牙龈卟啉单胞菌来源的脂多糖通过X盒结合蛋白1调控脂肪细胞胰岛素信号通路的机制研究
2022-03-31陆佳艺伍倩琪陈伊燕叶蕾蕾苏媛
陆佳艺伍倩琪陈伊燕叶蕾蕾苏媛,
1.南方医科大学顺德医院(佛山市顺德区第一人民医院)口腔中心,顺德528300;
2.南方医科大学口腔医院牙周科,广州510280
牙周炎是口腔中常见的慢性炎症性疾病,主要表现为牙齿周围软硬组织的炎性破坏[1]。研究发现,牙周炎对2型糖尿病(type 2 diabetes mellitus,T2DM)的发生发展具有促进作用[2-3],但具体机制尚不清楚。牙龈卟啉单胞菌(Porphyromonas gingivalis,P.gingivalis)是牙周炎的主要致病菌,其外膜蛋白脂多糖(lipopolysaccharid,LPS)是主要毒力因子。牙龈卟啉单胞菌脂多糖(Porphyromonas gingivalis-lipopolysaccharid,P.gingivalis-LPS)通过破溃的牙周组织进入血循环到达远隔器官,影响机体的糖代谢水平[4-5]。而作为糖代谢的重要靶组织脂肪组织,是否受牙周炎的影响目前尚未证实。
近年来,内质网应激(endoplasmic reticulum stress,ERS)的关键信号分子X盒结合蛋白1(Xbox binding protein 1,XBP1)被证实可受P.gingivalis-LPS的调控[6],而XBP1同时又参与胰岛素信号通路的传导[7-8]。因此,XBP1有望成为P.gingivalis-LPS调控大鼠脂肪细胞胰岛素信号的关键靶点。本实验通过体外构建pLVX-XBP1载体、p-LVX-XBP1-RNAi载体分别转染到大鼠原代脂肪细胞中,采用P.gingivalis-LPS刺激转染后的细胞,验证XBP1能否调控大鼠脂肪细胞胰岛素信号通路关键蛋白胰岛素受体底物(insulin receptor substrate,IRS)-1、下游磷脂酰肌醇依赖性激酶(phosphoinositide dependent protein kinase,PDK)-1和蛋白激酶B(protein kinase B,PKB),又称AKT的表达。
1 材料和方法
1.1 材料
4~5周雄性SD大鼠(南方医科大学动物实验中心),杜氏培养液(Dulbecco’s Modified Eagle Medium/F12,DMEM/F12)、胎牛血清(Gibco公司,美国),油红O试剂盒(南京建成生物工程研究所),多聚赖氨酸(Sigma公司,美国),P.gingivalis-LPS(InvivoGen公司,美国),pLVX-XBP1慢病毒、pLVX-XBP1-RNAi慢病毒、逆转录试剂盒、实时定量聚合酶链反应(real-time quantitative polymerase chain reaction,RT-qPCR)试剂盒(重庆威斯腾生物医药科技有限责任公司),ab245314型IRS-1抗体(Abcam公司,美国),3848T型p-PDK-1抗体、4060T型p-AKT抗体(CST公司,美国)。
1.2 实验方法
1.2.1 分离培养原代大鼠脂肪细胞 断颈处死SD大鼠后,在无菌条件下剪开大鼠腹部皮毛,取附睾脂肪组织,采用眼科剪将脂肪组织剪成0.5~1.0 mm大小的均匀颗粒,采用0.1%Ⅰ型胶原酶消化液在37℃条件下消化脂肪组织,每隔5 min手动振荡30 s以促进消化,消化时间为60 min,直到脂肪组织被消化成乳糜液样液体。将消化后的乳糜样液体加入4 mL PBS,轻轻吹打混匀,通过200目孔径的不锈钢细胞筛过滤,滤液转移入离心管中,向离心管中加入10 mL PBS洗细胞3次。以250g·min-1的转速离心5 min,用吸管吸出上清液,加入另一试管中,加入PBS 10 mL洗细胞,再次离心、去除下清液,重复此步骤2次。再用10 mL含20%胎牛血清的DMEM/F12细胞培养液洗涤并重悬细胞,调整密度为2×106个·mL-1,取50μL加入每孔2 mL的培养液的6孔板中,盖以20 mm×20 mm多聚赖氨酸处理过的无菌盖玻片,放37℃、5%CO2饱和湿度培养箱中培养。
1.2.2 油红O鉴定原代大鼠脂肪细胞 用油红O固定液固定原代大鼠脂肪细胞10~15 min,吸出固定液后,放于流通的空气中10~15 min,加入新配制好的油红O染色剂浸染15 min;吸出染色液,加入60%异丙醇漂洗20~30 s,流水冲洗,蒸馏水稍微清洗;吸出蒸馏水,加入苏木素染色液,复染核2 min;去除苏木素染色液,流水冲洗,晾干;200倍倒置光学显微镜观察拍照。
1.2.3 蛋白质印迹(Western Bolt)检测P.gingivalis-LPS刺激脂肪细胞后IRS-1、磷酸化PDK-1(p-PDK-1)、磷酸化AKT(p-AKT)的表达 将培养48 h的脂肪细胞接种于6 cm培养皿24 h后,分为空白对照组和刺激组,空白对照组为100 ng·mL-1P.gingivalis-LPS刺激的大鼠脂肪细胞0 h;刺激组为100 ng·mL-1P.gingivalis-LPS分别刺激大鼠脂肪细胞4、8、12、24 h。刺激后提取细胞总蛋白,用BCA蛋白质定量法检测总蛋白浓度,每个上样孔等量蛋白(20μg)通过聚丙烯酰胺凝胶电泳分离蛋白后,10%脱脂牛奶摇床封闭1 h;将含有目的蛋白IRS-1、p-PDK-1、p-AKT和内参蛋白β肌动蛋白(β-actin)的聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF)放于相应的抗体中,4℃孵育过夜;用含Tween 20的Tris缓冲盐水(Tris buffered saline with Tween 20,TBST)漂洗后,加入辣根过氧化物酶标记的免疫球蛋白(immunoglobulin,Ig)G二抗室温孵育1 h。TBST漂洗,化学发光显影并拍照。用Image J图像分析软件对各组条带行灰度值分析,以β-actin作为内参,目的蛋白与内参蛋白的灰度比值为IRS-1、p-PDK-1、p-AKT蛋白表达的相对表达水平。
1.2.4 RT-qPCR检测过表达、干扰XBP1慢病毒载体转染的脂肪细胞 过表达XBP1慢病毒载体pLVX-XBP1、干扰XBP1慢病毒载体pLVX-XBP1-RNAi由重庆威斯腾生物医药科技有限责任公司构建。实验分为空白对照组、空载过表达慢病毒(pLVX-NC1)、过表达XBP1慢病毒(pLVX-XBP1)、空载干扰慢病毒(pLVX-NC2)、干扰XBP1慢病毒(pLVX-XBP1-RNAi)。
将pLVX-NC1、p-LVX-XBP1、pLVX-NC2、pLVX-XBP1-RNAi分别转染到大鼠原代脂肪细胞后,分别接种于6 cm培养皿24 h后,用TRIzol法分别提取5个实验组细胞的总RNA,并测定RNA浓度和纯度。用逆转录试剂盒将RNA逆转录成互补DNA,RT-qPCR检测转染后的脂肪细胞内XBP1的mRNA表达水平。
XBP1引物序列:上游AAAGAAAGCCCGGATGAGC,下游ATTCATCCCCAAGCGTGTCC。甘油醛-3-磷酸脱氢酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)引物序列:上游AGTGCCAGCCTCGTCTCATA,下游GAGAAGGCAGCCCTGGTAAC。扩增条件均为95℃30 s;然后反复95℃5 s,60℃30 s循环40次。以2-ΔΔCT公式法计算目的基因XBP1的mRNA相对表达水平。
1.2.5 Western Bolt检测在过表达XBP1、干扰XBP1慢病毒转染脂肪细胞后P.gingivalis-LPS刺激下IRS-1、p-PDK-1、p-AKT的表达 本实验采用以下4组:pLVX-NC1、pLVX-XBP1、pLVX-NC2、pLVX-XBP1-RNAi分别转染大鼠原代脂肪细胞,分别接种于6 cm培养皿24 h,通过100 ng·mL-1P.gingivalis-LPS刺激转染后的大鼠脂肪细胞4、8、12、24 h,用BCA法检测蛋白浓度。
每个上样孔皆取等量蛋白(20μg)通过聚丙烯酰胺凝胶电泳分离蛋白后,10%脱脂牛奶摇床封闭1 h。将含有目的蛋白IRS-1、p-PDK-1、p-AKT和内参蛋白β-actin的PVDF膜放于相应的抗体中,4℃孵育过夜。TBST漂洗后,加入辣根过氧化物酶标记的IgG二抗室温孵育1 h。TBST漂洗,化学发光显影并拍照。用Image J图像分析软件对各组条带行灰度值分析,以β-actin作为内参,目的蛋白与内参蛋白的灰度比值为IRS-1、p-PDK-1、p-AKT蛋白表达相对表达水平。
1.3 统计学分析
实验数据用GraphPad Prism 7软件进行分析,实验数据以均数±标准差表示,两组间单因素比较采用t检验,组间双因素比较采用双因素方差分析,P<0.05为差异有统计学意义。研究数据均来自3次或3次以上独立实验。
2 结果
2.1 大鼠脂肪细胞原代分离培养以及油红O鉴定脂肪细胞
原代细胞培养的形态观察显示,细胞贴壁后为类圆形如图1所示。培养2周后,胞内出现脂肪颗粒积聚,经过油红O染色,确定细胞分化成为脂肪细胞如图2所示。
图1 原代培养的大鼠脂肪细胞 倒置光学显微镜 ×200Fig 1 Primary cultured rat adipocytes inverted optical microscope ×200

图2 油红O染色鉴定原代大鼠脂肪细胞 ×200Fig 2 Primary rat adipocytes were identified by red staining O×200
2.2 Western Bolt检测P.gingivalis-LPS刺激脂肪细胞后胰岛素信号通路蛋白表达结果
Western Bolt检测P.gingivalis-LPS刺激脂肪细胞0、4、8、12、24 h后,胰岛素信号通路IRS-1、p-PDK-1、p-AKT的蛋白表达如图3A所示,图3B~D所示刺激4、8、12、24 h后,IRS-1、p-AKT、p-PDK-1蛋白表达与空白对照组0 h相比呈明显下降趋势,差异有统计学意义(P<0.05)。统计结果表明,脂肪细胞在P.gingivalis-LPS刺激下,胰岛素信号通路蛋白IRS-1、PDK-1、AKT受抑制,随着刺激时间的推移,IRS-1、p-PDK-1、p-AKT蛋白的表达呈下降趋势。
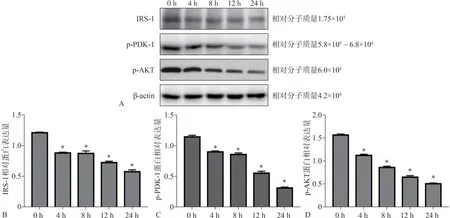
图3 Western Bolt检测P.gingivalis-LPS刺激脂肪细胞后IRS-1、p-PDK-1、p-AKT的蛋白表达结果Fig 3 Western Bolt detected theprotein expression of adipocytes IRS-1,p-PDK-1 and p-AKT after stimulated by P.gingivalis-LPS
2.3 RT-qPCR检测过表达、干扰XBP1慢病毒载体转染脂肪细胞结果
结果如图4所示,pLVX-XBP1组XBP1 mRNA相对表达量为2.06±0.01,与pLVX-NC1组(1.03±0.01)相比明显升高,差异有统计学意义(P<0.05);pLVX-XBP1-RNAi组XBP1 mRNA相对表达量为0.53±0.03,与pLVX-NC2组(1.12±0.1)相比明显降低,差异有统计学意义(P<0.05)。综上,p-LVX-XBP1、pLVX-XBP1-RNAi成功构建。
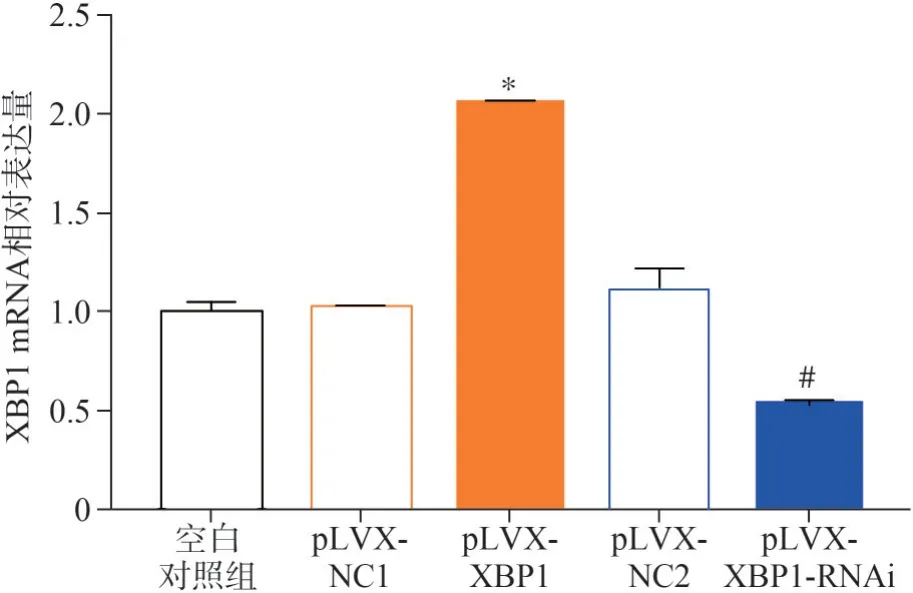
图4 qRT-PCR检测慢病毒转染脂肪细胞结果Fig 4 qRT-PCR detected the transfection results of lentivirus into adipocytes
2.4 Western Bolt检测在pLVX-XBP1、pLVX-XBP-1-RNAi转染脂肪细胞后P.gingivalis-LPS刺激下胰岛素信号通路的表达
pLVX-XBP1转染脂肪细胞后,Western Bolt检测胰岛素信号通路IRS-1、p-PDK-1、p-AKT的蛋白表达如图5A所示,图5B~E所示,P.gingivalis-LPS刺激4 h后,pLVX-XBP1组IRS-1的蛋白表达明显高于pLVX-NC1组,差异有统计学意义(P<0.05);pLVX-XBP1组p-PDK-1的蛋白表达略低于pLVX-NC1组,差异无统计学意义;pLVX-XBP1组p-AKT的蛋白表达明显低于pLVX-NC1组,差异有统计学意义(P<0.05)。刺激8、12 h后,pLVX-XBP1组IRS-1、p-PDK-1、p-AKT的蛋白表达明显高于pLVX-NC1组,差异有统计学意义(P<0.05)。刺激24 h后,pLVX-XBP1组IRS-1的蛋白表达明显高于pLVX-NC1组,差异有统计学意义(P<0.05);pLVX-XBP1组p-PDK-1、p-AKT的蛋白表达明显低于pLVX-NC1组,差异有统计学意义(P<0.05)。

图5 Western Bolt检测pLVX-XBP1转染脂肪细胞后P.gingivalis-LPS刺激下IRS-1、p-PDK-1、p-AKT的蛋白表达结果Fig 5 Western Bolt detected theexpression of protein IRS-1,p-PDK-1 and p-AKT,stimulated by P.gingivalis-LPSafter pLVX-XBP1 transfecting adipocytes
pLVX-XBP1-RNAi转染脂肪细胞后,Western Bolt法检测胰岛素信号通路IRS-1、p-PDK-1、p-AKT的蛋白表达如图6A所示,如图6B~E所示,P.gingivalis-LPS刺激4、8、12、24 h后,pLVXXBP1-RNAi组IRS-1、p-PDK-1、p-AKT的蛋白表达皆明显低于pLVX-NC2组,差异有统计学意义(P<0.05)。
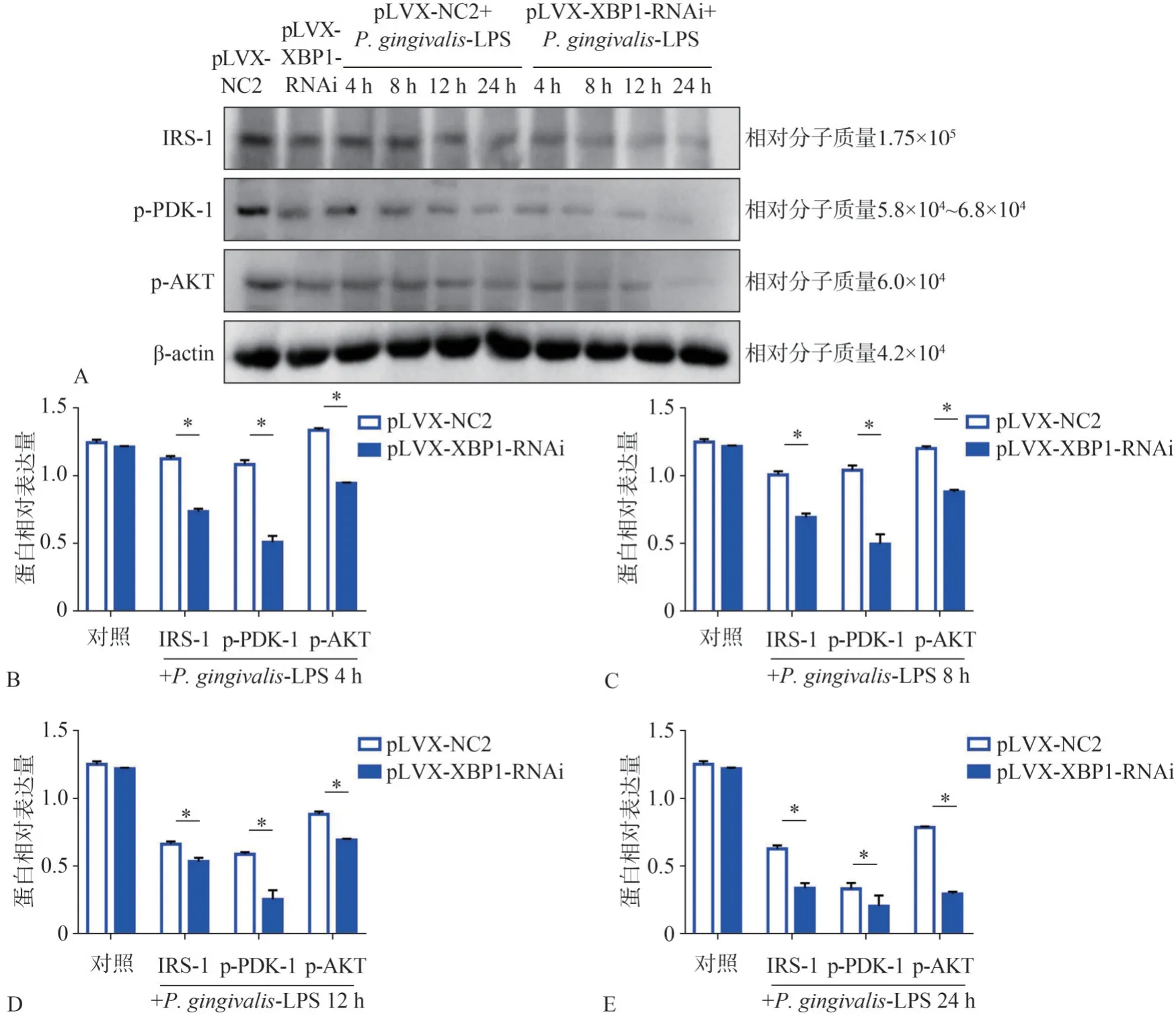
图6 Western Bolt检测pLVX-XBP1-RNAi转染脂肪细胞后P.gingivalis-LPS刺激下IRS-1、p-PDK-1、p-AKT的蛋白表达结果Fig 6 Western Bolt detected the expression of protein IRS-1,p-PDK-1 and p-AKT,stimulated by P.gingivalis-LPSafter pLVX-XBP1-RNAi transfecting adipocytes
3 讨论
流行病学调查[9]显示,牙周炎的严重程度与T2DM的糖代谢水平相关。在临床上,牙周基础治疗能降低糖化血红蛋白和空腹血糖,改善患者的糖代谢水平[10]。因此,牙周炎对T2DM的影响意义重大,是目前交叉学科领域的研究热点之一。
T2DM的重要病理机制是胰岛素抵抗(insulin resistance,IR),它是指肝脏、脂肪和骨骼肌等胰岛素靶组织对胰岛素敏感性下降,胰岛素信号传导受阻,造成糖代谢异常[11]。其中,脂肪组织胰岛素信号受阻对全身糖代谢水平有重要影响[12],是诱导机体发生IR的起始[13]。研究[14]发现,牙周炎可引起大鼠内脏脂肪组织发生炎症性改变,并且这种改变与机体的IR高度相关。但牙周炎能否造成脂肪组织胰岛素信号通路的改变,目前尚未见报道。本研究发现,牙周炎的主要毒力因子P.gingivalis-LPS可造成脂肪细胞IRS-1蛋白表达降低,初步揭示了牙周炎可调控脂肪细胞的胰岛素信号。胰岛素通过与靶细胞表面的胰岛素受体相结合激活IRS,调控下游信号转导。IRS-1是IRS家族中的重要成员,对胰岛素信号起关键调节作用。IRS-1的激活进一步促进下游PDK-1和AKT的磷酸化,从而促进葡萄糖转运,糖原合成等多种生物学效应。因此,IRS-1、PDK-1、AKT通路作为一条经典的糖代谢通路被广泛关注。本研究中,P.gingivalis-LPS刺激大鼠脂肪细胞后,IRS-1的蛋白表达减少,其下游的PDK-1和AKT磷酸化水平降低,说明P.gingivalis-LPS可抑制大鼠脂肪细胞中的IRS-1、PDK-1、AKT信号通路。
本研究旨在揭示P.gingivalis-LPS通过何种机制调控脂肪细胞IRS-1信号通路,结果显示,ERS相关信号分子XBP1是P.gingivalis-LPS调控IRS-1信号通路的关键靶点。ERS是真核细胞的一种保护性应激反应,细胞受外界刺激发生ERS时,位于内质网腔内的分子伴侣蛋白-结合免疫球蛋白(binding immunoglobulin protein,BiP)与内质网的3种跨膜蛋白解离并主动结合到未折叠或错误折叠蛋白,激活内质网跨膜蛋白并启动3条信号通路:肌醇需要酶1α(inositol-requiring enzyme 1α,IRE1α)、双链RNA依赖的蛋白激酶R样内质网激酶(PKR like-ER-resident kinase,PERK)和活化转录因子6(activating transcriptionfactor 6,ATF-6)。近年来研究表明,ERS在IR的发生发展中扮演重要作用,而XBP1是其中的关键信号分子。XBP1的活化可促进脂联素的多聚化,改善肥胖小鼠的葡萄糖耐量和胰岛素敏感性。这与本实验结果相一致,抑制XBP1的脂肪细胞在P.gingivalis-LPS刺激后,IRS-1、PDK-1和AKT表达明显下降,胰岛素信号通路表达受阻,胰岛素敏感性降低;过表达XBP1的脂肪细胞在P.gingivalis-LPS刺激后,IRS-1的表达升高,说明上调XBP1能改善IRS-1的蛋白表达;P.gingivalis-LPS刺激8、12 h后,PDK-1和AKT表达升高,但24 h后明显下降,初步分析调控PDK-1和AKT信号通路的上游较为复杂,可能受XBP1以外的其他信号通路的影响,具体机制还有待进一步探索。因此,本研究证实XBP1是介导牙周炎调控脂肪组织IRS-1信号通路的关键靶点。
综上,P.gingivalis-LPS刺激大鼠脂肪细胞后,胰岛素信号通路IRS-1/PDK-1/AKT的表达随时间推移呈下降趋势。过表达XBP1的脂肪细胞,IRS-1、PDK-1、AKT的蛋白表达水平得到改善;而抑制XBP1的脂肪细胞,IRS-1、PDK-1、AKT的蛋白表达受阻。本研究通过体外模型证实,牙周毒力因子P.gingivalis-LPS可通过XBP1调控脂肪细胞IRS-1、PDK-1、AKT信号通路。
利益冲突声明:作者声明本文无利益冲突。
