小粒种咖啡光强和CO2响应曲线拟合模型筛选
2022-04-05杨雄白学慧马关润周华赵明珠萧自位
杨雄 白学慧 马关润 周华 赵明珠 萧自位


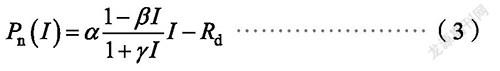
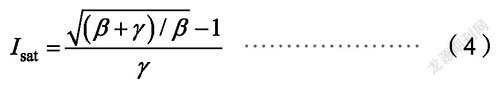

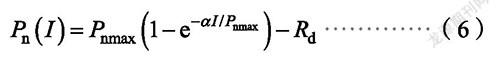

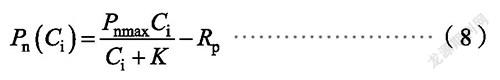

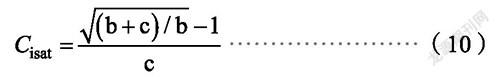
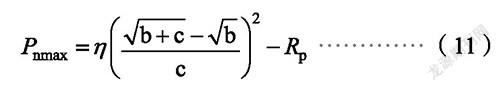






摘要 通過测定不同时间段(上午9:00—11:30和下午15:00—17:30)小粒种咖啡叶片的各项光合响应参数,运用直角双曲线模型、非直角双曲线模型、指数模型、直角双曲线修正模型对小粒种咖啡不同时间段的光响应曲线进行拟合;用直角双曲线模型、Michaelis-Menten模型、直角双曲线修正模型拟合CO响应曲线。本研究旨在选择合适的数学模型拟合小粒种咖啡光响应曲线和CO响应曲线。结果表明:2个时段直角双曲线修正模型的决定系数(R)均在0.960以上,高于其他模型。在上午时段,直角双曲线修正模型拟合的最大净光合速率(P)、光饱和点(I)最接近实测值;下午时段,直角双曲线修正模型拟合的光响应曲线各项参数均与实测值更为相近;采用直角双曲线修正模型拟合上午和下午的CO响应曲线模型的决定系数最高,分别是0.977和0.924,2个时段的最大净光合速率(P)、CO补偿点(Γ)最接近实测值。综合分析,直角双曲线修正模型较适合用于拟合小粒种咖啡的光响应曲线和CO响应曲线。
关键词 小粒种咖啡;光合作用;光响应曲线;CO响应曲线
中图分类号 S533 文献标识码 A DOI:10.12008/j.issn.1009-2196.2022.02.012
Selection of Models of Light Response and CO Response Curves for Coffea arabica
YANG Xiong1 BAI Xuehui2 MA Guanrun2 ZHOU Hua2 ZHAO Mingzhu2 XIAO Ziwei2
(1. College of Tropical Crops,Yunnan Agricultural University,Pu'er,Yunnan 665099,China;2. Yunnan Dehong Research Institute of Tropical Agricultural Sciences,Ruili,Yunnan 678600,China)
Abstract Parameters of photosynthesis response of Coffea arabica leaves were measured at different time intervals (9:00-11:30 AM and 15:00-17:30 PM). The light response curves at different time intervals were fitted with rectangular hyperbola model,non-rectangular hyperbola model,exponential equation model and modified rectangular hyperbola model,and the CO response curves were fitted with the rectangular hyperbola model,the Michaelis-Menten model and the modified rectangular hyperbola model in a view to selecting an optimal mathematical model to fit the light response curve and CO response curve for the leaves of C. arabica. The results showed that the determination coefficients (R)of the two modified rectangular hyperbola models are above 0.960,which are higher than those of the other models. In the morning,the maximum net photosynthetic rate (P)and light saturation point (I)fitted by the modified rectangular hyperbola model are the closest to the measured values. In the afternoon,all the parameters of the light response curve fitted by the modified rectangular hyperbola model are more similar to the measured values. The maximum net photosynthetic rate (P)and CO compensation point (Γ)fitted by the modified rectangular hyperbola model in the morning and the afternoon are 0.977 and 0.924,respectively,which are closest to the measured values. In conclusion,the modified rectangular hyperbola model is more suitable for fitting the light response curve and the CO response curve of the leaves of C. arabica.
Keywords Coffea arabica;photosynthesis;photosynthesis-light response curve;CO response curve
小粒种咖啡原产于非洲,是一种常绿小乔木[1],适宜生长在静风、温凉、湿润的环境中[2-3]。咖啡叶片的光合能力不仅影响植株生长发育,同时也是影响咖啡产量及品质的重要因素[4]。光照强度对咖啡叶片光合速率影响较大。研究表明,不同光照环境下咖啡叶片的光合响应参数各不相同[5-6]。在研究光照强度对植物叶片光合特征的影响时,通过拟合植物叶片光合响应曲线来反映叶片的各项光合响应参数是一个有效的方法,利用这些参数有助于判定叶片光合机构的运转状况和叶片利用光、CO等资源的能力[7-8]。目前用于拟合植物光响应曲线方面的模型主要有直角双曲线模型、非直角双曲线模型、指数模型和直角双曲线修正模型[9],这些模型在拟合植物光响应曲线时各有优缺点,同时由于不同植物对光的响应存在着生理生化上的差异[7],其适宜的光合响应模型可能各不相同因此,对特定植物选择最适宜的光合响应模型才能更好地反映叶片的光合响应特征。
CO不仅是光合作用的底物,而且是C植物光合作用的主要限制因子[10].要深入了解CO对植物光合作用的影响,就必须知道与之相关的CO饱和点(C)、最大净光合速率(P)、CO补偿点(Γ)、光呼吸速率(R)和初始竣化效率(η)等重要光合参数[11-12]。目前,测定CO响应曲线是获得这些参数的主要手段[13-14]。在过去,国内外学者已建立多个CO响应曲线的数学模型,其中常用的模型有直角双曲线模型、Michaelis-Menten模型、直角双曲线修正模型[13]。这些模型虽已广泛应用于定量研究植物对CO浓度升高造成的生理生态响应[13],但由于不同植物对CO浓度变化的敏感度及响应方式存在较大差异,因此,不同植物的最适CO响应曲线模型也各不相同[12]。选择最优的模型拟合CO响应曲线,有助于更准确深入地了解植物光合对CO浓度变化的响应机制。
虽然国内外学者已对小粒种咖啡叶片的光合作用做了许多研究[4,15],但未见有关小粒种咖啡光合-光响应模型和光合-CO响应模型研究的报道。为此,本研究拟通过测定相同环境下小粒种咖啡树2个时段叶片的各项光合指标,用不同的光合-光响应模型和光合-CO响应模型进行拟合分析,选择出最佳拟合模型,为深入了解小粒种咖啡的光合生理生态特征,科学指导小粒种咖啡在云南热区栽培提供参考,同时也为不同光合- 光响应模型和光合-CO响应模型适用性评价提供试验对比。
1 材料及方法
1.1 材料
1.1.1 ; 样地概况 试验地位于云南省德宏热带农业科学研究所的咖啡生产基地内(24°1′12.88″N,97°51′21.46″E),属南亚热带季风气候,年平均气温约21℃,年降雨量约1 390 mm,分干湿两季,雨季降雨量占全年降雨量的80%以上。试验地土壤含水量约21%,容重约1.23 g/cm。
1.1.2 试材 试验材料为小粒种咖啡卡蒂姆品系,2017年定植,按生产种植常规管理,植株生长良好。
1.2 方法
1.2.1 试验设计 2020年9—12月,选择20棵长势一致的咖啡树,随机分为2组,每组10棵。采用美国PP-Systems公司生产的CIRAS-3便携式光合作用仪测定系统对所选叶片进行测定。测定的时间分为2段:上午(9:00—11:30)和下午(15:00—17:30)。测定时设定叶室面积25 mm×7 mm,用CO钢瓶供气,将叶温设置在27℃。
1.2.2 光合-光响应曲线测定 选取植株中上部生长良好的一分枝,以一分枝条顶端以下的第三对或第四对叶片作为测量光响应曲线的材料。测定净光合速率P(I)过程中,大气相对湿度设置为50%~60%,将叶温设置为27℃,CO浓度设为390 μmol/(m·s),选用光合仪内置光源将光合有效辐射(I)设为0、50、100、150、200、250、300、350、400、500、600、700、800、1 000、1 200 μmol/(m·s),共15个梯度。每个光照强度等待120 s后开始测定,每个梯度连续重复测定3次,每次重复间隔10 s。
1.2.3 光合-CO响应曲线测定 光响应曲线测定完后,对同一叶片的光合-CO响应曲线进行测定,测定净光合速率P(C)过程中,大气相对湿度设置为50%~60%,光合有效辐射(I)设定为700 μmol/(m·s),CO濃度梯度为350、250、150、50、150、250、350、450、550、650、750、850、950、1 050、1 250、1 450、1 650 μmol/(m·s),共17个梯度。稳定时间和数据记录与测定光响应曲线时相同。
1.2.4 曲线拟合方法
1.2.4.1 光响应曲线 直角双曲线模型数学表达式
式中,P(I)为净光合速率,I为光强,α为植物光合作用对光响应曲线在I-0时的斜率,即光响应曲线的初始斜率,也称为初始量子效率,P为最大净光合速率,R为暗呼吸速率。
非直角双曲线模型数学表达式
式中,P(I)为净光合速率,I为光强,θ为曲线的曲率,α为光响应曲线的初始斜率,P为最大净光合速率,R为暗呼吸速率。
直角双曲线的修正模型
式中,α是光响应曲线的初始斜率,β和γ为系数,I为光合有效辐射,R为暗呼吸速率。饱和光强I为:
最大净光合速率用P为:
指数模型数学表达式
式中,P(I)、α、P、R和I的定义与前述相同。
1.2.4.2 光合-CO响应曲线 直角双曲线模型数学表达式
式中,P(C)为净光合速率,C为胞间CO浓度,η为CO响应曲线的初始斜率,也称为初始竣化效率,P为最大净光合速率,R为光呼吸速率。
Michaelis-Menten模型数学表达式
式中,P(C)、C、P和R的定义与前述相同,K为Michaelis-Menten常数。
直角双曲线修正模型数学表达式
式中,P(C)、C、η和R的定义与前述相同,b和c为系数。植物的饱和胞间CO浓度C为:
植物的最大净光合速率用P为:
1.2.5 光和-CO饱和点的计算由于直角双曲线、非直角双曲线和指数函数模型3种模型曲线是一条没有极点的渐近线,因此无法求出光饱和点(LSP),所以本文参照前人的研究[16-17],在弱光强条件下(≤200 μmol/(m·s)利用公式P(I)=αQ-R(12)拟合直角双曲线、非直角双曲线的光饱和点(LSP ),当P=P时的Q即为光饱和点(LSP);在低CO浓度条件下[≤200 μmol/(m·s)]的CO响应数据,利用公式P(C)=ηQ-R(13)拟合直角双曲线、Michaelis-Menten模型的CO饱和点(CSP),当P=P时的Q即为CO饱和点(CSP)。对于指数模型,需假设光合速率为0.999 P所对应的光强为饱和光强[18]。
1.2.6 数据处理 通过均方根误差(RMSE)和决定系數(R)检验各模型的拟合精度。
式中,P为预测值,Q为实测值,n为样本个数。RMSE值越小,表明预测值与实测值越接近。
利用叶子飘等的光合计算软件对各种响应曲线进行拟合,得出相应的响应参数。用Microsoft Excel对数据进行整理,采用SPSS进行数据分析。对测定的光合参数进行正态性检验,如果服从正态分布,则对数据进行方差分析;如果不满足正态分布,则选用2个独立样本的非参数检验中的Mann-Whitney检验。
2 结果与分析
2.1 光合-光响应曲线
运用直角双曲线模型、非直角双曲线模型、直角双曲线修正模型、指数模型对实测值进行拟合的曲线如图1所示,各拟合参数见表l。结合图1和表1可知,采用直角双曲线修正模型在2个时段拟合的曲线趋势与实测结果最为接近,其上午和下午的模型决定系数(R)分别0.981和0.960,均方根误差(RMSE)分别为0.144和0.129,决定系数(R)均高于其它模型,而均方根误差(RMSE)最小,表明该模型拟合效果最好。而直角双曲线模型拟合出的曲线和实测值差异最大,非直角双曲线模型和指数模型所拟合的曲线和实测值均有一定的差异。
上午直角双曲线修正模型拟合的最大净光合速率(P)最接近实测值,仅比实测值低0.009。下午非直角双曲线模型拟合的最大净光合速率(P)最接近实测值,比实测值小0.016,其次是直角双曲线修正模型拟合的最大净光合速率(P),比实测值小0.372。与直角双曲线模型、非直角双曲线模型和指数模型相比,直角双曲线修正模型拟合的光饱和点(I),无论是上午还是下午都最接近实测值,分别比实际值大36.278和68.491 μmol/(m·s)。在上午,直角双曲线模型拟合的光补偿点(I),最接近实测值,仅比实测值大1.141 μmol/(m·s),下午也是直角双曲线修正模型的拟合值最接近实测值,相差3.697 μmol/(m·s)。在暗呼吸速率(R)方面,上午和下午非直角双曲线模型拟合的暗呼吸速率(R)与实测值最接近,分别与实测值相差0.081和0.067 μmol/(m·s)。4个模型的初始量子效率(a)均在0~0.125的理论范围,且直角双曲线模型在2个不同时间段的初始量子效率(a)均比其他几个模型的大,分别为0.080和0.085,其余3个拟合模型的初始量子效率(a)变化不大,在0.017~0.045。
对上午、下午用最优直角双曲线修正模型拟合的数据进行分析比较,2个时段的拟合参数比较见表1 “[]”中,咖啡叶片上午的最大光合速率[P=7.991 μmol/(m·s)]显著高于下午的最大光合速率[P=3.628μmol/(m·s)](p<0.05),而上午和下午的光饱和点(I)、光补偿点(I)和暗呼吸速率(R)和初始量子效率(a)无显著差异(p>O.O5)。虽然上午和下午的光饱和点(I)无显著差异,但上午[I= 636.278μmol/(m·s)]高于下午[I= 558.491 μmol/(m·s)]。由图2可看出,光合有效辐射(I)在0~500 μmol/(m·s)时,净光合速率(P)迅速上升;当光合有效辐射(I)在500~700 μmol/(m·s)时,净光合速率(P)升幅减弱;当光合有效辐射(I)大于700 μmol/(m·s)时,净光合速率(P)随着光合有效辐射(I)的增加而出现下降的趋势,表现出一定的光抑制现象。与上午不同的是,到了下午,光合有效辐射(I)在0~400 μmol/(m·s)时,净光合速率(P)迅速上升;当光合有效辐射(I)在400~600 μmol/(m·s)时,净光合速率(P)升幅趋于平缓,光抑制现象同样出现在光合有效辐射(I)大于700 μmol/(m·s)的时候。
2.2 光合-CO响应曲线
分别采用直角双曲线模型、Michaelis- Menten模型、直角双曲线修正模型对CO响应曲线进行拟合,拟合曲线模型如图2所示,各拟合曲线模型参数见表2。由表2可知,2个时间段3个拟合模型的决定系数(R)和均方根误差(RMSE)相差不大,但直角双曲线修正模型的决定系数(R)最高,均方根误差(RMSE)最低,拟合精度稍高一些。
由表2可知,2个时间段直角双曲线修正模型拟合的最大净光合速率(P)最接近实测值,仅相差0.007和0.599 μmol/(m·s),其余2个拟合模型拟合的最大净光合速率(P)均约为实测值的2倍。虽然2个时段直角双曲线修正模型拟合的CO饱和点(C)与实测值最为接近,但还是比实测值分别小94.247和132.222 μmol/(m·s)。3种拟合模型拟合的CO补偿点(Γ)均和实测值无显著差异,2个时段直角双曲线修正模型的拟合值分别比实测值大0.245、0.162μmol/(m·s),上午和下午直角双曲线修正模型拟合的光呼吸速率(R)最接近实测值,分别比实测值大0.780、0.267μmol/(m·s)。直角雙曲线模型和直角双曲线修正模型的初始竣化效率(η)均在0~0.200的理论范围[12],直角双曲线的初始竣化效率(η)最大,上午和下午分别为0.063和0.058。由图2可知,2个不同时间段直角双曲线修正模型的拟合曲线最佳,并且是3种模型中唯一能够呈现出CO 浓度达到CO饱和点(C)后净光合速率(P)随CO浓度升高而降低的现象,直角双曲线模型和Michaelis-Menten模型的拟合曲线和实测值略微有一点差异。
选用直角双曲线修正模型拟合上午和下午的CO响应曲线,见表2“[]”中方差分析结果。上午的最大净光合速率(P=18.007 μmol/(m·s))显著高于下午的最大净光合速率(P=13.401 μmol/(m·s))(p<0.05),而CO饱和点、CO补偿点(Γ)和光呼吸速率(Rp)以及初始竣化效率(η)没有显著差异(p>0.05)。从图2可以看出,在上午CO浓度达到750μmol/(m·s)前,随着CO浓度的增加,净光合速率(P)迅速增加;CO浓度达到750 μmol/(m·s)之后,净光合速率(P)随着CO浓度的增加而缓慢上升。而在下午,CO浓度达到650 μmol/(m·s)前,随着CO浓度的增加,净光合速率(P)迅速增加;CO浓度在650~1 450 μmol/(m·s)时,净光合速率(P)随着CO浓度的增加而缓慢上升;CO浓度达到1 450 μmol/(m·s)之后,随着CO浓度的增加,净光合速率(P)开始呈现出缓慢下降的趋势。
3 讨论与结论
叶子飘等[13,18]研究发现,对于光合-光响应拟合模型来说由于初始量子效率(a)和最大净光合速率(P)均大于0,所以直角双曲线模型和指数模型是一条没有极点的渐近线,非直角双曲线模型是直角双曲线模型的一种特殊情况(θ≠0),即3种模型均为没有极点的渐近线,所以不能直接求出光饱和点(I)和最大净光合速率(P);对于直角双曲线模型和非直角双曲线模型来说,只能通过最小二乘法估算光饱和点(I)和最大净光合速率(P),而该方法的缺点是会使估算出来的最大净光合速率(P)远大于实测值,光饱和点(I)远小于实测值;对于指数模型,则需假设光合速率为0.999 P所对应的光强估算饱和光强,也会与实测值有一定的差异。由3种拟合模型的公式可知,即使光合有效辐射(I)超过植物的光饱和点,净光合速率(P)也依然会随着其升高而增大,所以这3种拟合模型无法拟合出超过光饱和点(I)后的净光合速率(P)随光合有效辐射(I)升高而降低的过程;而直角双曲线修正模型与其他3个拟合模型的最主要区别就是有极点,即可以直接表示出最大净光合速率(P)及光饱和点(I),且与实测值最接近,也能表示出超过光饱和点(I)后净光合速率随(P)光合有效辐射(I)升高而降低的过程,贺正等[19]、王海珍等[20]的研究结果也验证了这一观点。同理,对于光合-CO拟合模型来说,直角双曲线模型是没有极点的渐近线,而M-M模型在本质上与直角双曲线模型完全相同,即只有直角双曲线修正模型可以直接得出CO饱和点和最大净光合速率,其余2个拟合模型估算的拟合值与实测值存在很大的差异,且无法表现出过CO饱和点(C)后,随着CO浓度的升高,净光合速率(P)出现一定程度的下降的过程。
本试验光合-光响应曲线可以分为3个阶段,第一阶段为线性阶段,净光合速率(P)随光合有效辐射(I)的增加呈直线上升;第二阶段,随着光合有效辐射(I)的增加,净光合速率(P)的增幅减弱,直至到达光饱和点(I),出现最大净光合速率(P);第三阶段,随着光合有效辐射(I)增加,净光合速率(P)逐渐减弱,出现一定的光抑制现象,这与李虎军等[21]的研究基本相同。与光合-光响应曲线类似,光合-CO响应曲线也可分为3个阶段,第一阶段,随着CO浓度升高,净光合速率(P)呈线性增加;第二阶段,随着CO浓度的升高,净光合速率(P)增幅减弱,直到出现CO饱和点(C);第三阶段,超过CO饱和点(C)后,随着CO浓度的升高,净光合速率(P)出现一定程度的下降。试验发现,光合-光响应曲线和光合-CO响应曲线小粒种咖啡上午的光合能力(P)均强于下午,这表明二者光合能力存在差异。胡发广等[22]研究发现,咖啡叶片的光合速率日变化呈双峰曲线,下午净光合速率峰值仍低于上午,而正午由于阳强、温度高而出现午休现象。光合作用是一种酶促反应[23],而温度对酶活性影响很大,温度过高会抑制其酶的活性,进而降低其光合能力;此外,下午也比上午湿度小,低湿环境下,植物为了防止自身水分过度的流失,会关闭一部分气孔,使得进入植株的CO减少,植物的光合作用也会受到影响[24]。
由最优模型拟合所得的上午咖啡叶片的初始量子效率(a)为0.039,下午的初始表光效率(a)為0.029,均低于正常环境下的植物初始量子效率(a),且上午和下午的值并无显著差异,造成这一结果的原因有待进一步从小粒种咖啡的适应性角度出发进行研究。最优模型拟合所得的上午的初始竣化效率(η)为0.046,下午的初始竣化效率(η)为0.035,二者都在0~0.200的理论范围[12],说明模型拟合能够在很大程度上真实反映小粒种咖啡的初始竣化效率(η)。
通过各项参数综合分析,采用直角双曲线修正模型拟合的光合-光响应曲线和光合-CO响应曲线效果最佳。上午叶片的最大净光合速率(P)、光饱和点(I)、CO饱和点(C)都高于下午,表明小粒种咖啡在上午时段具有更强的光合速率。因此,在生产实践中,应注意采用“早向阳,晚遮阴”的园区设计原则,以提高小粒种咖啡叶片的光合能力,达到增产、稳产的效果。
参考文献
[1]董云萍,黎秀元,闫林,等.不同种植模式咖啡生长特性与经济效益比较[J].热带农业科学,2011,31(12):12-15+32.
[2]龙乙明,王剑文.云南小粒种咖啡栽培技术[M].昆明:云南科技出版社,2009.
[3]罗星明.云南热区咖啡种植气候条件的模糊区域性划分[J].热带农业科技,1994,17(4):20-24.
[4] Damatta F M. Ecophysiological constraints on the production of shaded and unshaded coffee:a review[J]. Field Crops Research,2004,86:99-114.
[5] Cavatte P C,Oliveira ÁA G,Morais L E,et al. Could shading reduce the negative impacts of drought on coffee?A morphophysiological analysis[J]. Physiologia Plantarum,2012,144:111-122.
[6] Neto F J D S,Bonfanti L,Gazaffi R,et al. Effects of shade trees spatial distribution and species on photosynthetic rate of coffee trees[J]. CoffeeScience,2019,14(3):326-337.
[7]张赞齐,高世伦,卫星杓,等.光合模型对无患子叶片光合响应参数计算结果的影响[J].北京林业大学学报,2019,41(4):32-40.
[8]高照全,冯社章,张显川,等.不同辐射条件下苹果叶片净光合速率模拟[J].生态学报,2012,32(4):1 037-1 044.
[9]梁星云,刘世荣.基于冠层塔吊原位测定长白山温带阔叶红松原始林群落主要树种的光合特征[J].应用生态学报,2019,30(5):1 494-1 502.
[10]泰兹,奇格尔.植物生理学[M].北京:科学出版社,2009.
[11]康华靖,陶月良,权伟,等.植物光合CO响应模型对光下(暗)呼吸速率拟合的探讨[J].植物生态学报,2014,38(12):1 356-1 363.
[12]李思嘉,陈志德,王晓婧,等.花生不同光合-CO响应曲线拟合模型的比较[J].中国农学通报,2020,36(12):33-38.
[13]叶子飘.光合作用对光和CO响应模型的研究进展[J].植物生态学报,2010,34(6):727-740.
[14]李丽霞,刘济明,黄小龙,等.不同氮素条件米槁幼苗光合作用对CO响应特征[J].东北农业大学学报,2017,48(2):29-36.
[15] Damatta F M,Ramalho J. Impact of drought and temperature stress on coffee physiology and production:a review[J]. Brazilian Journal of Plant Physiology,2006,18(1):55-81.
[16]侯维海,王建林,旦巴,等.青棵光合作用5种光响应模型的比较分析[J].作物杂志,2017(4):96-104.
[17]刘洪波,白云岗,张江辉,等.葡萄光合速率对光及CO浓度的响应特征[J].黑龙江农业科学,2015(7):77-80.
[18]叶子飘,王建林.植物光合-光响应模型的比较分析[J]. 井冈山大学学报,2009,30(4):9-13.
[19]賀正,刘志,苗芳芳,等.宁夏引黄灌区滴灌玉米4种光响应曲线模型比较[J].玉米科学,2020,28(1):124-131.
[20]王海珍,韩路,徐雅丽,等.干旱胁迫下胡杨光合光响应过程模拟与模型比较[J].生态学报,2017,37(7):2 315- 2 324.
[21]李虎军,王全九,李苏君,等.红提葡萄光合速率和气孔导度的光响应特征[J].干旱地区农业研究,2017,35(4):230-235+262.
[22]胡发广,刘红明,毕晓菲,等.云南干热河谷区7个小粒咖啡叶片光合特性日变化的研究[J].江西农业学报,2020,32(10):52-56.
[23]王为民,王晨,李春俭,等.大气二氧化碳浓度升高对植物生长的影响[J].西北植物学报,2000,20(4):676-683.
[24] FarquharGD,Sharkey T D. Stomatal conductance and photosyntliesis[J]. Annu Rev Plant Physiol,1982,33(1):317-345.
