高寒草甸根系分布特征及与土壤温湿度关系的研究
2022-03-28房昊天翁白莎常文娟杨裕恒宫晓艳
房昊天, 翁白莎, 常文娟, 杨裕恒, 宫晓艳
(1. 三峡大学水利与环境学院, 湖北 宜昌 443002; 2. 流域水循环模拟与调控国家重点实验室中国水利水电科学研究院,北京 100038)
根系作为植物生长和发育的核心器官,是陆地生态系统中地上与地下沟通的桥梁,起到物质及信息交换的作用。土壤中的水分在渗透压的作用下通过根系吸水进入植株[1],会对其根系的空间分布状况产生直接影响[2],随着土壤含水量的增大,根系分布的分形维数有增大的趋势[3],当土壤含水量高于一定阈值时,植物根系呼吸作用会受到抑制,导致烂根、地下生物量减少[4]。土壤水与根系作用是相互的,根系的分布格局和存量变化也是影响根区土壤水分参数的基本要素[5]。另外,温度是促进根系分布格局和存量变化的关键因子[6]。由于根系对周围环境变化敏感,适当的增温会促进草本植物地下生物量的增加[7],而温度过高或过低都会抑制根系的发育[8-9],导致根系短而弱,在土壤中的垂直分布较浅,无法形成壮棵[9]。由于青藏高原冻土区土壤水热动态变化过程与气候变化联系紧密[10],植物地下部分的响应是地区生态系统气候变暖评估中必不可少的组成部分[11]。因此,研究草地根系分布特征及与土壤温湿度的关系十分重要。
高寒草甸是青藏高原主要的植被类型之一,其地下生物量占总生物量的80%以上[12]。目前对高寒草甸根系与环境因子的研究主要集中在以下三个方面:一、根系生物量等特征参数的地下分配格局,如呈“T”形态锯齿状分布[13],部分研究发现季节性冻融会导致高寒草甸根系分布浅层化[14];二、增温、积雪等气候因素变化对根系生物量的影响,如马丽等[15]发现梯度增温对地下生物量的影响不显著;三、放牧、草地恢复措施、氮和磷添加[16]等人为干扰措施对根系特性的影响,如陈冬明等[17]研究表明,轻度放牧和适当短期休牧能够增加根系分泌速率,并可能通过正反馈作用刺激植物地上补偿生长。但这些研究多是集中在地下生物量方面,而对根系形态特征如根系密度、根系深度等的研究还较少。在青藏高原气候变暖的背景下,冻土融化,高寒草甸的根系深度是否会增加?土壤温度升高,残根腐烂速率增加,根系密度又是否会减少?这些还有待于深入探索。
本研究以怒江源区那曲流域的高寒草甸为研究对象,以高寒草甸的根系形态特征参数作为切入点,利用微根管法[18]对高寒草甸根系进行生态调查监测,探讨植被根系的分布特征以及环境因子对其造成的影响,旨在揭示怒江源区高寒草甸根系分布状况,对改善该地区的环境和维持生态的可持续发展有着重要的意义。
1 材料与方法
1.1 研究区概况
那曲流域位于西藏北部那曲地区,是怒江的源头区,地理坐标为91°08′~92°54′ E,30°53′~32°44′ N,流域控制面积16 967 km2,平均海拔4 500 m。流域大部分区域属高原亚寒带季风半湿润气候区,其气候特征是年均气温低,多年平均气温—0.6℃。该地年降水量在317.4 mm~610.5 mm之间,降水集中于5—9月,占全年降水总量的80%,年际降水分布不均;流域主要冻土类型为多年冻土和季节冻土,分布广泛;主要植被类型为高寒草甸,以嵩草属(Kobresia)植物为建群种,优势种有小嵩草(Kobresiapygmaea)、矮嵩草(Kobresiahumilis)等,生长季为5—9月,其中返青期主要集中在5—6月。
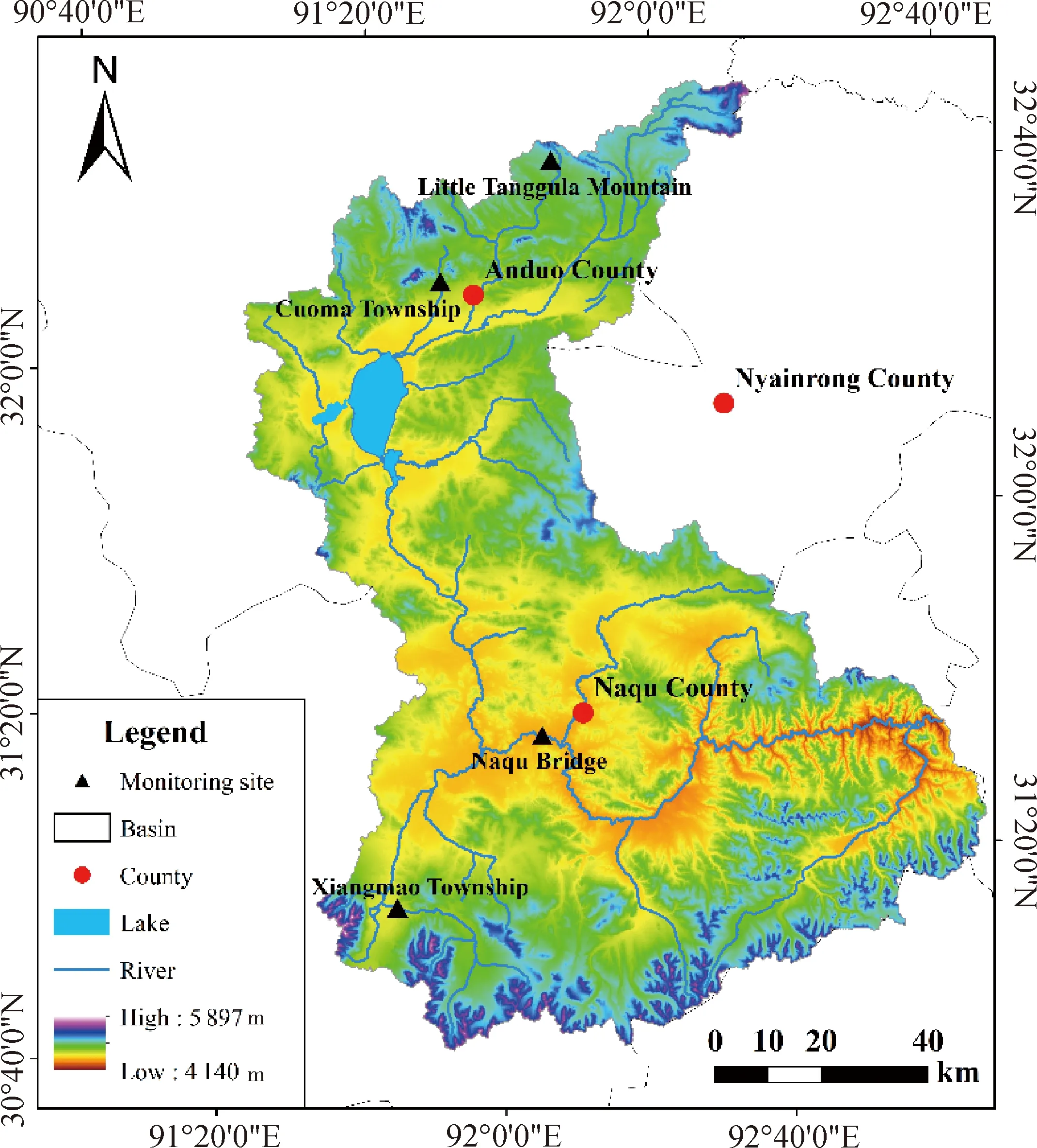
图1 那曲流域地理位置及监测点Fig.1 The geographical location and monitoring sites of Naqu watershed
1.2 试验设计
经过实地考察与勘测,按海拔梯度在流域内选择小唐古拉山、措玛乡、香茂乡、那曲大桥4个监测点(表1),在每个监测点分别选择微生境和植被覆盖类型相似的区域埋设硬质聚氯乙烯透明微根管(长100 cm,外径7 cm,内径6.4 cm)。提前一年埋设,使根系在微根管周围的空间定植和营养水平尽可能恢复到扰动前的水平,且每个试验场地埋设5根微根管以保证覆盖较全的根系信息。微根管与地面呈30°夹角,垂直深度约为40 cm,地面部分用橡胶盖密封防止进水。

表1 样地调查特征Table 1 Characteristics of the sampling plots
分别在四处布设的试验场地埋设土壤温湿度传感器(仪器型号5TM),自动布控土壤温度和土壤含水量(所测为体积含水率),测量精度分别为±1℃,±2%,按5 cm,10 cm,20 cm,35 cm的土壤深度每小时自动记录一次,时间跨度为1年。
1.3 根系特征参数的获取与计算
分别于2020年5月和2020年11月采集根系图像,使用AZR-100仪器镜头伸入微根管从地表往下拍摄,镜头拍摄范围为1.35 cm×1.35 cm,为保证拍摄根系的完整性,同一层每拍完一张照片后顺时针旋转45°,即拍摄8张照片后下移一层,直到镜头内无法找到根系为止,为了更好地与土壤温湿传感器埋设深度对应,仪器镜头每层下移距离约为5 cm。
采用WinRHIZO Tron MF根系分析软件对采集的图片进行处理,测定每张图片中根系的长度、表面积、投影面积、体积、根系直径等数据。考虑到有的微根管会出现损坏、浸水等情况,拍摄的图片并不清晰,因此将各微根管相同土层的根系数据取平均值处理。
单位体积根长密度(Root length density per unit volume,RLDV,m·m-3)[18]按公式(1)计算:
RLDV=RL/(A×DOF)
(1)
式中,RL(m)为微根窗中观察到的根系长度,A(m2)为采集图像面积,DOF(m)是微根窗管到周围土壤的距离。其中DOF在本次试验取固定的0.003 m。
1.4 数据统计与分析
使用SPSS 24.0对高寒草甸根系表面积、体积、根长密度等参数进行单因素方差分析,最小显著极差法(LSD)进行多重比较,不同季节间的差异采用配对样本T检验。并用Canoco 4.5采取冗余分析法(RDA)对土壤环境因子(温度、湿度)与根系特征参数(根系长度、表面积、体积、不同根径所占百分比)进行相关性分析,其中环境向量与根系参数向量夹角的余弦值代表之间的相关系数,若夹角小于90°,表示之间正相关;大于90°,表示之间负相关;此外,从样方点到各因子的箭头投影点,可以表示该因子的值在样方内的情况。采用Microsoft Office制作图表。
2 结果与分析
2.1 高寒草甸根系分布特征
2.1.1返青期(5月)和枯黄期(11月)根系分布特征 根据根系图像质量(无雨污覆盖在微根管表面,根系完整可见)选择拍摄效果最优的香茂乡作为研究对象,对比返青期和枯黄期不同土壤深度下的根系特征参数。
由图2可知,返青期5~35 cm深度草甸根系整体长度、表面积、体积随着土壤深度的增加均呈先下降后上升再下降趋势,其中35 cm土层根系参数最小,与其它各土层差异显著(P<0.05)。枯黄期草甸根系整体根长随着土壤深度的增加呈先上升后下降趋势,表面积、体积则是呈先下降后上升再下降趋势,同样在35 cm土层处达到最小值。

图2 不同时间下根系特征参数随土壤深度的变化Fig.2 Variation of root characteristic parameters with soil depth at different time注:不同大写字母表示同一生长季不同土壤深度间差异显著(P<0.05),不同小写字母表示同一土壤深度不同生长季间差异显著(P<0.05)Note:Different capital letters indicate significant differences between soil depths at the 0.05 level for the same growing season,different lowercase letters indicate significant differences between growing seasons at the 0.05 level for the same soil depth
就生长季节而言,返青期草甸的根长在土层5 cm和25 cm的长度均要大于枯黄期,季节间差异显著(P<0.05);在其余土层返青期与枯黄期根长差异不显著。在表层土壤至土深20 cm处,根系的表面积、投影面积和体积枯黄期要大于返青期,在25 cm处返青期根系的表面积、投影面积和体积则大于枯黄期,随后面积和体积均下降,在土深30 cm与35 cm处与枯黄期无显著性差异。
2.1.2根系特征参数垂直分布特征 由图3(a)可知,高寒草甸根系主要分布在0~25 cm土层中,约占地下根系总量的80%。其中土深25 cm处根长密度最大,为7.41 m·m-3,占总根长密度的18.8%;其次为土深5 cm处,根长密度为6.87 m·m-3,占总根长密度的17.5%;35 cm土层处根长密度最小,为2.27 m·m-3,仅占总根长密度的5.8%,与25 cm土层处相差13%。
图3(b)则是不同土壤深度根系径级分布状况。可以看出,直径大于1 mm的根基本分布在0~15 cm的土壤表层,且随着土壤深度的增加而减少;0.5~1 mm根的比例则随土壤深度的增加先升高后降低;而0~0.5 mm细根的占比随土壤深度的增加呈先上升后下降再上升趋势,至30~35 cm土层达到最大值,约占这一层根系的91%。
具有良好的抗压性能,这是保障路面可以承受重物反复碾压的先决条件,公路上的车流量大,载重较重,在实际的使用过程中,路面承受巨大的压力,严重影响密实度,机构会产生缝隙,会导致路面出现不同程度的沉降,所以在进行沥青材料配比是都应该保持其具有良好的抗压性能,使路面在使用中不会产生剪切破坏或弯曲疲劳破坏的现象。

图3 香茂乡草甸返青期不同土壤深度的根长密度(a)及径级比例(b)变化特征Fig.3 Variation characteristics of root length density (a) and diameter class ratio (b) at different soil depths in Xiangmao Township Meadow during the greening period
综上,高寒草甸根系的垂直分布大体呈纵向延伸的趋势,且0~25 cm土层分布较为均匀,土壤深处细根占据主导地位。靠近地表处的根密且粗,并随着土壤深度的增加变得疏且细。
2.1.3根系特征参数随海拔与土壤深度的变化 随着海拔的变化,高寒草甸根系生长环境不同,监测点海拔顺序从高到低依次为小唐古拉、措玛乡、香茂乡、那曲大桥,以高寒草甸返青期的数据为例,分析不同海拔下根系形态特征指标随土壤深度的变化情况。
由图4可知,随土壤深度的增加,根系各形态指标参数均呈现了不同幅度的波动。其中在5,10,20,25 cm土深下,香茂乡根系根长、表面积、体积均显著大于同海拔的措玛乡和高海拔的小唐古拉山(P<0.05);而在土深15 cm处小唐古拉山根系长度增加,与香茂乡相比无显著性差异。在25~35 cm深度,香茂乡根系参数均显著大于小唐古拉山(P<0.05)。
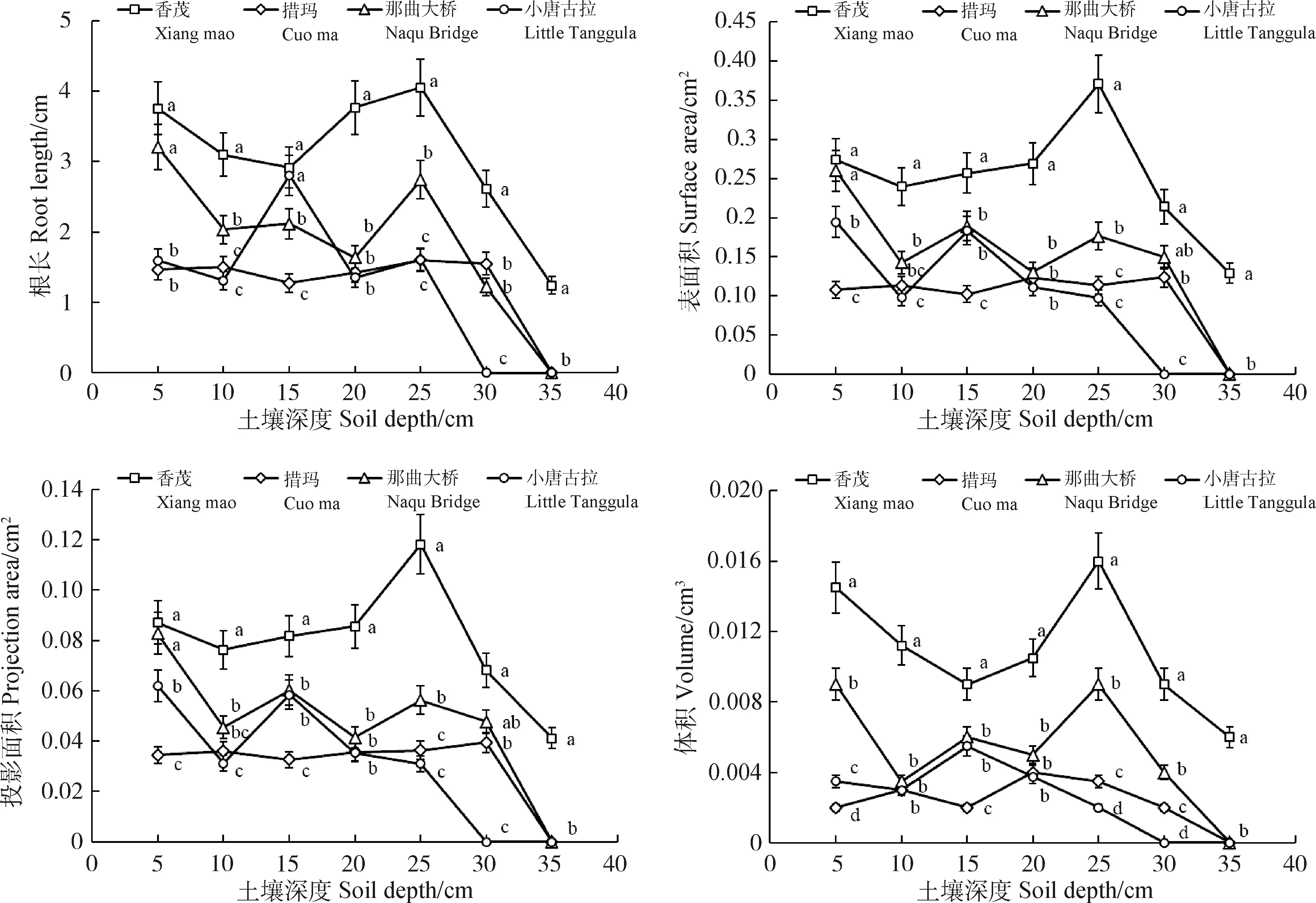
图4 不同海拔下根系形态特征指标随土壤深度变化的关系Fig.4 Relationship between root morphological characteristics and soil depth at different altitudes注:不同小写字母表示同一土壤深度不同海拔间差异显著(P<0.05)Note:Different lowercase letters indicate significant differences between altitude at the 0.05 level for the same soil depth
对不同海拔间根系根长密度进行多重比较(LSD)发现(表2),5~10 cm和20~25 cm土层处,香茂乡根长密度最大,显著高于小唐古拉山和措玛乡(P<0.05)。在15 cm土层处,小唐古拉山根长密度为5.13 m·m-3,占总根长密度的32.3%,所占比例远大于其余三个监测点;在30 cm处,措玛、香茂和那曲大桥的根长密度均显著大于小唐古拉山(P<0.05);35 cm处仅有香茂乡存在根系,占总根长密度的5.79%。根长密度可以反映出植物根系吸收水分和养分的能力。由此可以看出,海拔越高,根系越往土壤较浅层分布,且根系参数呈减少的趋势。

表2 不同海拔下根长密变化特征Table 2 Variation characteristics of root length density at different altitudes 单位:m·m-3
2.2 高寒草甸生长季土壤温湿变化
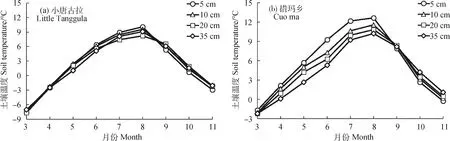
2.2.1土壤温度变化特征 图5为2020年3—11月土壤温度的月动态变化。可以看出,四处监测点0~35 cm土层的土壤温度变化呈先上升后下降趋势,其中小唐古拉山、措玛乡、那曲大桥的土壤温度均在8月达到顶峰,香茂乡土壤温度则是在7月份达到顶峰。在监测期内各点位均是在3月份处于最低气温。其中,海拔最高的小唐古拉山在5 cm土层处达到—7.63℃,比相同土层下措玛、那曲、香茂温度低5.93℃,8.01℃,8.31℃。
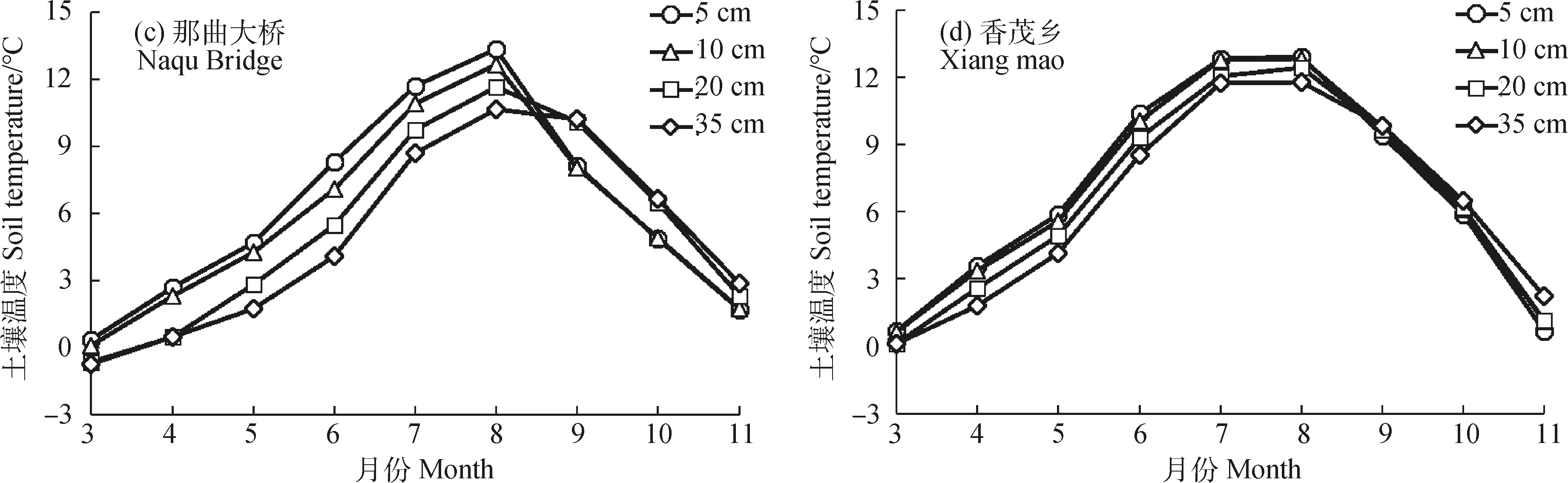
图5 土壤温度动态变化Fig.5 Dynamic changes of soil temperature
从纵向剖面看,3月份天气转暖之后土壤自上而下呈吸热状态,温度随土壤深度的增加而降低。在夏季温度到达顶峰后,土壤温度自上而下呈放热状态,9—11月各监测点浅层土壤温度均小于深层土壤温度。
2.2.2土壤含水量变化特征 3—11月土壤含水量月动态如图6所示。随着时间的变化,四处监测点的土壤含水量均呈现先增加后减少的趋势。其中海拔最高的小唐古拉山在6月份土壤含水量达到最大,措玛、那曲和香茂则是在8月份达到最大。

图6 土壤含水量动态变化Fig.6 Dynamic change of soil moisture
2.3 高寒草甸根系特征参数与土壤温湿特征的关系
利用Canoco 4.5软件对根系参数数据进行趋势对应分析,第一轴梯度最大为1.79(小于3.0),因此选择冗余分析(Redundancy analysis,RDA)来量化不同监测点根系特征参数与环境因子之间的关系。
RDA分析发现,根系特征参数与土壤温湿度之间存在显著相关性。以小唐古拉山为例,前两个排序轴的特征值为0.945和0.013,两轴物种-环境的相关系数分别为0.974和0.698,表明根系特征参数与土壤环境变量之间存在着显著的相关关系。由图7(a)可知,在小唐古拉山返青期土壤含水量对草甸根系分布影响较大,土壤温度次之,且对5~20 cm的土壤表层处影响更强。
在措玛乡(图7b),土壤含水量与土壤温度呈负相关关系,且相对于土壤含水量而言,根系在土壤表层对土壤温度的变化更为敏感。此外,土壤含水量的增加对土壤表层根径大于1 mm的根系生长有抑制作用,对土壤深层根径小于1 mm的根系生长有促进作用,从而进一步影响根系表面积、体积等参数。
那曲大桥(图7c)与香茂乡(图7 d)情况比较相似,土壤温度和湿度呈正相关关系,影响根系参数变化的主要环境因子是土壤温度,土壤含水量主要通过影响根径大于0.5 mm的根来影响根系参数,且这种影响均在土层较浅处相对显著。

图7 根系参数与环境因子RDA排序图Fig.7 RDA sequence diagram of root parameters and environmental factors注:SWC为土壤含水量;ST为土壤温度;SA为表面积;R.D为根径;R.L为根长;R.V为根体积Note:SWC indicate soil water content;ST indicate soil temperature;SA indicate surface area;R.D indicate root diameter;R.L indicate root length;R.V indicate root volume
3 讨论
3.1 不同季节根系变化特征及其影响因素
高寒草甸生长前期各土层的温度自上而下呈减少的态势(图5),土壤表层温度较高,冻土开始消融,加上往年根系生物量的累积,草甸根系的水分和养分得以大量的供应[19],开始进入快速发育阶段,根系集中于土壤表层,呈T字形分布[20]。而生长末期则正好相反,表层较低的土壤温度抑制了草甸根系的生长,使其进入生长停滞期,但由于根系已经过一个生长季的发育,有大量的细根自然分解[21],这就使得草甸表层根系此时的径级较粗,根长减小;而草甸生长前期的根系状态则是径级较细,根长增大。本研究中(图3)返青期和枯黄期的根系参数在土深5~10 cm处的结果也验证了这一结论。随着季节的变化,土壤会吸热和放热,不同深度土壤温度产生变化。同时土壤含水量也在不同的土壤深度出现水分高值区(图6),这是外界温度、降水、日照时长等因素共同决定的,这些变化也间接影响了根系的分布特征。在整个生长季,根系在雨季生长迅速,在旱季生长缓慢[22],且土壤含水量的变化可能造成土壤深度的根系参数出现突变情况,刺激根系的生长,这与本研究中土壤深度20~25 cm处根系特征参数发生变化的情况一致。
3.2 不同空间根系变化特征及其影响因素
根系的空间结构是反映植物地下部分协调生长的重要指标[23]。本研究中,根长密度随着土壤深度的增加呈先下降后上升再下降的趋势,并且约有80%的根系分布在25 cm以上的土层,根长密度在不同土层间的大小可以反映出土壤中水分和养分的分布格局[24],香茂乡5月份的土壤含水量也是在土壤较浅层高,较深层低,这与根长密度变化情况相对应。梅莉等[25]也发现土壤养分在剖面中的分布情况影响了根系的垂直分布,养分含量随土壤深度逐层递减,尤其是土壤氮含量和土壤有机质含量下降速度最快。同时,本研究发现高寒草甸0~0.5 mm的细根在土壤中的占比较高,并随土壤深度的增加比例增大,呈纵向发散趋势。徐满厚等[26]在研究增温对根系的影响中指出,增温使高寒草甸根系生物量向土壤深处纵向发展,细根占比增加,这是为了增加根系与土壤接触的表面积。由此表明0~0.5 mm径级的细根对高寒草甸根系系统贡献最大。
海拔的不同会引起土壤结构、温度、湿度等各方面的变化[27-28]。侯晓娟[29]在研究武功山草甸根系时指出,植被根系特征参数随着海拔的升高有增大的趋势。本研究发现,随着海拔的升高,根系特征参数却有减小的趋势,海拔最高的小唐古拉山根系各参数在大部分土层处显著小于香茂乡,这可能是由于高海拔所引起长期的低温抑制了根系对营养物质的吸收[30],并且2 000℃的积温等值线在海拔5 000 m左右的位置发生偏移[31],导致小唐古拉山植被覆盖稀疏,退化程度高,根系发育较差。阿的鲁骥[32]在研究积雪对高寒草甸根系生长的影响中指出,高海拔处积雪量增加过多,会引起地下储冰量的增加,土壤温度过低同样会抑制根系的生长,这与本研究结论一致。在15 cm土层处,海拔最高的小唐古拉山区域根长密度百分比与其它监测点差异显著(P<0.05),这可能是因为在这种冻融低频模式下,土壤内液态水向融化锋面汇集[33],随着土壤深度的增加,10~20 cm处土壤含水量增加(图6a),刺激了根系的生长。此外,海拔最低的那曲大桥以及与香茂乡同海拔的措玛乡在根长密度百分比之间无显著差异,这可能是气象、土壤环境等多方面因素共同造成的,特别是低海拔处较高频率的冻融循环可能会破坏土壤团聚体大小,改变土壤的物理性质,导致土壤持水和导水能力的下降[34],从而影响到了根系的生长和发育。
4 结论
研究结果表明,根系的变化具有较明显的季节特征,海拔、季节对高寒草甸根系形态分布特征的影响主要是通过土壤水热变化来调控的。不同监测点的土壤含水量与土壤温度的差异性会导致高寒草甸根系采取不同的生长策略以适应环境变化,约有80%的根系分布在0~25 cm土层,并且海拔越高,根系越往土壤较浅层分布。此外,本文未将放牧因素对土壤温湿度以及根系分布的影响考虑进去,在后续的工作中还需进一步研究和探讨。本研究对那曲流域草地生态修复提供了一定的理论依据。
