贵州雷公山水青冈群落特征与分布格局研究
2022-04-01杨礼旦陈应强
杨礼旦,陈应强
(贵州省台江县林业局,贵州 台江 556300)
壳斗科(Fagaceae)水青冈属(Fagus)是经过第三纪残留的古老高大乔木,属于欧亚-北美间断分布类型,是北半球温带森林植被的重要建群种,间断分布于北半球亚热带山地[1—2]。水青冈(Fagus longipetiolata)群落是中亚热带中山、亚高山的一种地带性群落,常在其分布的垂直林带下部与组成纬度地带性常绿阔叶林上限的常绿阔叶树种组成混交林,这种现象为中国所独有,具有重要的科学价值[3]。水青冈在我国分布于秦岭以南、五岭南坡以北各地,生于海拔300~2400 m 山地杂木林中。目前对水青冈属其他种的群落研究较多,如亮叶水青冈(Fagus lucida)[5—6],对水青冈的研究则相对较少[7—11]。高香琴等[7—8]对梵净山水青冈属群落木本植物组成特征进行分析,认为梵净山同时拥有3 种水青冈属植物,其群落木本植物物种组成丰富,区系复杂、古老、热带性质明显,这在全世界范围内是独一无二的,具有重要的世界遗产价值;赵冬林等[9]对云南九龙山水青冈群落的物种组成、重要值、物种多样性、种群年龄结构进行研究,表明种群自然更新不良,为衰退型群落;李登江[10]等从群落的物种组成、重要值、物种多样性等方面对雷公山野猪塘水青冈群落进行分析,认为该群落中水青冈占绝对优势地位,为建群种;杨礼旦等[11]初步分析了雷公山水青冈群落的物种多样性以及水青冈和青冈栎(Cyclobalanopsis glauca)两个乔木种群分布格局,认为水青冈群落乔木层、灌木层物种多样性均较高,草本层多样性偏低,群落的物种多样性较高,种群分布格局随径级增大,水青冈种群由随机分布向集群分布过渡,而青冈栎种群则由集群分布向随机分布过渡。
水青冈群落是雷公山地带性群落,分布在海拔800~1800 m 山地杂木林中,形成原生性群落。水青冈树体冠幅大,对保持水土至关重要,在该地区生态安全维护中有着极大的作用[4]。此前对贵州雷公山水青冈群落特征[10]和种群空间分布格局[11]的研究多是局部的,设置样地过少,难以完全反映雷公山水青冈群落特征和空间分布格局,对其深入研究可为揭示该地区群落的稳定与物种多样性的维持机制提供参考。同时,水青冈作为研究植物系统演化和植物地理学的关键类群之一[6],对其深入研究具有重要意义。本研究旨在通过研究雷公山水青冈群落年龄结构、空间分布格局、生物多样性,揭示该群落动态特征,预测其未来演变趋势,以期建立水青冈群落的适应性保护措施。
1 研究区域概况
研究地位于贵州省雷公山国家级自然保护区和台江县南宫州级自然保护区内(26°24′~26°38′N,108°16′~108°29′E)雷公山主体山脉,处于长江水系与珠江水系的分水岭地带,是清水江和都柳江主要支流的发源地和水源补给区。该区地貌属侵蚀低中山地形,河流侵蚀切割强烈,地形破碎。该区水资源十分丰富,气候温暖湿润,年均温15 ℃,年均积温3500~4500 ℃,年均降水量1450 mm。土壤为山地黄壤,土层较薄,但有机质、全氮、全钾含量高,土质疏松,透气性好[12]。该区原生植被为中亚热带湿润常绿阔叶林,有原生性较好的森林植被,森林覆盖率86%。调查区人为活动少,植被未受到较强的人为干扰。
2 研究方法
2.1 野外调查
在踏查的基础上,参考有关文献[5—11],选择水青冈分布地段设置面积30 m × 20 m标准样地11 个,每个样地的基本概况如表1。将标准样地划分为6个10 m × 10 m 样方,在西南角顶点设置样桩,然后将6 个样方分别划分为4 个5 m × 5 m 的小样方,以该小样方作为外业的基本单位。调查乔木层(高度≥5 m 起测)种类,记录树种、株数、树高、胸径、枝下高、冠幅、生活力等;选取每个西北角的小样方(5 m × 5 m)作为灌木调查单元,调查灌木树种、株丛数、地径、高度、生长状况、分布状况和盖度等;在灌木样方的西北角设置1 m × 1 m 样方作为草本植物观测区,调查草本植物种类、株数、冠径、高度、生长情况、盖度等。

表1 雷公山水青冈群落样地基本信息Table 1 Basic information of Fagus longipetiolata community plots in Leigong Mountain
2.2 数据处理
2.2.1 重要值计算
统计群落的物种并分别计算样地内乔木层、灌木层、草本层各物种的重要值和物种多样性。重要值计算参照张金屯[13]的计算方法,即重要值(IV)=(相对密度+相对优势度+相对频度)/3。
2.2.2 龄级、径级结构划分标准
径级按胸径(DBH)分为5 个立木等级[14]:Ⅰ级(DBH<5 cm)(含幼苗DBH<5 cm、H<0.5 m 和幼树DBH<5 cm、H>0.5 m),Ⅱ级(5 cm≤DBH<10 cm),Ⅲ级(10 cm≤DBH<20 cm),Ⅳ级(20 cm≤DBH<30 cm)和Ⅴ级(30 cm≤DBH)。由于径级立木划分标准无法准确反映乔木层的垂直结构,故同时采用高度级分析,以植株高度(H)每3 m 为一个等级 (Ⅰ级:0<H≤3 m;Ⅱ级:3 m<H≤6 m;Ⅲ级:6 m<H≤9 m;Ⅳ级:9 m<H≤12 m;Ⅴ级:12 m<H≤15 m;Ⅵ级:15 m<H≤18 m;Ⅶ级:18 m<H≤21 m;Ⅷ级:21 m<H≤24 m;Ⅸ级:H>24 m)[15]。
2.2.3 物种多样性计算
多样性计算公式如下:
Gleason 丰富度指数(E):
Gimpson 多样性指数(D):
Shannon-Wiener 多样性指数(H):
Pielou 均匀度指数(Jsw):
式中,S为物种数目,A为单位面积,Ni为第i个物种的个体数,N为所有种的个体总数,Pi为频度,即Pi=Ni/N。
2.2.4 空间分布格局
使用方差比均值(V/M)的t检验法分析不同发育阶段的种群空间分布格局,并使用丛生指数(I)、平均拥挤度(M*)、Cassie 指数(CA)、Lloyd 聚块性指数(m*/m)、负二项式参数(K)共5 种指标判断其聚集强度[16—17]。
2.2.5 数据处理
采用SPSS 25.0 软件进行数理统计和分析。
3 结果与分析
3.1 群落物种组成
经调查,水青冈群落有维管植物50 科77 属103 种(含种下等级,下同),其中蕨类植物3 科3属3 种,种子植物47 科74 属100 种,包括裸子植物3 科3 属3 种,被子植物44 科71 属97 种。以壳斗科5 属7 种、樟科(Lauraceae) 4 属7 种、蔷薇科(Rosaceae) 5 属7 种、杜鹃花科(Ericaceae) 2属5 种为主,这4 科包含了16 属26 种,分别占总属数的20.78%、总种数的25.24%,为群落优势科。从表1 可见,含1 种的科有27 科,占总科数的54.00%;含1 种的属有58 属,占总属数的75.32%,群落以单种科、单种属占优势。总的来看,水青冈群落物种组成较复杂。
水青冈群落乔木层物种共有42 种,占总种数的40.78%,以壳斗科最多。乔木层建群种有水青冈、青冈(Cyclobalanopsis glauca)、西南木荷(Schima wallichii)、云贵鹅耳枥(Carpinus pubescens)、多脉青冈(Cyclobalanopsis multinervis),其次有木荷(Schima superba)、甜槠(Castanopsis eyrei)、柯(Lithocarpus glaber)、青榨槭(Acer davidii)、响叶杨(Populus adenopoda)、杉木(Cunninghamia lanceolata)、香港四照花(Dendrobenthamia hongkongensis)、化香树(Platycarya strobilacea)、日本杜英(Elaeocarpus japonicus),其他种较少。
水青冈群落灌木层共有77 种,占总物种数的74.76%。在样地内青冈、水青冈、亮毛杜鹃(Rhododendron microphyton)、硬壳柯(Lithocarpus hancei)为优势种,其次为野柿(Diospyros kakivar.silvestris)、细枝柃(Eurya loquaiana)、云贵鹅耳枥(Carpinuspubescens)、格药柃(Eurya muricata)、溪畔杜 鹃(Rhododendron rivulare) 、腺萼马银花(Rhododendron bachii) 、薄叶山矾(Symplocosanomala)、宜昌润楠(Machilus ichangensis)、多脉青冈,其他种较少。
水青冈群落草本层共有14 种,占总物种数的13.59%,以竹叶草(Oplismenus compositus)、沿阶草(Ophiopogon bodinieri) 占优势,绒叶斑叶兰(Goodyera velutina)、淡竹叶(Lophatherum gracile)、凤尾蕨(Pteris cretica)次之,建兰(Cymbidium ensifolium)、羊耳蒜(Liparisjaponica)等零星分布。
3.2 群落植物区系分析
根据吴征镒等[18]对我国种子植物科分布区类型的划分原则与方法,水青冈群落组成的47 科种子植物划分为10 个类型(表2)。科的区系类型以泛热带分布及其变型为主,共13 科,如樟科、山茶科(Theaceae)、山矾科(Symplocaceae)等,占27.65%;北温带分布科类型及其变型,如壳斗科、桦木科(Betulaceae)、金缕梅科(Hamamelidaceae)等以及世界分布,如蔷薇科、鼠李科(Rhamnaceae)、报春花科(Primulaceae)等次之,分别为11 科、9 科,分别占23.40%、19.15%。热带性分布型21 科,占44.68%;温带性分布型为17 科,占36.17%。说明本群落科的区系成分为热带性质占优势。

表2 雷公山水青冈群落物种组成Table 2 Species composition of Fagus longipetiolata community in Leigong Mountain
根据吴征镒等[19]对我国种子植物属分布类型划分,水青冈群落的种子植物属分布区类型有11 个,反映出水青冈群落地理成分复杂(表2)。以北温带分布及其变型,如杜鹃花属(Rhododendron)、桦木属(Betula)等为主,有16 属,占总属数的21.62%;泛热带分布及其变型,如紫金牛属(Ardisia)、山矾属(Symplocos)等和热带亚洲分布及其变型,如青冈属(Cyclobalanopsis)、润楠属(Machilus)、木荷属(Schima)等次之,各为11 属,分别占总属数的14.87%。热带性分布区类型34 属,占45.96%;温带性分布区类型35 属,占47.29%。说明本群落属的区系成分为温带性质占优势。

表3 雷公山水青冈群落植物科、属区系分析Table 3 Floristic analysis of Fagus longipetiolata community in Leigong Mountain
3.3 群落结构特征分析
3.3.1 优势种和重要值分析
从水青冈群落乔木层、灌木层和草本层物种的重要值可以看出(表4、表5、表6),乔木层水青冈(0.1702)重要值最大,为建群种;次之为青冈(0.0656)、云贵鹅耳枥(0.0534)、青榨槭(0.0485)、甜槠(0.0481)、西南木荷(0.0464),为优势种;此外重要值大于0.03 的还有木荷(0.0424)、柯(0.0372)、响叶杨(0.0372),为乔木层伴生种。灌木层重要值大于0.03 的有4 种,以青冈(0.1021)最大,水青冈(0.0562)、亮毛杜鹃(0.0461)、硬壳柯(0.0362)次之,为灌木层优势种;此外还有野柿(0.0288)、细枝柃(0.0283)、云贵鹅耳枥(0.0260)等。草本层重要值以竹叶草(0.2526)最大,沿阶草(0.1519)、绒叶斑叶兰(0.0965)、淡竹叶(0.0940)次之,为草本层优势种。

表4 雷公山水青冈群落乔木层重要值Table 4 Value of tree layer of Fagus longipetiolata community in Leigong Mountain
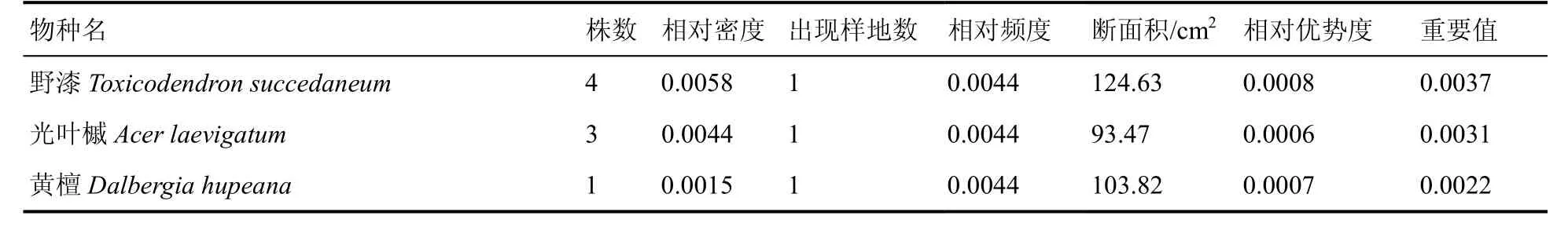
续表

表5 雷公山水青冈群落灌木层物种重要值Table 5 Important values of shrub layer species of Fagus longipetiolata community in Leigong Mountain

续表
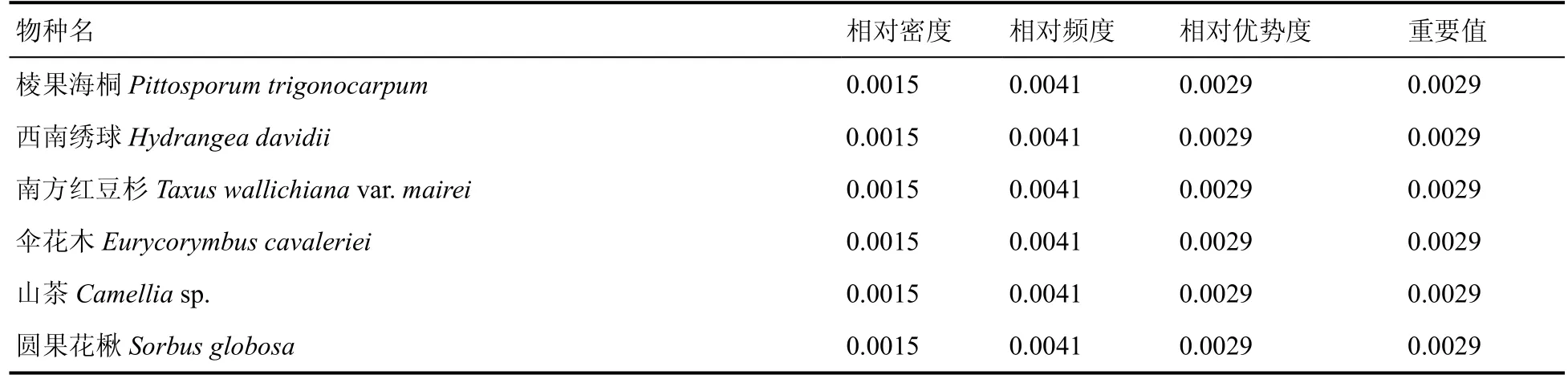
续表
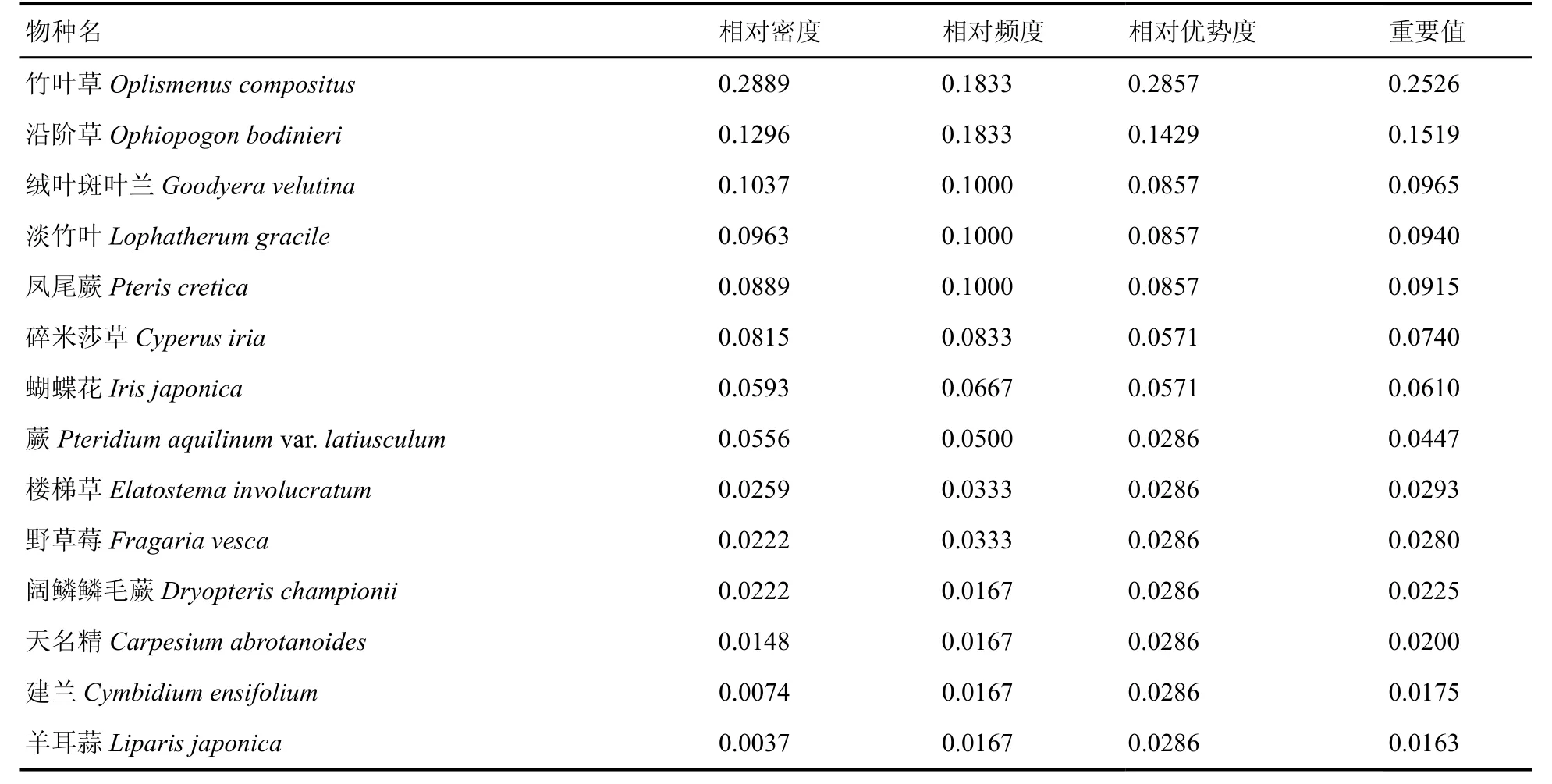
表6 雷公山水青冈群落草本层物种重要值Table 6 Important values of herbaceous layer species in the Fagus longipetiolata community in Leigong Mountain
3.3.2 群系优势种重要值
按照《中国植被》的命名原则,将雷公山水青冈群落分为7 个群系,各群系的优势种重要值如表7。水青冈是7 个群系中重要值处于第一位的物种,是主要建群种。在水青冈+云贵鹅耳枥群系中,水青冈重要值最大,为0.5321;其次是水青冈+木荷群系,为0.4637;水青冈+杉木群系最小,为0.2837。水青冈+青冈群系分布广,涉及1、3、9 号样地,是雷公山水青冈群落最常见的群系,其次水青冈+云贵鹅耳枥群系和水青冈+木荷群系,分别涉及4、8 号样地和5、7 号样地,也较常见。水青冈+杉木群系,涉及6 号样地,分布于山体中部,属偶见群系。

表7 雷公山水青冈群落各群系优势种重要值Table 7 Importance values of dominant species in each group of Fagus longipetiolata community in Leigong Mountain
3.3.3 群落立木结构与更新分析
种群的径级及高度级结构可反映不同年龄个体的水平分布及垂直分布,可作为群落动态及其发展趋势的指示,也在一定程度上反映种群与环境的相互关系,及其在群落中的作用和地位[15]。对群落重要值前3 的水青冈、青冈、云贵鹅耳枥进行径级、高度级分析(表8)。水青冈径级从Ⅰ级到Ⅴ级都有分布,Ⅰ级和Ⅴ级数量较多,分别为63 株和31 株,分别占总株数的47.73%和23.48%,呈双峰形分布;青冈Ⅰ级数量大,126 株,占总株数的72.00%,Ⅱ级和Ⅴ级数量少,分别为3 株和2 株。水青冈、青冈虽幼苗数量大,林内更新较好,但Ⅱ级数量少,说明种内竞争激烈,多数幼苗不能长成幼树。同时Ⅱ级数量少还是一个时期更新差的结果。在以前的调查中,水青冈、青冈均无幼苗。从表8 还可看出,水青冈Ⅳ级、Ⅴ级共有49 株,占总株数37.12%,大径级植株比重大。从调查来看,不同的群系,立木结构不相同,只有水青冈+青冈群系,水青冈、青冈从Ⅰ级到Ⅴ级有分布;其他群系多无Ⅰ级、Ⅱ级分布,存在年龄断层现象。云贵鹅耳枥有Ⅰ级和Ⅲ级、Ⅳ级,无Ⅱ级、Ⅴ级,存在年龄断层现象。

表8 雷公山水青冈群落径级、高度级结构表Table 8 Structure of diameter class and height class of Fagus longipetiolata community in Leigong Mountain
从高度级来看,水青冈、云贵鹅耳枥占据本群落的最高级Ⅶ级,是群落的主林层,青冈种群处于群落Ⅰ级至Ⅵ级,个体数量较多,是竞争的优势种。水青冈高度级Ⅵ级、Ⅶ级有21 株,占总株数15.91%,占3 种优势种Ⅵ级、Ⅶ级株数70.00%(表8)。
调查中发现,水青冈Ⅰ级、Ⅱ级个体均生长在林窗和稀疏林冠下,水青冈群落林窗的形成有利于水青冈更新,只有1、3、9、10 号样地有水青冈Ⅱ级个体,现有的Ⅰ级、Ⅱ级个体多是2008年雪凝灾害后更新起来的。不同的群系水青冈幼苗分布不同,水青冈+青冈群系幼苗多,3 个样地19 株,水青冈+云贵鹅耳枥群系幼苗2 个样地9株,水青冈+杉木群系幼苗1 个样地4 株,水青冈+甜槠群系幼苗1 个样地3 株,水青冈+木荷群系幼苗少,2 个样地只有1 株,反映出落叶阔叶林水青冈更新比针阔混交林的好,针阔混交林比常绿落叶阔叶混交林好。
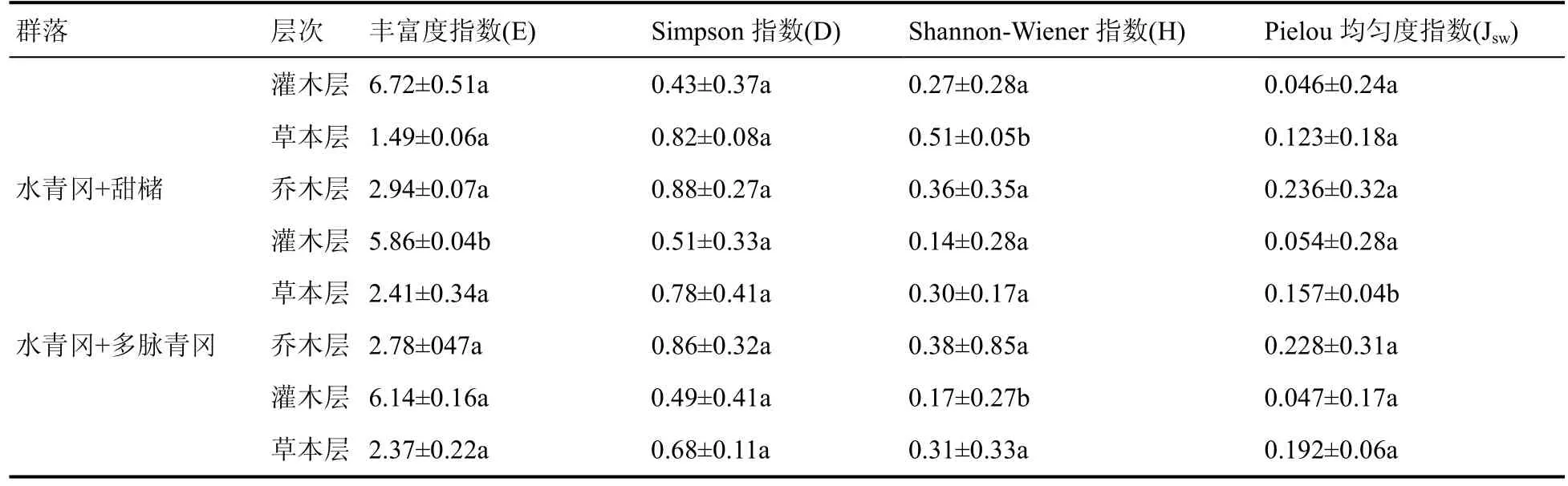
3.3.4 群落物种多样性
总的来看,水青冈群落各群系丰富度指数灌木层>乔木层>草本层;优势度指数乔木层>草本层>灌木层;H 多样性指数乔木层>草本层>灌木层;均匀度指数乔木层>草本层>灌木层(表9)。说明水青冈群落灌木层物种丰富度最明显;乔木层优势度最明显;乔木层H 多样性指数最高,草本层次之;乔木层均匀度指数最高,草本层次之。
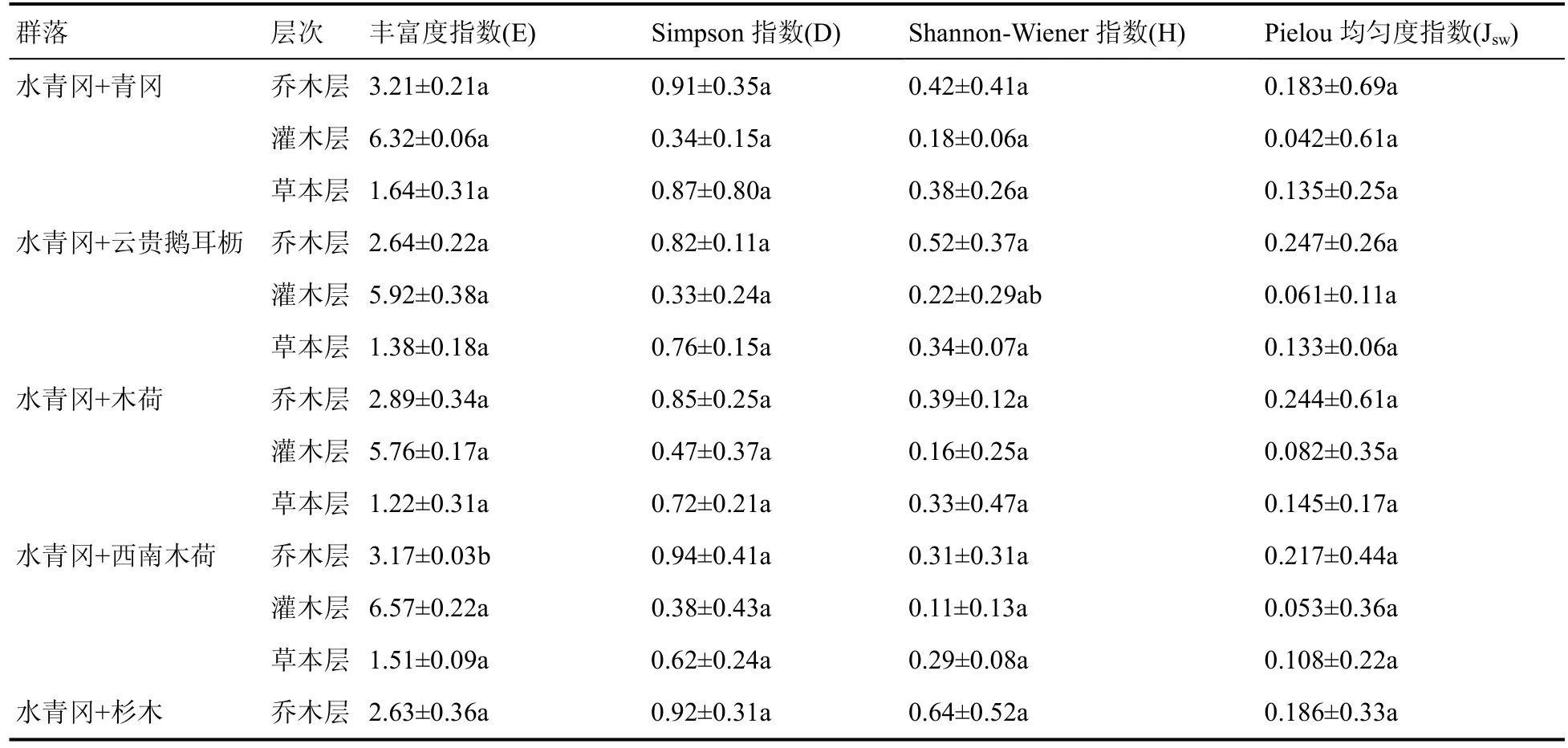
表9 雷公山水青冈群落物种多样性指数Table 9 Species diversity index of Fagus longipetiolata community in Leigong Mountain
续表
3.4 水青冈群落的分布格局
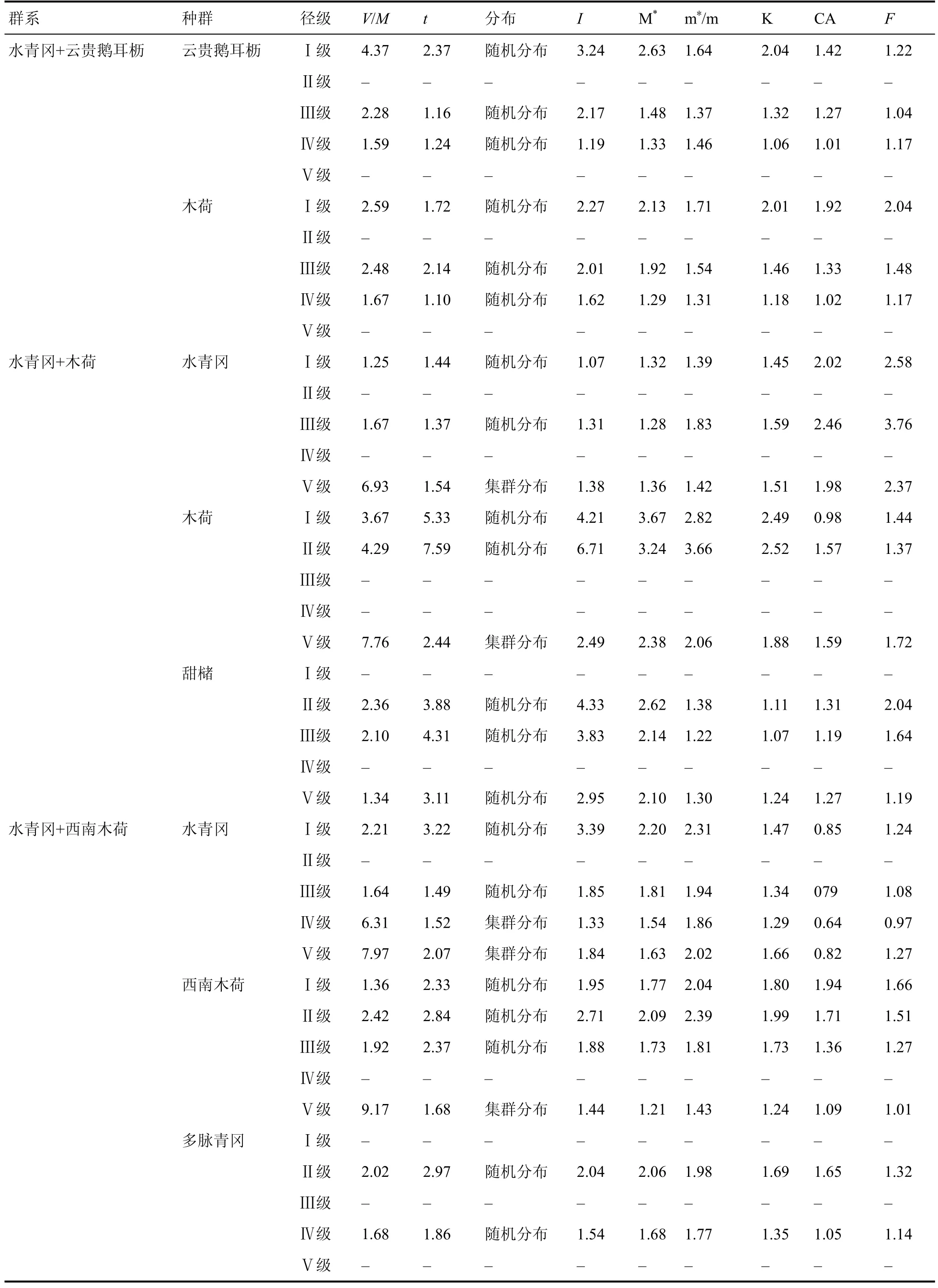
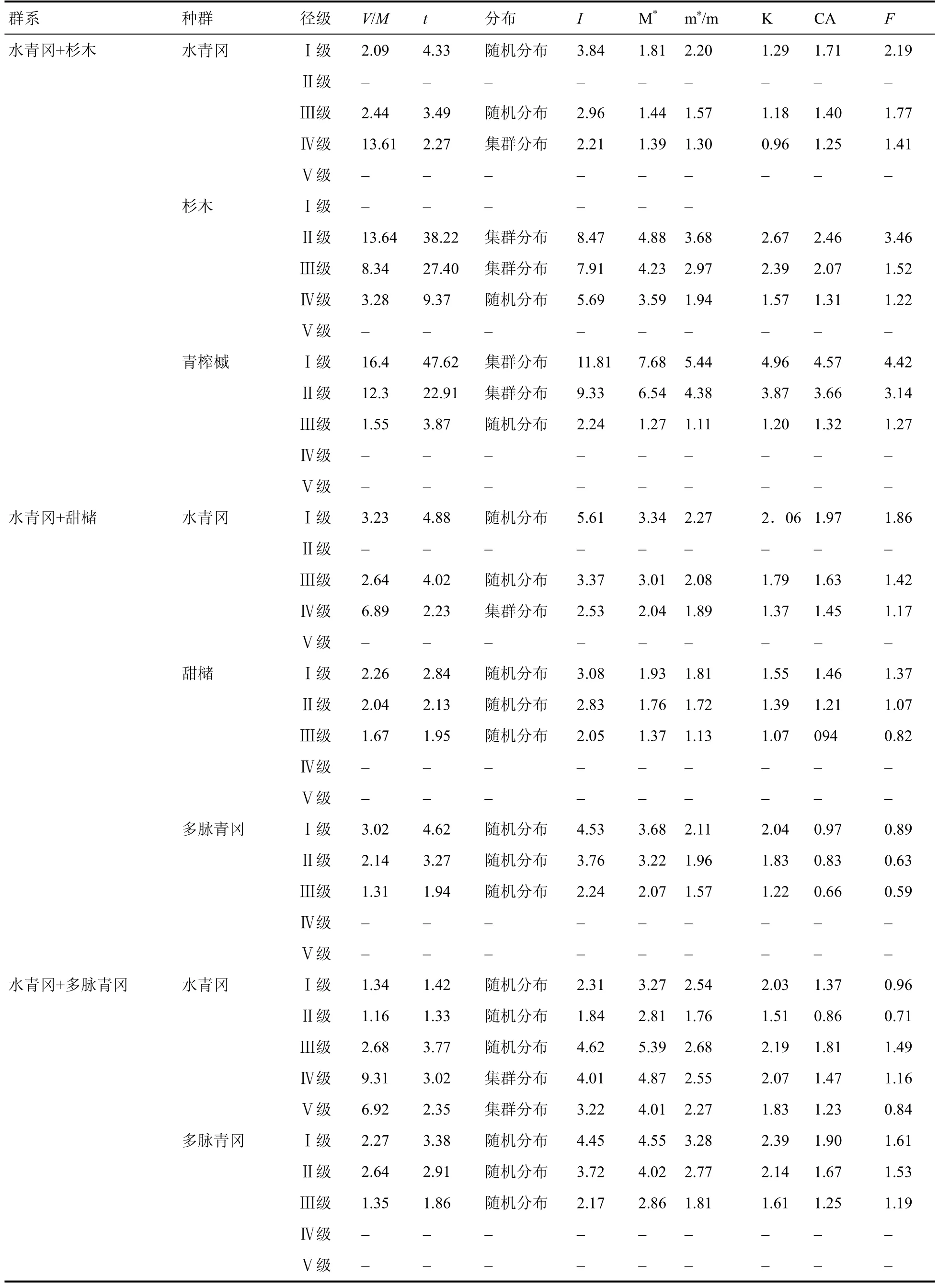
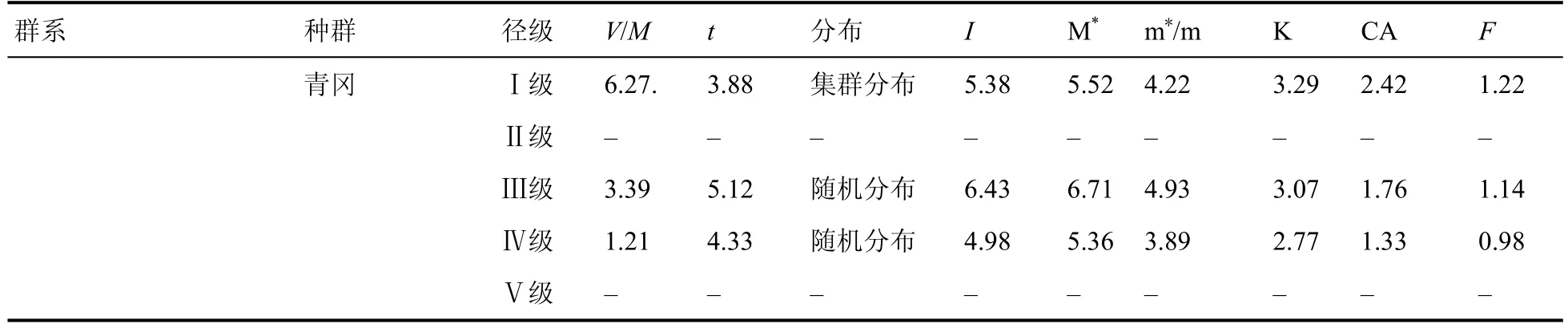
雷公山水青冈群落的7 个群系各优势种群的方差均值比(V/M)都明显大于 1 (表10),而且t检验值都为极显著差异。另外丛生指数 I 大于 0、平均拥挤指数值较高、负二项式指数 K 较小也验证了集群分布的结论。

表10 雷公山水青冈群落优势种群各立木级分布格局Table 10 Distribution pattern determination of dominant populations at each standing level in Fagus longipetiolata community in Leigong Mountain
续表
续表
续表
从表10 可以看出水青冈种群在Ⅰ、Ⅱ、Ⅲ级幼树呈随机分布,随着径级的增大水青冈种群由随机分布向集群分布过渡;优势种青冈则由集群分布向随机分布过渡;其他优势种多为随机分布。
4 结论与讨论
经调查,雷公山水青冈群落有维管植物50 科77 属103 种,物种组成较复杂、丰富。乔木层水青冈重要值最大(0.1702),是建群种,且各群系的水青冈重要值都较高,在群落中占优势地位;次之有青冈、云贵鹅耳枥、青榨槭、甜槠、西南木荷,为优势种,在群落中占有非常重要的地位。灌木层以青冈最大,水青冈、亮毛杜鹃次之,这三种是优势种。草本层重要值以竹叶草为优势种。与云南九龙山水青冈群落比较,雷公山水青冈群落物种更丰富,各样方水青冈重要值略高[9],两者在物种组成上存在较大差异;但比梵净山[7]水青冈群落物种数要少些,物种组成上也存在一定的差异。雷公山水青冈群落的物种组成具有自身的特点。
雷公山水青冈群落径级从Ⅰ级到Ⅴ级都有分布,Ⅰ级和Ⅴ级数量较多,呈双峰形分布,这说明该群落由衰退型向增长型转变。在调查中发现,水青冈Ⅰ级、Ⅱ级个体均生长在林窗和稀疏林冠下,水青冈群落林窗的形成有利于水青冈的更新,这作者早期调查结果基本相一致[11]。调查中,只有1、3、9、10 号样地有水青冈Ⅱ级个体,其余样地均无Ⅱ级个体,而作者早期调查无Ⅰ级个体[11],即现有的Ⅰ级、Ⅱ级个体多是2008 年雪凝灾害后更新的。调查时还发现当年的雪害使得多数水青冈大树断枝严重,形成林窗。外力(如雪害、风灾)使得水青冈群落形成林窗和稀疏林冠,是水青冈群落更新的驱动力。水青冈群落多个样地存在径级断层现象,可能是这一时期群落受外力影响较小,群落内郁闭度高,导致群落更新受阻。不同水青冈群系在更新上也存在差异,落叶阔叶林水青冈更新比针阔混交林的好,针阔混交林比常绿落叶阔叶混交林好,这是由于落叶阔叶林更易形成林窗,而常绿落叶阔叶混交林不易形成林窗,使得林下过荫所致,过荫的环境不利于水青冈更新[9—11]。
从高度等级分析来看,树高成层性明显,高度等级大致呈反“J”型分布,随着树高的增加,株数减少,常绿树木的比例也随之减少,这与梵净山水青冈属群落木本植物组成特征相似。
研究发现水青冈群落不同的群系生物多样性存在差异,这可能与群系的乔木层树种组成有较大的关系,乔木层阳性树种组成占比高的群系生物多样性相对较高,阴性树种组成占比高的群系生物多样性较低。这是因为阳性树种组成占比高的群系林内透光性稍好,林木对光照、养分等的竞争不太激烈,乔木层均匀指数稍低;而阴性树种组成占比高的群系,群系的乔木层均匀指数稍高。在森林群落,建群种乔木层均匀指数高导致林冠郁闭度变化,从而引起林下光照和湿度等条件的差异,直接制约灌、草层植物光合作用及有效养分的利用效率,从而制约群落总体多样性[20]。
种群空间分布格局的形成与种群自身的特性(种子扩散机制、植物繁殖特性、种内种间竞争等)、外部生境条件(地形、土壤、水分、光照等)有关,是不同尺度上各种生物因子和非生物因子长期综合作用的结果[21—22]。研究发现,水青冈种群在Ⅰ、Ⅱ、Ⅲ级幼树呈随机分布,随着径级的增大水青冈种群由随机分布向集群分布过渡,优势种群青冈则由集群分布向随机分布过渡,其他优势种多为随机分布。这是因为种间竞争激烈,特别是水青冈与青冈的竞争尤为激烈,随机分布有利于充分利用光照条件,减少种内竞争,维持更新能力。随着后期对环境的需求,种内竞争加剧,再加上种间竞争,个体死亡率升高,种群个体只有占据合理的空间才能降低急剧竞争而生存下来,于是种群分布格局逐渐集群分布。不同群系各优势种群的空间分布格局有所不同,水青冈+青冈群系在幼苗阶段是集群分布,然后向随机分布再向集群分布转变,这可能是水青冈群落的林窗多集中在水青冈大树下的缘故。
总的来看,雷公山水青冈群落其物种组成既不同于云南等地,也不同于贵州梵净山的水青冈群落,其径级结构呈双峰形分布,在自然状态下群落能由衰退型向增长型转变,具有自我循环和修复的能力,是水青冈群落作为雷公山的一种地带性群落得以长期保存缘故。
致 谢:外业调查中台江县南宫镇生态护林员给予大力支持,谨此致谢。
