基于EST-SSR标记的福建云霄县和尤溪县野生茶树遗传多样性分析
2022-04-07杨军孔祥瑞王让剑
杨军,孔祥瑞,王让剑
(福建省农业科学院茶叶研究所/国家茶树改良中心福建分中心/农业部福建茶树及乌龙茶加工科学观测实验站,福建 福州 350013)
福建省内茶树种质资源遗传多样性丰富,存在大量野生茶树种质资源。其中,尤溪野生茶位于福建尤溪县汤川乡赤基村腾峰山苦竹林中,灌木为主,少量小乔木,较直立,花冠白色(少数淡黄色)[1],制成茶形似雀舌,汤色碧绿,香气清醇,滋味甘甜,冲泡五、六次仍清爽可口[2];云霄野生茶位于福建云霄县火田镇大帽山,对云霄县野生茶树子房的茸毛数量、茸毛长度、子房外壁纹饰等调查发现,该地区种质资源子房外壁纹饰以凹陷型为主,存在秃房野生茶种质资源群体[3]。同时,茶树是山茶科山茶属茶组的多年生常绿木本植物[4],且为异花授粉植物,具有高度杂合性,杂交结实率低[5],遗传基础复杂,茶树种质资源传统鉴定方法时间长、鉴定难度大。EST-SSR分子标记在茶树种质资源鉴定与利用方面上有着广泛应用[6-10],尤其在野生茶树种质资源鉴定方面有大量研究。彭靖茹[11]等研究表明广西省9份野生古茶与15份茶树栽培种属于2个类群,亲缘关系较远;陈立杰[12]等基于群体结构分析结果显示,贵阳花溪古茶树资源分为乔木型与灌木型,且由乔木型向灌木型进化;安红卫[13]等认为遗传变异存在于144份贵州茶树种质资源个体间,不同海拔群体间存在较强的基因交流;有研究分别对安徽省太平猴魁茶产区柿大茶群体种[14]、广西省野生茶树种质资源[15]、云南西双版纳茶资源[16]、云南大叶茶种质资源[17]和四川大邑移栽野生大茶树[18]进行分组与遗传多样性分析。因此目前野生大茶树种质资源鉴定主要集中在遗传多样性、资源间亲缘关系与基因交流等方面。本研究对云霄、尤溪茶树种质资源进行分组、亲缘关系与遗传多样性研究,旨在对云霄、尤溪茶树种质资源进行鉴定与筛选,为茶树种质资源保护与利用提供初步依据。
1 材料与方法
1.1 试验材料
参试材料来源于福建省农业科学院茶叶研究所与福建省云霄县、尤溪县,福建省茶树品种资源与云霄、尤溪地方野生茶树种质资源共66个(表1),采集各材料一芽二叶鲜叶后用液氮迅速冷冻处理,保存在-80℃冰箱中备用。
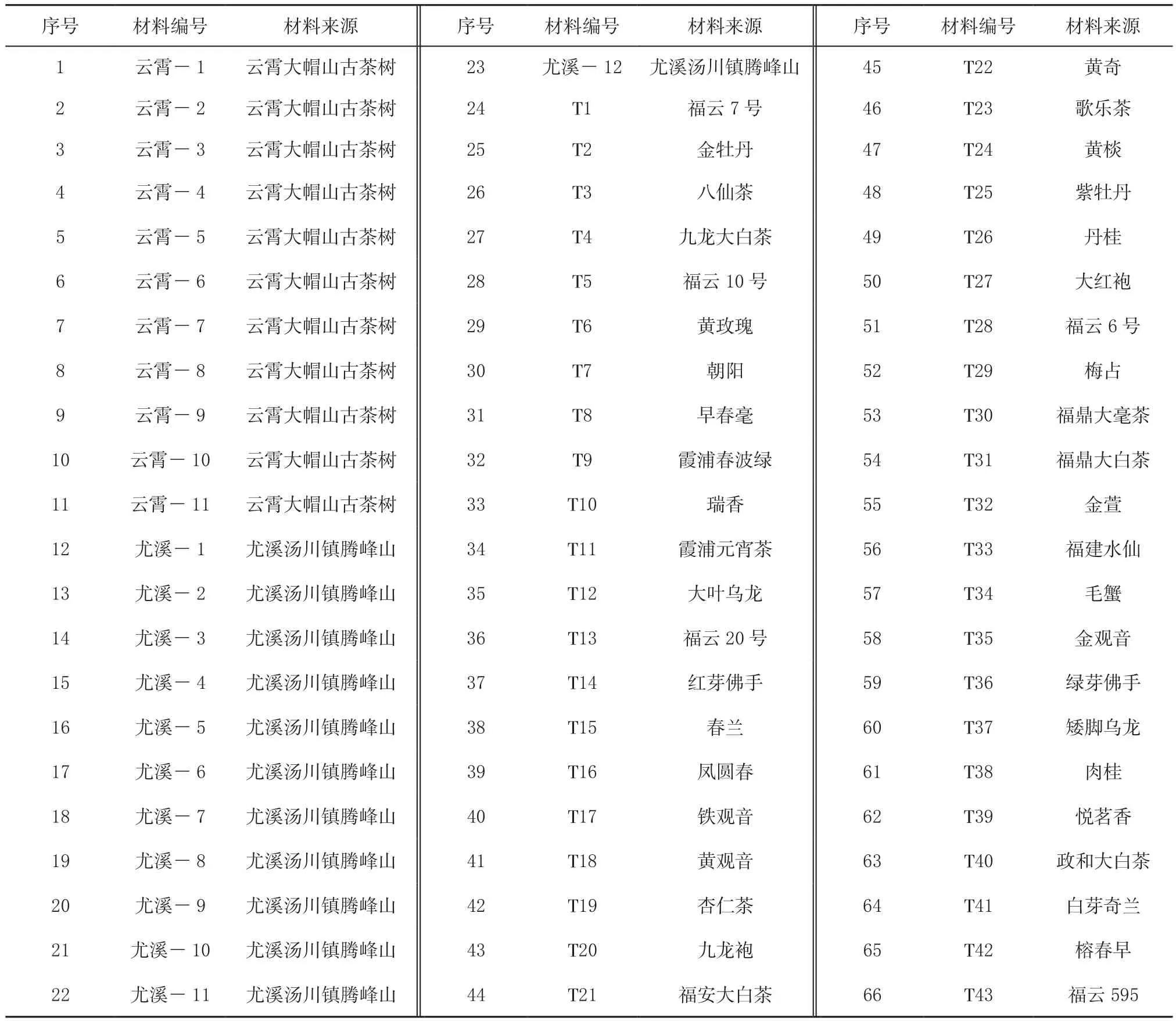
表1 66份供试材料的名称与来源Table 1 Names and origins of 66 specimens collected
1.2 试验方法
1.2.1 农艺性状调查
农艺性状调查参考陈亮等[19]撰写的《茶树种质资源描述规范和数据标准》。
1.2.2 基因组DNA的提取
采用改进的CTAB法[20]提取茶树基因组DNA。用1.0%的琼脂糖凝胶电泳进行茶树基因组分子量大小检测,用756-MC型紫外分光光度计测定茶树基因组DNA的纯度(OD260/280比值)。
1.2.3 引物合成
参照文献[21]引物序列,序列由上海Sangon公司合成,引物信息见表2。

表2 29对SSR引物及其序列Table 2 Nucleotide sequences of 29 primer pairs
1.2.4 PCR扩增和产物鉴定
PCR反应体系为:ddH2O 18.8 μL,10×Buffer(Mg2+)2.5μL,dNTP(10 mmol·L-1)0.5 μL,上、 下 游 primer(10 μmol·L-1) 各 0.5 μL,Taq polymerase(0.5 U)0.2 μL,模板 DNA 2 μL。PCR反应于美国ABI-9600型扩增仪上进行,热循环程序为:94℃预变性4 min,使模板DNA充分变性,然后进入下列温度循环:94℃变性45 s,不同温度条件退火60 s,72℃延伸75 s,重复35个热循环;72℃延伸10 min,最后4℃保存。
引物统一在反向引物(R)的5’段标记荧光(FAM/TAMRA),由上海百力格生物科技有限公司合成。扩增产物 0.5 μL,GeneScan™ 500 ROX™ 0.5 μL,HiDi 9 μL,混匀后使用ABI公司3730XL进行毛细管电泳。
1.3 数据处理
采用ABI公司的GeneMapper4.0软件,选择GeneScan™ 500 ROX™作为分子量标准,对每一个扩增出的条带记录大小。运用PopGen3.2软件计算Shannon信息指数(I)和Nei’s基因多样性指数(H)。运用NTSYSpc2.1中的Tree plot程序进行聚类分析。利用GenAlEx 6.4.1(Peakall and Smouse,2006)进行分子方差分析(analysis of molecular variances,AMOVA)分析。
2 结果与分析
2.1 云霄、尤溪茶树种质资源叶片性状调查
从表3中看能出,云霄、尤溪茶树种质资源在叶面、叶片长宽比性状上差异不大。两地茶树种质资源在叶面性状上皆为微隆起与隆起,叶片长宽比均属于长椭圆形(2.5<长宽比<3.0)。在叶身性状、叶缘性状、叶面积、叶齿数、叶脉对数等方面上有所差异。云霄茶树种质资源叶身多为平与稍背卷,叶缘多为微波与波状,尤溪茶树种质资源多为平与内折,叶缘多为平与微波;云霄茶树种质资源叶面积范围11.29~58.74 cm2,平均值为28.56 cm2,说明云霄茶树种质资源多为中叶种(20 cm2<叶面积<40 cm2),少量种质资源为大叶种;尤溪茶树种质资源叶面积范围18.89~84.01 cm2,平均值为41.94,说明尤溪茶树种质资源多为大叶种(40 cm2<叶面积<60 cm2),少量种质资源为特大叶种;云霄茶树种质资源叶齿数平均值大于尤溪茶树种质资源,叶脉数平均值小于尤溪茶树种质资源。

表3 农艺性状调查结果Table 3 Agronomic characteristics of germplasms

(接表3)
2.2 亲缘关系分析
66个参试材料可以划分为4个类群(图1)。类群A主要为八仙茶、早春毫与云霄地方茶树种质资源,说明云霄地方茶树种质资源与八仙茶、早春毫亲缘关系相对较近;类群B与类群C内的主要茶树品种资源按照不同茶类聚类在一起,类群B主要为福建省乌龙茶品种资源,类群C主要为福建省红、绿茶品种资源,推测茶树品种适制性与茶树品种的亲缘关系相关;类群D皆为尤溪地方茶树种质资源,单独形成一个聚类,说明尤溪地方茶树种质遗传背景比较特殊,属于当地原生茶树种质资源,有待进一步鉴定与挖掘。
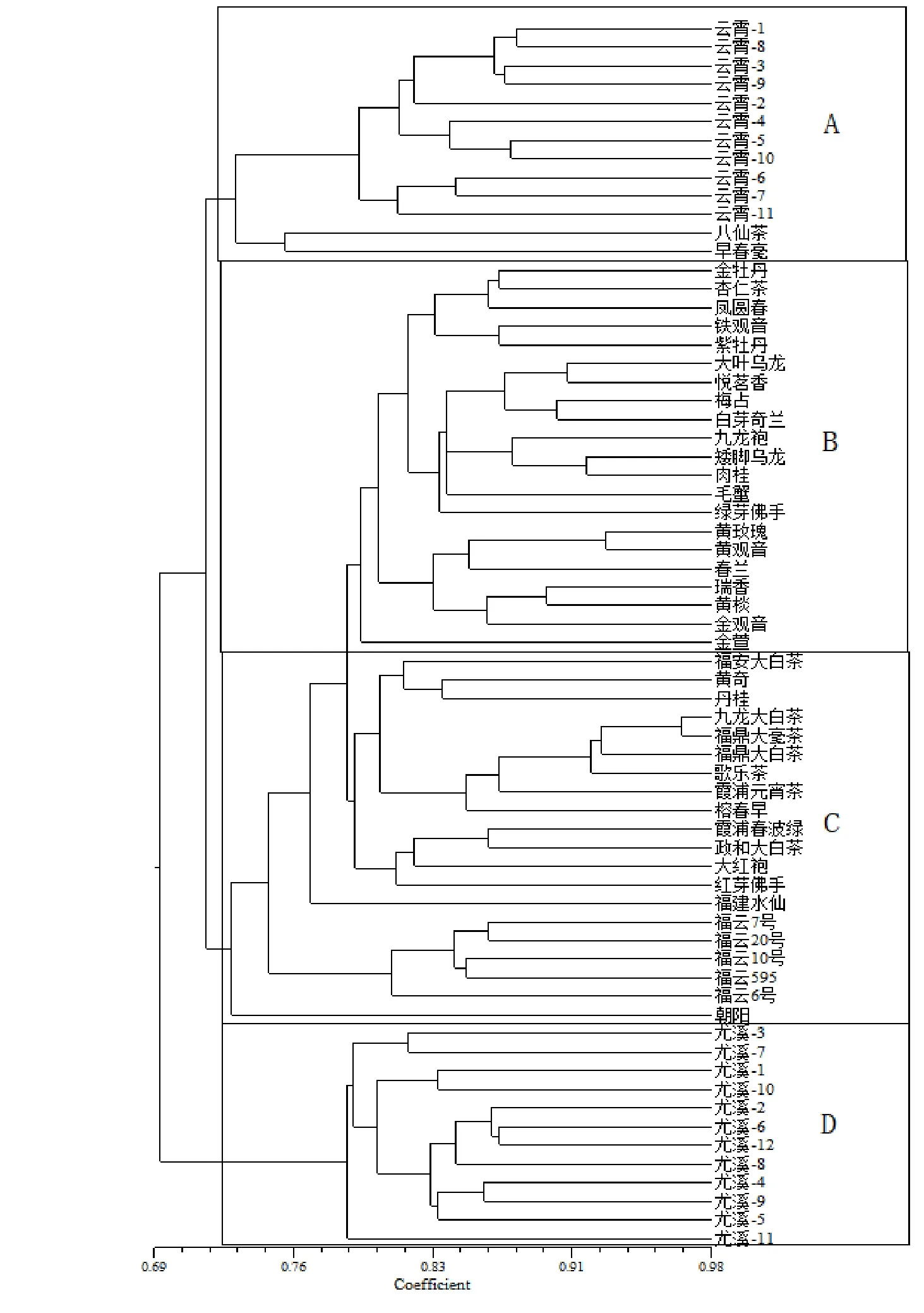
图1 遗传相似系数聚类图Fig. 1 Dendrogram on genetic similarity
2.3 群体遗传多样性分析
从表4中看出,A-D群体中类群C的Shannon遗传多样性指数最高,类群C主要为福建省绿茶品种,其中福云系列茶树品种具有云南大叶种遗传背景,增加类群C的遗传多样性;尤溪、云霄茶树种质资源Shannon遗传多样性指数均高于类群B,类群B主要为福建省乌龙茶品种资源,因此,尤溪、云霄茶树种质资源遗传多样性较为丰富。
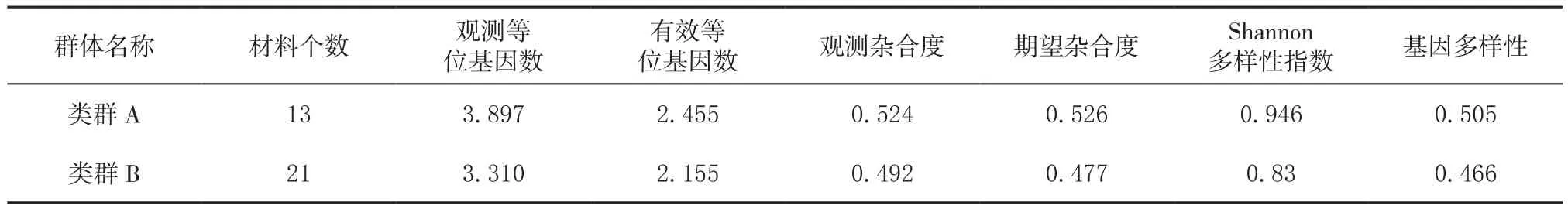
表4 对比4个群体遗传多样性Table 4 Genetic diversity of 4 groups
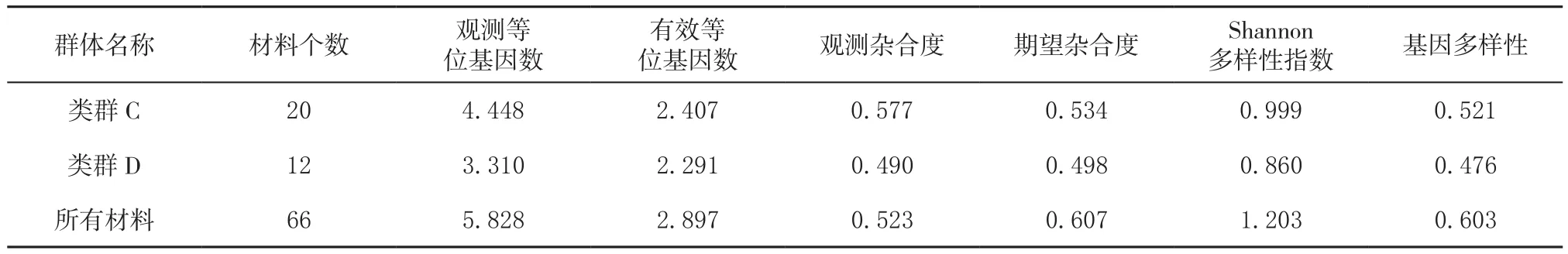
(接表4)
Hardy-Weinberg遗传偏离指数D主要反映Ho(观测杂合度)和He(期望杂合度)之间的平衡关系,D值越接近0,基因型的分布就越接近于平衡状态,D值大于0说明存在杂合子过剩,D值小于0,说明存在杂合子缺失现象[22]。其中,类群A的Ho与对应的He相当,说明类群A基因型的分布就越接近于平衡状态,说明云霄地方茶树种质资源群体相对稳定;类群B、C的Ho大于对应的He,说
明存在杂合子过剩,由类群B、C内主要为福建省茶树品种,品种之间亲缘关系相对较近等原因造成,基因型的分布就未处于平衡状态;类群D的Ho小于对应的He,说明存在杂合子缺失现象,类群D单独为尤溪茶树种质资源,与外来资源遗传交换较少,亲缘关系较远,因此可能存在杂合子缺失。
2.4 群体遗传分化与基因流分析
Wright[23]等认为种群间的遗传分化系数FST值越大,种群的分化程度越高,FST≤0.05时,两群体几乎无分化;0.05<FST≤0.15时,种群中度分化;0.15<FST≤0.25,种群发生明显分化;当FST>0.25时,种群极度分化。FST越接近0,种群间越相似,越接近于1,分化越显著。由表5中可以看出,类群D(主要为尤溪地方茶树种质资源)与类群A、类群B、类群C间的FST值为0.15~0.25,说明尤溪地方茶树种质资源与其它类群发生明显分化;类群B与类群C间的FST为0.589,非常接近0.05,说明类群B与类群C几乎无分化;类群A与类群B、类群C的FST值为0.05~0.15,说明类群A(主要为云霄地方茶树种质资源)与类群B、类群C处于中度分化。
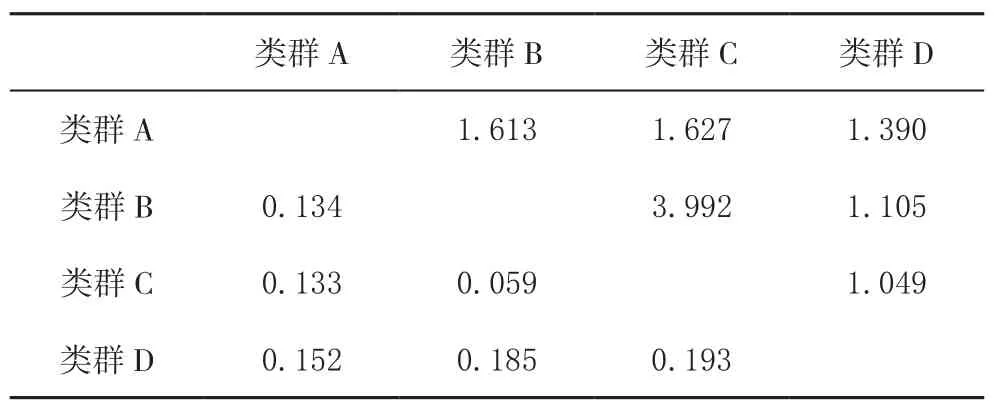
表5 4个类群之间遗传分化系数(对角线以下)和基因流(对角线以上)Table 5 Genetic difference (below diagonal line) and gene flow (above diagonal line) among 4 groups
基因流(Nm)则可衡量群体间基因交流的水平。不同地理种群的基因流用Nm值表示,一般当Nm>4时,表明种群之间的基因流比较充分;若Nm<1,则表明种群可能由于遗传漂变而发生了分化[24]。类群D与类群C的Nm为1.0489,非常接近1,说明类群D与类群C间可能由于遗传漂变而发生了分化;类群B与类群C的Nm为3.9918,非常接近4,表明类群B与类群C间的基因流比较充分。
2.5 不同群体的AMOVA分析
不同群体遗传变异的分子变异分析结果见表6,不同地理种群间的遗传分化(Fst)变异较大(Fst=0.207,P<0.001);不同个体间的遗传分化(Fit)变异较大(Fit=0.220,P<0.001)。78%遗传变异主要发生在个体间,而群体间的遗传变异为21%,1%遗传变异发生在群体内个体间,说明参试材料的遗传变异主要来自于不同群体间和个体间,群体内个体间遗传分化(Fis)变异较小。

表6 群体内和群体间AMOVA分析结果Table 6 AMOVA for intra-and inter-population variations
3 讨论与结论
3.1 云霄、尤溪茶树种质资源亲缘关系分析
本实验以福建省主要栽培品种为参照对云霄、尤溪茶树种质资源进行初步鉴定,鉴定结果显示,福建主要栽培品种分为2个类群,分别主要为类群B(乌龙茶类型群)与类群C(绿茶类型群),而云霄、尤溪茶树种质资源分别形成单一类群,未与参照品种(福建省主栽茶树品种)聚类在一起,亲缘关系较远。其中,云霄茶树种质资源分别与类群B、类群C的遗传分化的系数FST值为0.1342、0.1332,处于中度分化;与八仙茶、早春毫(为广东省凤凰水仙茶树品种的F2代)亲缘关系相对较近,且王泽涵等[3]鉴定显示云霄茶树种质资源中存在子房无茸毛情况与广东省凤凰水仙相似,推测云霄茶树种质资源与广东省凤凰水仙等类型茶树种质资源相似。尤溪茶树种质资源与类群A、类群B与类群C的遗传分化的系数FST值均大于0.15,平均值为0.1765,处于明显分化;与类群B、类群C的Nm分别为1.1049、1.0489,非常接近1,存在遗传漂变分化,基因交流少。同时,群体间的遗传变异为21%,且达到显著性水平,说明云霄、尤溪茶树种质资源与参照品种的遗传差异达到显著性水平,有必要进一步开发其价值。
3.2 云霄、尤溪茶树种质遗传多样性分析
茶树种质资源Shannon多样性指数大小是衡量茶树种质资源遗传多样性的重要指标之一。38个湖南黄金茶Shannon多样性指数为0.55[25];122个云南野生茶树Shannon多样性指数为0.88[26];14份广西野生茶树种质资源Shannon多样性指数0.97[11];广东历史名茶群体种曲江罗坑茶与丰顺马图茶Shannon多样性指数分别为1.22与1.23[27];16个白化、黄化茶树品种Shannon信息指数0.79[28];云霄茶树种质资源Shannon多样性指数0.95,尤溪茶树种质资源Shannon多样性指数0.86。云霄、尤溪茶树种质资源遗传多样性相对较高,且云霄茶树种质资源遗传多样性高于尤溪。
3.3 云霄、尤溪茶树种质基因杂合度分析
基因杂合度是指群体中某个位点上杂合子的频率,反映群体的遗传变异程度,被认为是衡量群体遗传变异的最适参数[29]。本实验参试材料的期望杂合度(0.6072)远高于观测杂合度(0.5231),存在杂合子缺失。其中,类群B与类群C的期望杂合度低于观测杂合度,属于杂合子过剩,可能部分参照品种(如茗科1号、金牡丹等)是通过人工杂交获得,增加类群B与类群C杂合子数量;云霄茶树种质资源的期望杂合度与观测杂合度相当,基因型的分布就越接近于平衡状态;尤溪茶树种质资源期望杂合度高于观测杂合度,与参试材料一样存在杂合子缺失。参试材料的杂合子缺失可能由于云霄、尤溪茶树种质资源与参照品种之间缺少遗传交流,云霄、尤溪茶树种质资源开发利用较少,具有当地资源的独特性。
