生物特征分析氮添加条件下不同土壤特性对土壤呼吸和N2O排放影响的研究尚不多见.
本研究基于高氮沉降地区江苏省不同土地利用方式下(包括农、林、草地及果园和竹园)的13种土壤不同生物和理化特性,通过模拟氮沉降的室内培养试验,分析不同土地利用方式下的土壤呼吸和N2O 排放对氮添加的响应,确定氮添加条件下影响土壤呼吸和N2O排放的主要土壤决定因子,结果可为了解在氮沉降的条件下,不同土壤特性对土壤呼吸和N2O排放的影响提供科学依据.
1 材料与方法
1.1 供试材料
江苏省地处中国东部(116°18′~121°57′E,30°45′~35°20′N),地形以平原为主,境内年降雨量在1 000 mm 左右,年均气温13~16 ℃.江苏地理上跨越了南北地区,气候和植被也同时具有南方和北方的特征.不同土地利用方式主要包括林地、草地、农田、果园和竹林等.供试土壤共13种,分别代表江苏省境内不同土地利用方式下的土壤,包括了林地(W)、农田(F)、草地(M)、果园(O)及竹林(B)等.采样地点如图1所示,从北至南分别为徐州2个(F1和W1)、连云港2个(F2和W2)、涟水3个(O1、O2和F3)、洪泽1个(M1)、盐城1个(M2)、南京3个(W3、W4和F4)和宜兴1个(B1).

图1 采样地点示意Fig.1 Locations of the sampling sites
供试土壤采自地表0~20 cm,多点混合后,除去植物残体等杂物,取部分新鲜土样,放于4 ℃冰箱中冷藏,供有效氮、土壤微生物等指标的测定,另取部分土样风干后过筛,供土壤pH值、质地、有机碳、全氮等指标的测定.
1.2 实验设计
将供试土样过筛混匀后称取100 g(以烘干土质量计)置于590 mL的玻璃瓶内,调节并保持土壤水分为0.30 g·g-1(模拟旱地条件),每个土样中添加6.349 mg NH4NO3(相当于江苏省的年平均氮沉降量50 kg·hm-2),置于25 ℃培养箱中培养.同时做对照,即每种土样不添加氮源直接进行室内培养.加氮处理代码为T,不加氮处理代码为CK.每处理3重复,恒温恒湿(重量法调节)培养.CO2和N2O的排放速率测定采用间歇密闭培养-气相色谱法.每天采样1次,每次取样前抽取背景样品,并用胶塞封口,密闭培养2~4 h,用外接三通阀的60 mL的注射器从培养瓶上部空间抽取40 mL样品气体,采完气后敞口继续恒温恒湿培养.至每种土样T和CK的CO2和N2O 气体连续2次排放速率无差异为止,共培养20 d.
气体样品用改装的Agilent 4890D气相色谱仪同步测定N2O和CO2的浓度[22].N2O测定所用载气为99.999%高纯氩甲烷(Ar2和CH4的体积比为 95∶5),检测器为电子捕获检测器(ECD),检测温度为330 ℃.CO2测定所用载气为99.999%高纯氮,检测器为氢火焰检测器(FID),检测温度为200 ℃.根据气体的分子量、培养温度、培养容器内气体的有效空间、培养时间内气体的浓度差和培养容器内烘干土的质量,计算各气体的排放速率,具体计算公式如下:
F=Δm/(W·Δt)=ρ·V·Δc/(W·Δt),
(1)
式中:F为气体排放速率;V为培养容器内气体的有效空间(L);ρ为气体密度(g·L-1),ρ=(气体相对分子质量×273)/[22.4 ×(273 +T)];Δc为Δt时间内的气体浓度差;T为培养温度(℃);W为培养容器内的烘干土质量(g);Δt为培养时间(h).CO2和N2O的累积排放量由其每天平均排放速率和培养时间累积计算得出.
1.3 土壤理化性质及微生物特征的测定

MC=EC/0.38,
(2)
MN=EN/0.45,
(3)
其中:EC,EN分别为熏蒸土样的有机碳量或全氮与未熏蒸土样之间的差值(mg·kg-1);0.38、0.45分别为氯仿熏蒸杀死的微生物体中的碳(C)或氮(N)被浸提出来的比例.土壤微生物细菌(BA)、放线菌(AC)和真菌(FUN)数量采用稀释平板涂布法测定[24-25].
1.4 数据处理
结果用平均值±标准误表示.用OriginPro 2021软件制作图表,用SPSS 23.0进行统计分析.
2 结果与分析
2.1 供试土壤生化特征
表1所示的是供试土壤的土壤类型、利用方式及各种基本理化、生物性质.13种供试土壤包括黄棕壤(W3、F4)、褐土(F1、W1)、棕壤(F2、W2)、潮土(G1、G2、F3)、棕红壤(B1)、黄褐土(W4)、草甸土(M1)、滨海盐土(M2)等江苏省主要土壤类型,包括农田(F1、F2、F3、F4)、林地(W1、W2、W3、W4)、果园(O1、O2)、草地(M1、M2)、竹林(B1)等不同土地利用方式.表1中供试土壤各基本理化性质的变异系数范围为18.4 %~131.3 %,变化范围较宽,说明各供试土壤性质差异较大,具有一定的代表性.

表1 供试土壤生物和理化特征Table 1 Physical and bio-chemical properties of the tested soils
2.2 氮添加后不同土壤间CO2和N2O排放速率的差异分析
图2所示的是代表土壤呼吸和N2O排放低(W1)、中(F3)、高(O2)的3类典型土壤的CO2和N2O排放速率随培养时间的动态变化情况.CO2-CK即CO2对照,为未添加氮源的土壤排放的CO2;CO2-T即CO2处理,为添加氮源的土壤排放的CO2;N2O-CK即N2O对照,为未添加氮源的土壤排放的N2O;N2O-T即N2O处理,为添加氮源的土壤排放的N2O.由图2可以看出,不同土壤的CO2和N2O排放速率峰值时间不同,部分土壤培养初期即出现最大值,如O2,部分土壤峰值出现时间推迟至培养的第3天,如F3.所有土壤培养15 d之后,土壤CO2排放速率(以C计)和N2O排放速率(以N计)在T和CK之间没有显著差异(P>0.05).

图2 典型土壤W1、F3和O2的CO2和N2O排放速率的动态变化Fig.2 Variations in CO2 and N2O emission rates of typical soils (W1,F3 and O2)during incubation
2.3 氮添加对不同土壤CO2和N2O累积排放的影响
图3为不同土壤的CO2和N2O累积排放量.由
图3可以看出,即使同种土地利用方式下的不同土壤类型的CO2和N2O累积排放量(分别以C、N计)也各不相同.无论是否添加氮源,均是土壤O2的CO2累积排放量最高,显著高于其他土壤(P<0.05),W1的土壤呼吸累积排放量最低.未添加氮源即CK中,土壤F4的N2O累积排放量最高,显著高于O2(P<0.05).而添加氮源后即T处理中土壤O2的N2O 累积排放量最高,显著高于其他土壤(P<0.05).无论是否添加氮源,B1、F1、M2、O1、W1和W4的N2O排放均较低.
不同土壤CO2和N2O排放对氮添加的响应不同.T与CK处理之间,土壤F1、W1和W3的土壤呼吸无显著差异(P>0.05),土壤F4、M2、W1和W2的N2O排放无显著差异(P>0.05);其他土壤无论是CO2还是N2O排放,T处理与CK间均有显著差异(P<0.05).为了解氮源添加对土壤CO2和N2O排放的影响,进一步计算了不同土壤外源单位氮的CO2和N2O排放因子:外源单位氮的CO2排放因子是指相对于CK,每添加1 g N土壤多排放的CO2-C量(%,即100×g(CO2-C)· g(N)-1),代码为EFCO2;外源单位氮的N2O排放因子是指添加的外源氮转化成N2O-N的百分比(%,即100×g(N2O-N)· g(N)-1),代码为EFN2O.结果见图3.外源氮的加入对不同土壤的呼吸速率刺激效果不同,对原本处在中等呼吸速率水平的土壤F2和W2的刺激效果最高,即外源单位氮的CO2排放因子F2和W2较大,而F1和W1最低;外源单位氮的N2O转化率仅F2和O2较高,其他土壤均较低,尤其W1最低.
将增氮显著促进CO2和N2O排放的土壤种类归为组Ⅰ,增氮未显著影响的土壤种类归为组Ⅱ.显著促进CO2排放的土壤即组Ⅰ包括B1、F2、F3、F4、M1、M2、O1、O2、W2、W4,组Ⅱ包括土壤F1、W1和W3;显著促进N2O排放的土壤即组Ⅰ包括B1、F1、F2、F3、M1、O1、O2、W2、W4,组Ⅱ包括土壤F4、M2、W1和W3.计算组Ⅰ和组Ⅱ各指标平均值的比值,选取与数值1偏离度25%以上的因子作为两组土壤有较大差异的指标,具体结果见图4箭头所示处.从图4中可以归纳出:增氮显著促进CO2排放的土壤具有较低的黏粒成分、土壤微生物碳和微生物碳氮比,较高的土壤基础呼吸、土壤有效氮及细菌和真菌数量.增氮显著促进N2O排放的土壤则具有较高的土壤基础呼吸,较低的内源N2O排放和较低的土壤总氮、有效氮、放线菌和真菌数量.
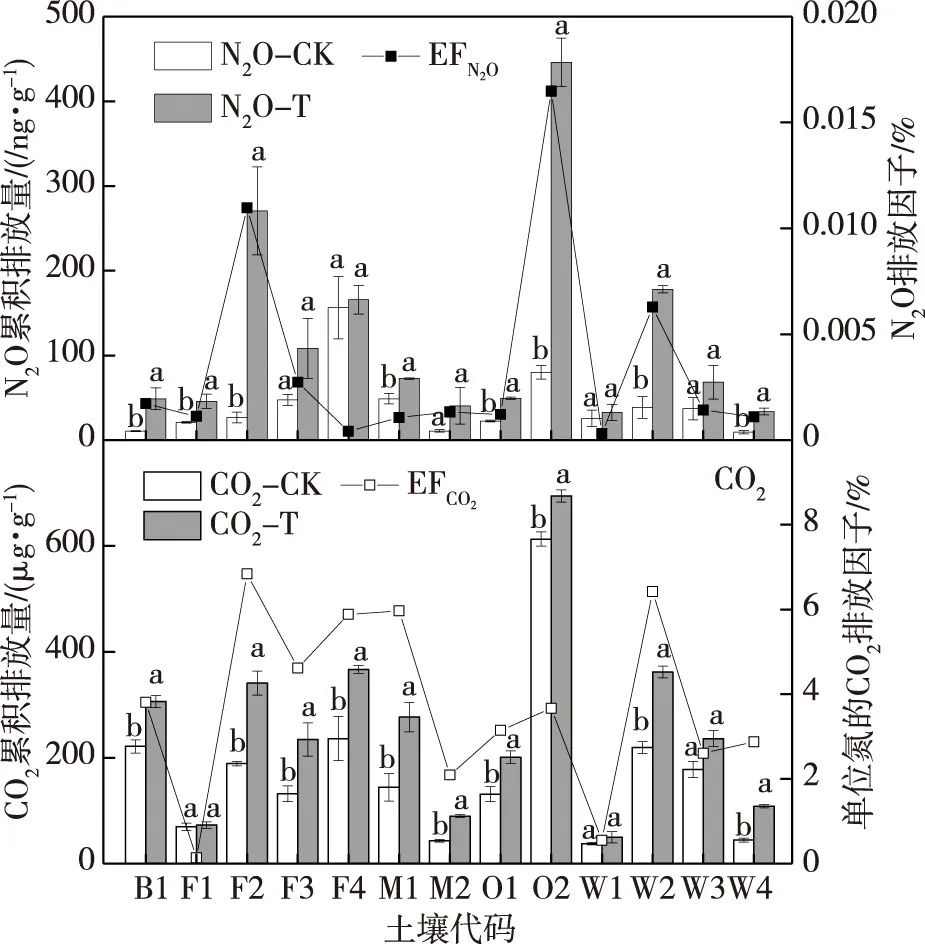
注:同一土壤代码上不同小写字母表示对照和处理在0.05水平上差异显著.图3 不同土壤CO2和N2O的累积排放量及单位氮的CO2和N2O排放因子Fig.3 Cumulative emissions and emission factors per gram nitrogen for CO2 and N2O from different soils
为进一步反映组Ⅰ和组Ⅱ的特征,令增氮处理T和对照CK的CO2累积排放量的差值ΔCO2=CO2-T-CO2-CK,N2O累积排放量的差值ΔN2O=N2O-T-N2O-CK,将此计算结果以及典型的土壤性质对土壤组别作BOX-PLOT箱图(图5).从图5中可以看出,尽管不同土壤的生物和化学特性差异较大,呈非正态分布(中位线和均值点有偏离),但增氮显著促进CO2排放的土壤组(组Ⅰ)的真菌数量明显较高于无显著影响组(组Ⅱ),而微生物碳氮比则明显小于组Ⅱ.增氮显著促进N2O排放的土壤组(组Ⅰ)则是放线菌数量和硝态氮含量均明显小于无显著影响组(组Ⅱ).

注:横坐标上的代码AN为有效氮,即铵态氮和硝态氮的和.图4 增氮显著促进CO2(a)和N2O(b)排放的土壤组(组Ⅰ)与未显著影响组(组Ⅱ)各指标的比值Fig.4 Ratios of soil parameters of Group Ⅰ (affected groups,in which nitrogen addition significantly promotes greenhouse gas emissions)to those of Group Ⅱ (unaffected groups,in which nitrogen addition has little effect on soil greenhouse gas emissions),(a)for CO2 and (b)for N2O
2.4 土壤呼吸和N2O排放对氮添加响应的主要土壤驱动因子
表2所示为土壤呼吸和N2O排放与土壤生化特性之间的相关性.由表2可以看出,未添加氮源条件下,N2O排放与铵态氮含量呈极显著正相关(P<0.01),与有机碳、全氮、有效氮、细菌、放线菌和真菌数量呈显著正相关(P<0.05),与其他因子无明显相关关系(P>0.05).CK的土壤呼吸即基础呼吸与细菌数量及微生物碳呈极显著正相关(P<0.01),与微生物氮呈显著正相关(P<0.05),与其他因子无明显相关关系(P>0.05).在添加氮源条件下,即T的土壤的N2O排放与基础土壤呼吸和细菌数量呈极显著正相关(P<0.01),与其他因子无明显相关关系(P>0.05).T的土壤呼吸与基础土壤呼吸和细菌数量呈极显著正相关(P<0.01),与微生物碳、氮呈显著正相关(P<0.05),与其他因子无明显相关关系(P>0.05).此外,添加氮源后即T组N2O排放和土壤呼吸具有极显著正相关关系(P<0.01),而CK组未见二者有显著相关关系(P>0.05).外源单位氮的CO2排放因子与有机碳、全氮、真菌数量呈显著正相关(P<0.05),与MC/MN呈显著负相关(P<0.05),与其他因子无明显相关关系(P>0.05).外源单位氮的N2O转化率与基础土壤呼吸和细菌数量呈极显著正相关(P<0.01),与其他指标无明显相关关系(P>0.05).
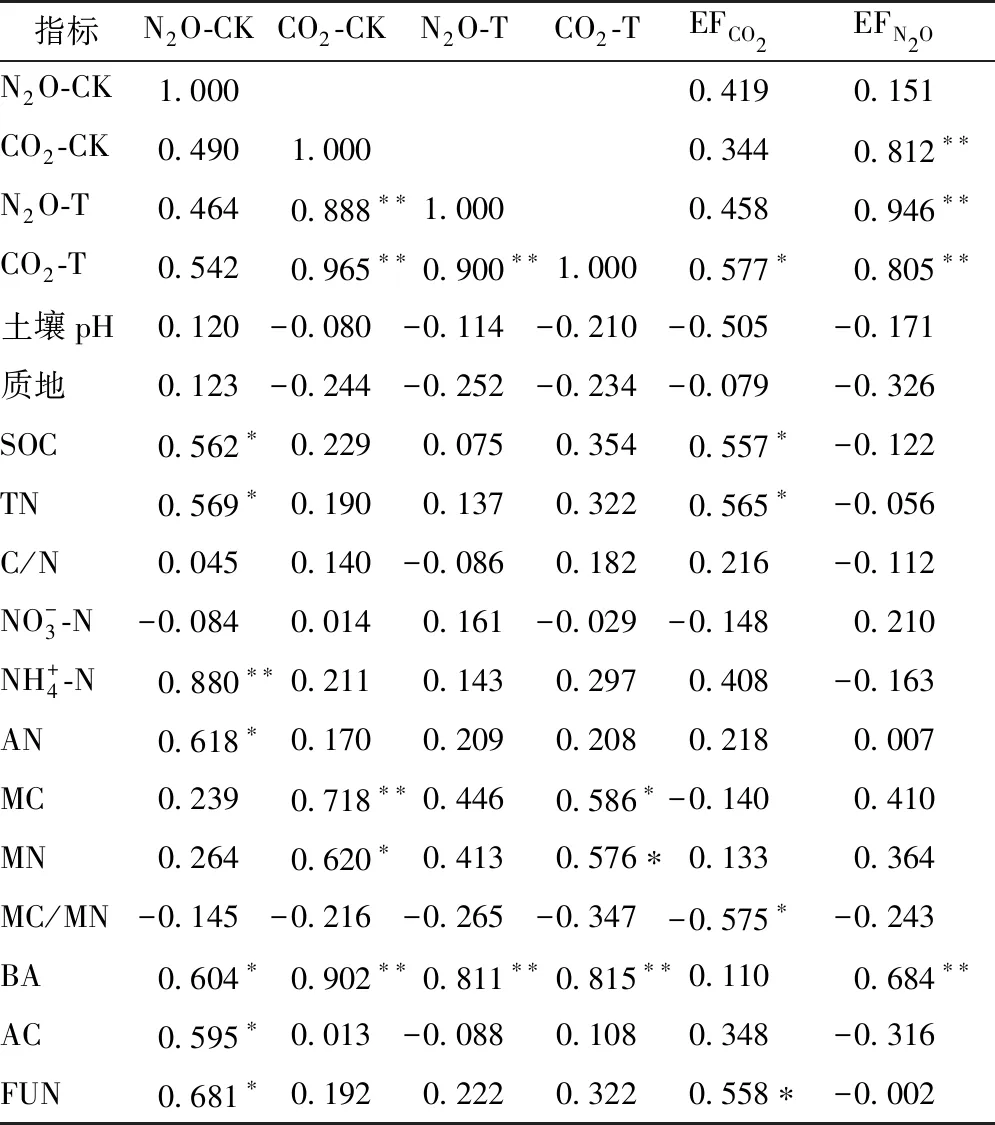
表2 土壤CO2、N2O累积排放量、单位氮的CO2和N2O排放因子与土壤生化因子的相关系数(n=13)Table 2 Pearson correlation coefficients of cumulative CO2 and N2O emissions,CO2 and N2O emissions factors per gram nitrogen against soil biochemical parameters (n=13)
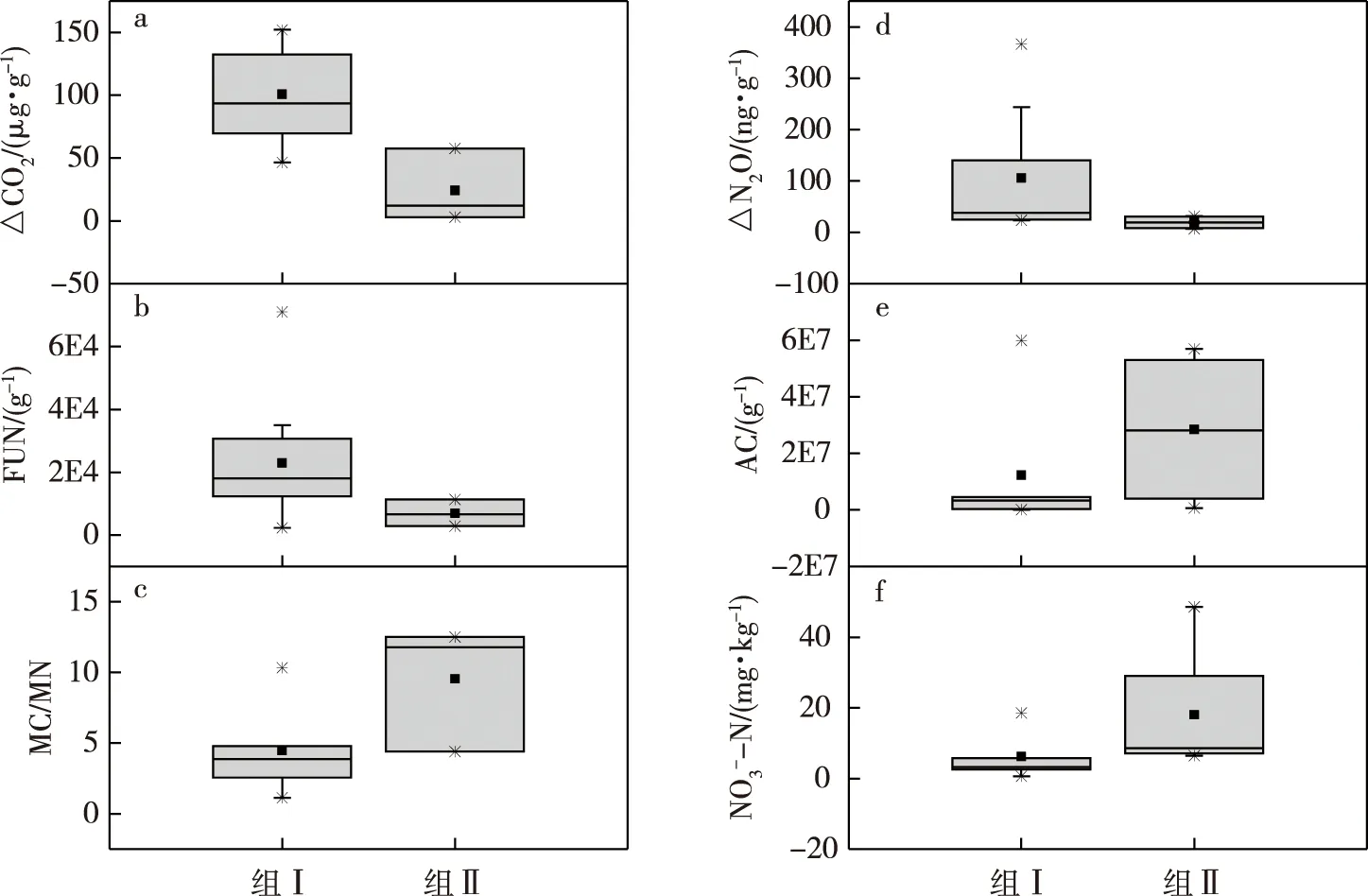
注:a—c分别反映组Ⅰ和组Ⅱ之间的△CO2、真菌和微生物碳氮比的差异;d—f分别反映组Ⅰ和组Ⅱ之间△N2O、放线菌和硝态氮的差异;箱顶和箱底分别为上下四分位数,■表示均值,*表示的是1%~99%范围.图5 典型指标对应土壤组别的箱形图Fig.5 Boxplots of soil properties versus the soil groups
进一步将土壤CO2和N2O累积排放量与土壤因子之间的关系进行逐步回归分析,回归方程见表3.结果表明,在本研究0.30 g·g-1的水分条件下,无论是否添加氮源,土壤呼吸均主要由土壤细菌数量和pH来决定,且与细菌数量成正比,与pH呈反比.未添加氮源处理的土壤N2O排放主要是由土壤细菌和铵态氮含量决定,添加氮源条件下,土壤N2O排放主要是由土壤细菌数量决定,且与之成正比.外源单位氮的CO2排放因子主要取决于MC/MN,与之成反比,外源单位氮的N2O转化率主要取决于土壤细菌数量,与之成正比.

表3 逐步回归分析结果Table 3 Results of the multiple regressions
3 讨论
本研究发现氮添加显著促进CO2排放的土壤具有较低的黏粒成分、土壤微生物碳和微生物碳氮比,较高的土壤基础呼吸、土壤有效氮及细菌和真菌数量(图4),反映在低MC/MN和高真菌数量上(图5),特别突出.早期研究表明在氮饱和的森林土壤中,氮增加减少了土壤呼吸,在氮限制的土壤中,氮增加对土壤呼吸没有显著影响[8-10].最近的研究则表明氮增加对土壤呼吸的抑制作用主要发生在N缺乏的土壤[26].也有研究发现增氮能增加土壤呼吸[27-28].本研究结果亦是如此,原因在于微生物在分解利用碳氮源时是基于一定的微生物碳氮比进行的,氮对土壤呼吸的影响最终取决于土壤的碳氮比是否适合微生物活动,如果增氮后土壤依旧处于适合微生物的碳氮比,则可能促进土壤呼吸,反之则抑制土壤呼吸.本研究发现外源单位氮的CO2排放因子主要取决于MC/MN,且与MC/MN呈显著负相关(P<0.05),则直接证实了这一点.
无论是否添加氮源,本研究中土壤呼吸均与细菌数量及微生物碳氮呈显著正相关(表2).逐步回归分析则表明土壤细菌和pH均为土壤呼吸的决定因子,且土壤呼吸与pH为负相关关系(表3).对不同利用方式下草地土壤微生物及土壤呼吸特性的既有研究发现,土壤呼吸速率与微生物碳成正相关[29].另有学者对洞庭湖滩地土壤微生物与土壤呼吸特征进行了分析,发现土壤呼吸速率变化与土壤细菌数量变化呈显著相关[30].最新的研究结果表明氮增加对土壤碳动力学的影响机理在于氮诱导酸化导致对pH敏感的微生物及地球化学性质的改变,进而改变碳的矿化与固持[31],也间接证明土壤呼吸与pH的紧密相关性.此外外源单位氮的CO2排放因子与有机碳、全氮、真菌数量呈显著正相关(P<0.05),与MC/MN呈显著负相关(P<0.05;表2).这说明土壤呼吸最终是由碳基质及土壤微生物决定,且细菌和真菌为主要贡献者.
此外,本研究还发现增氮显著促进N2O排放的土壤具有较高的土壤基础呼吸,较低的内源N2O排放和较低的土壤总氮、有效氮、放线菌和真菌数量(图4).尤其是硝态氮和放线菌数量均显著小于增氮未显著影响N2O排放的土壤组(图5).在氮饱和的土壤中,氮添加增加了土壤N2O的排放[9,12].在氮限制的松树土壤中,高氮也增加了土壤N2O的排放[8-10].在温带森林生长季,长期氮沉降降低了72.8%~85.7%的N2O排放通量,而在冻融期增加了11.2~17.8倍N2O排放,这与植物对氮的利用及土壤微生物活性有关[13].但也有研究认为氮沉降无论是对农业土壤还是森林土壤均增加了N2O的排放[15].本研究是纯土壤培养,不存在植物对氮的争夺利用情况,故N2O的排放主要取决于土壤碳氮源和微生物活性.
未添加氮源条件下,即CK的N2O排放与铵态氮含量、有机碳、全氮、有效氮、细菌、放线菌和真菌数量呈显著正相关(P<0.05).添加氮源条件下,即T的土壤N2O排放与土壤基础呼吸和细菌数量呈极显著正相关(P<0.01),与其他因子无明显相关关系(P>0.05,表2).半干旱草地土壤真菌硝化反硝化和细菌自养硝化主导N素转化过程,是N2O产生的主要途径[32].文献[33]等相关研究结果表明米槠天然林土壤在给定实验条件下真菌活性比细菌的大,但二者对土壤N2O产生的贡献几乎相等.本研究也表明放线菌和真菌与N2O的产生有关,但细菌对增氮的响应更明显.逐步回归分析表明未添加氮源处理的土壤N2O排放主要驱动因子是土壤细菌和铵态氮,而添加氮源条件下,主要驱动因子仅是土壤细菌.外源单位氮的N2O转化率也主要取决于土壤细菌数量.本研究土壤的含水量为0.30 g·g-1,土壤处于充氧条件,可能硝化作用占主导地位,铵态氮是硝化反应的底物.在不添加外加氮源的情况下,如果土壤的有效氮含量比较低,那么有效氮含量就会成为限制N2O排放的主要影响因子,而其他土壤因素对N2O排放的影响就很难表现出来[34].本研究结果也说明在没有添加外源氮的情况下,土壤较低的有效氮含量会成为主要限制因子,但当添加外源氮后,底物充足后N2O排放仅取决于微生物的数量.
土壤内源N2O排放和土壤基础呼吸无显著相关关系,但添加外源氮后,N2O排放和土壤呼吸具有极显著正相关关系(P<0.01).这可能是增氮促进了硝化反硝化微生物的活性,此部分微生物的呼吸在土壤总呼吸中占了较高的比重,故N2O排放和土壤呼吸具有极显著正相关关系[35-36].
总之,土壤生化特性尤其是微生物碳氮比、土壤pH及微生物活性在土壤呼吸和N2O排放对氮添加响应方面起了决定性作用.
4 结论
1)氮添加显著促进CO2排放的土壤具有较低的黏粒成分、土壤微生物碳和微生物碳氮比,较高的土壤基础呼吸、土壤有效氮及细菌和真菌数量等特点.无论是否添加氮源,土壤呼吸均主要取决于土壤细菌数量和pH值,与细菌数量呈正相关,与pH呈负相关.外源单位氮的CO2排放量与土壤有机碳、全氮、真菌数量呈显著正相关,与MC/MN呈显著负相关;外源单位氮的CO2排放量主要取决于MC/MN.
2)增氮显著促进N2O排放的土壤具有较高的土壤基础呼吸,较低的内源N2O排放和较低的土壤总氮、有效氮、放线菌和真菌数量等性质.在不添加氮源条件下,N2O排放主要取决于土壤细菌数量和铵态氮含量.添加氮源后土壤N2O排放主要取决于土壤细菌数量;外源单位氮的N2O转化率主要取决于土壤细菌数量,与之呈极显著正相关.
3)土壤内源N2O排放和土壤基础呼吸无显著相关关系,添加外源氮后,N2O排放和土壤呼吸具有极显著正相关关系.
