城市景观格局对鸟类群落的影响: 以南京溧水区为例
2022-03-27张文文李春林
朱 光,王 雪,张文文,吴 翼,雍 凡,李春林,崔 鹏①
(1.安徽大学资源与环境工程学院,安徽 合肥 230601;2.生态环境部南京环境科学研究所,江苏 南京 210042)
随着城市化发展,人类对土地的利用方式发生了巨大变化,城市景观中土地覆被格局也受到很大影响[1]。据估计,到2025年,城市人口将达到50亿,占世界总人口的65%[2]。城市人口增长导致城市生态系统扩张和野生动物栖息地减少,野生动物生存环境受到巨大威胁[3]。大规模城市化对鸟类觅食、栖息和繁殖产生巨大压力,进而可能影响其群落组成。
保护野生动物资源是生物多样性保护的重要环节。如何在人类与野生动物共存环境中实施有效保护措施,是生物多样性保护面临的巨大挑战之一[4]。鸟类是对栖息地变化最敏感的动物类群之一,并且容易调查和观测,因此,鸟类常被用作量化城市生态系统健康的重要指标[5-6],鸟类保护已经逐渐成为城市生物多样性保护的重点[7]。
在城市景观中,土地覆被格局与鸟类栖息地有着密切联系[8]。城市景观可为鸟类提供重要栖息地,城市化通过影响城市景观中的土地覆被格局,进而影响鸟类多样性空间格局[9]。不同土地覆被格局对鸟类多样性的影响有所差异,其中,植被组成和结构对鸟类多样性具有显著影响[10]。城市化通常会使自然景观变成碎片化斑块,城市绿地成为许多鸟类重要庇护所,城市绿地在为鸟类群落提供适宜栖息地方面发挥着重要作用[11]。城市建设用地的增加对土地覆被格局产生很大影响,鸟类可用栖息地正逐渐减少[12]。由于人类对城市鸟类保护意识提高,多数常见鸟类已经习惯于人类存在,对城市环境具有较强适应能力[13-14]。然而,城市土地覆被格局对鸟类群落的影响仍不甚明确。
以城市景观的鸟类群落为研究对象,分析土地覆被格局对不同季节鸟类群落物种多样性和食性集团的影响,以期了解城市鸟类物种数、个体数、多样性指数和均匀度指数与各种土地覆被格局面积占比和景观多样性指数之间的相关性,以及土地覆被格局对鸟类群落食性集团组成的影响。研究结果可为城市鸟类保护提供科学依据。
1 研究区概况
溧水区(31°23′~31°48′ N,118°51′~119°14′ E)位于江苏省南京市中南部,面积为1 067 km2,最高海拔为368.5 m(图1)。研究区属北亚热带季风气候区,四季分明,年平均气温为16.4 ℃,年平均降水量为1 037 mm[15]。研究区在地理上属于典型低山丘陵地区,地势东高西低,丘陵面积占全区总面积的72.5%。区内森林覆盖率约为34.2%,植被分为自然林地、草甸植被和人工栽培植被。自然植被包括针叶林、落叶阔叶林、落叶与常绿阔叶混交林和竹林等;栽培植被包括大田作物、蔬菜作物、经济林和果园等。
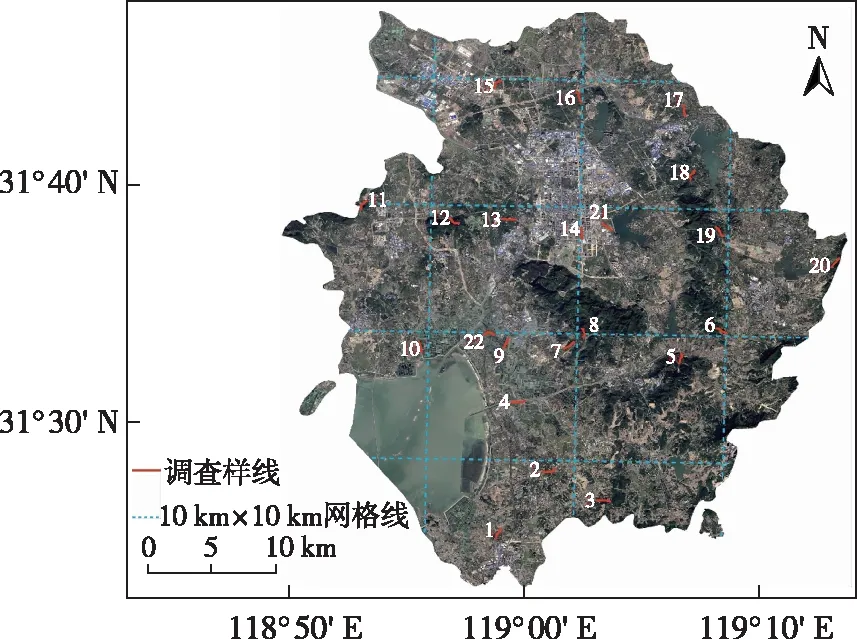
图1 研究区示意
2 材料与方法
2.1 样线布设
根据溧水区生境特征和用地类型分布格局,在研究区域内布设22条长度为1 km的固定调查样线(图1)。样线主要涉及林地、村庄、农田、草地和水库等生境。为了确保调查范围覆盖全区,在溧水区遥感影像图上绘制10 km×10 km网格线,在面积大于25 km2的区域内均布设样线。
2.2 鸟类调查
鸟类调查在2019年春(4月)、夏(8月)、秋(10月)和冬(1月)4个季节进行。调查频次为每季度1次。选择晴朗、风力较小的天气,在鸟类活动较频繁的时间段(日出后3 h和日落前3 h内)进行调查。野外调查时,2名经验丰富的观察员手持双筒望远镜(SWAROVSK 10×42),以大约2 km·h-1的速度沿样线前进,记录样线左右两侧各50 m范围内看到或听到的鸟类种类及数量。为避免重复记录数据导致的误差,不记录从身后向前飞行的鸟类。使用《中国鸟类野外手册》[16]对所观察到的物种进行鉴定,鸟类居留型分类依据《中国鸟类分类与分布名录》(第3版)[17]。此外,按照食性,将鸟类划分为5类[18-19]:(1)食虫鸟类,以昆虫及其幼虫为食;(2)杂食性鸟类,兼食昆虫和植物;(3)食果鸟类,以植物花蜜果实为食;(4)食谷鸟类,以植物种子和粮食作物为食;(5)肉食性鸟类,以哺乳动物、脊椎动物和大型无脊椎动物为食。该研究只对林鸟进行分析,不涉及水鸟。
2.3 土地覆被数据
在每条样线两侧各100 m范围内量化土地覆被格局及景观多样性,作为影响鸟类群落的环境因子[20-21]。利用LocaSpace Viewer软件对高分辨率遥感图进行判读,量化各种土地覆被类型面积。土地覆被类型分为农田(水田和旱田)、水域(河流、水塘和沼泽)、林地(有林地和灌草丛)、草地(低矮植被)和建设用地(居住地和交通用地)5类。景观多样性指数(H1′)[22-23]可反映景观要素的多少和比例的变化,其计算公式为
(1)
式(1)中,pi为第i种土地覆被类型面积占总面积的比例;m为每条样线土地覆被类型总数。
2.4 数据分析
采用物种数、个体数、Shannon-Wiener多样性指数和Pielou均匀度指数量化每条样线的鸟类物种多样性。
Shannon-Wiener多样性指数计算公式为
(2)
式(2)中,H2′为Shannon-Wiener多样性指数;Pi为第i个物种个体数占每条样线总个体数的比例;S为每条样线记录到的物种数。
Pielou均匀度指数(J)计算公式为
J=H2′/lnS。
(3)
按照不同物种个体数占鸟类总个体数的比例确定优势度(P):P≥0.100,优势种;0.010≤P<0.100,常见种;0.001≤P<0.010,稀有种;P<0.001,罕见种[24]。
采用向后逐步回归法分析环境因子(各土地覆被类型面积占比和景观多样性指数,连续变量)和季节(春、夏、秋和冬季,分类变量)对鸟类多样性指数的影响。首先采用Kolmogorov-Smirnov检验对各鸟类群落多样性指数进行正态检验,并通过对数转换和平方转换使其逼近正态分布。以转化后的鸟类多样性指数为响应变量,以环境因子和季节为解释变量进行向后逐步回归分析。
采用约束排序法分析鸟类集团与环境因子之间的关系。鸟类集团和环境因子均为多元数据,需要采用无趋势对应分析(DCA)方法对食性集团个体数进行探索性分析,以确定是否可以采用线性或单峰排序方法进行分析。鸟类食性集团个体数的DCA最大梯度长度小于3.0,故选用冗余分析(RDA)方法。RDA方法是基于对应分析发展而来的一种排序方法,将对应分析与多元回归分析相结合,能够分析环境变量与物种数据两两之间的关系。对变量进行逐步筛选的过程中,将共线性自变量从RDA模型中移除。采用蒙特卡罗检验评估模型显著性,只展示P值小于0.05的模型。采用RDA模型中调整后的R2来解释所选变量对方差的贡献,采用经调整后的R2乘以RDA排序轴的方差解释率作为各排序轴的实际方差解释率。
在进行向后逐步回归分析和冗余分析时,需要采用方差膨胀因子(VIF)来检验环境变量的共线性,将共线性很强的水域面积占比剔除后,所有环境变量的VIF值均小于4。经共线性变量筛选后,以上分析中涉及的环境变量有5个,分别为草地面积占比、林地面积占比、农田面积占比、建设用地面积占比和景观多样性指数。
以上所有数据处理采用R 3.6.3软件完成。
3 结果与分析
3.1 土地覆被格局
在研究区域内的22条调查样线中,林地面积占比较高的样线有11条,占比平均为38%;农田面积占比较高的样线有7条,占比平均为19%;水域面积占比较高的样线有3条,占比平均为12%;建设用地占比较高的有1条,占比平均为19%(表1)。
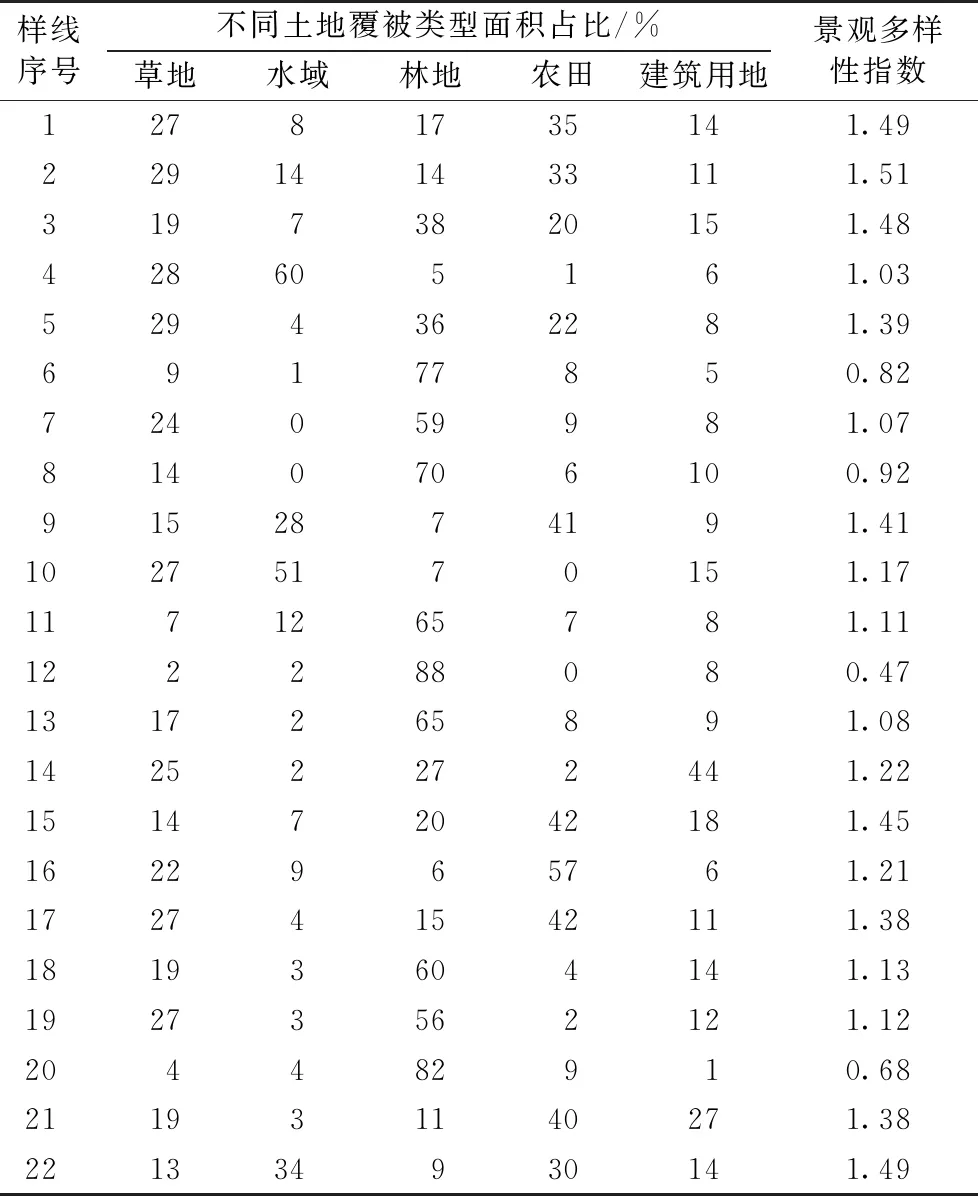
表1 溧水区鸟类调查样线两侧各100 m范围内土地覆被
3.2 鸟类物种组成

按居留型进行划分,夏候鸟、旅鸟各有9种,冬候鸟有18种,留鸟有46种,分别占全部个体数的3.8%、0.9%、17.3%和78.0%;根据优势度指数对鸟类进行划分,优势种有3种,常见种有19种,稀有种有27种,罕见种有33种,分别占鸟类总个体数的40.5%、48.3%、10.3%和0.9%。
3.3 鸟类多样性指数
如图2所示,不同季节鸟类物种数和多样性指数均存在显著差异,春季和秋季鸟类物种数显著大于冬季;春、夏和秋季多样性指数均显著大于冬季;春季均匀度指数显著大于冬季;不同季节鸟类个体数无显著差异。如表2所示,鸟类物种数与农田面积占比、林地面积占比和景观多样性指数呈显著正相关(F6,81=4.175,P<0.01);鸟类个体数与草地面积占比呈显著负相关,与景观多样性指数呈显著正相关(F2,85=4.355,P<0.01);鸟类多样性指数与农田面积占比呈显著正相关(F4,83=5.680,P<0.001)。
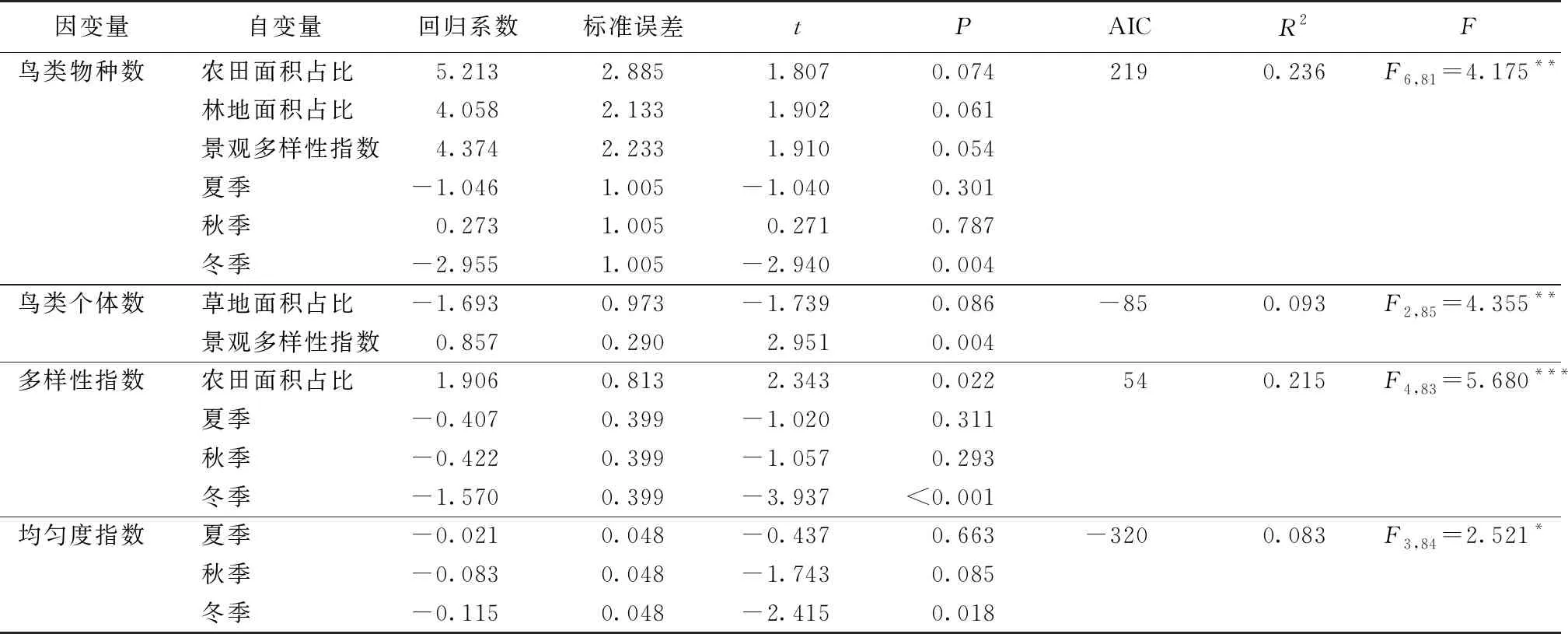
表2 鸟类多样性指数与环境因子和季节的回归分析结果
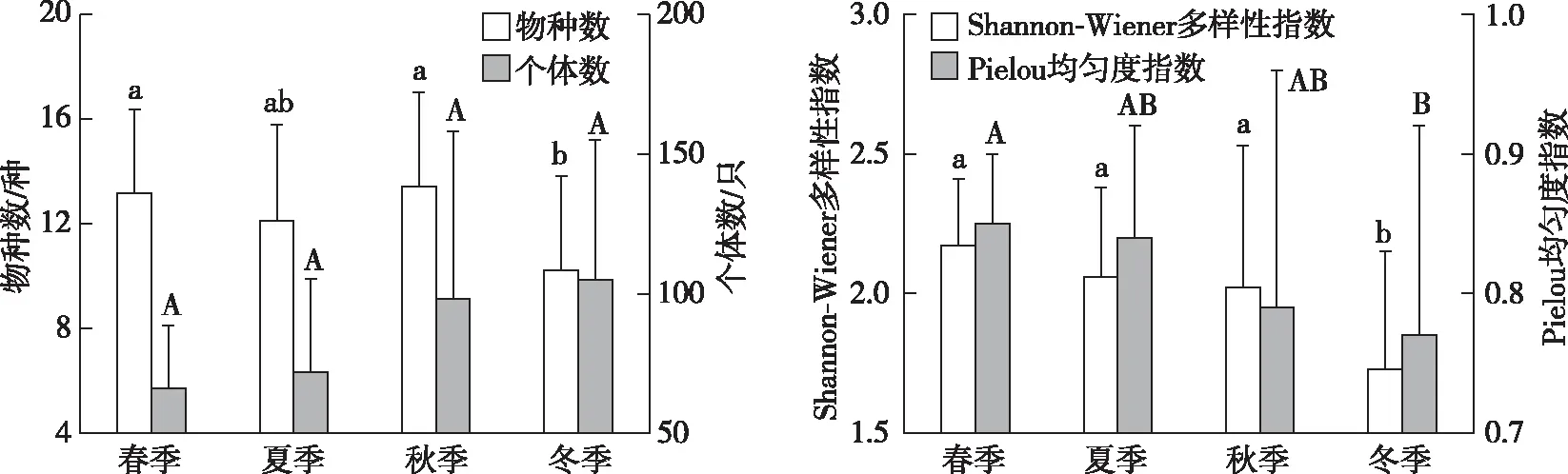
同一幅图中,直方柱上方英文大写或小写字母不同表示不同季节间某指标差异显著(P<0.5)。
3.4 鸟类食性集团结构
如表3所示,采用冗余分析方法分别对各个季节鸟类食性集团与环境因子进行分析,在所得到的4个季节的RDA模型中,春季(P=0.002)、夏季(P=0.001)、冬季(P=0.002)模型具有显著性,前2个排序轴的累计解释率依次为28.8%、27.6%和23.8%。秋季RDA模型不显著(P=0.326),前2个排序轴的累计解释率为15.3%。
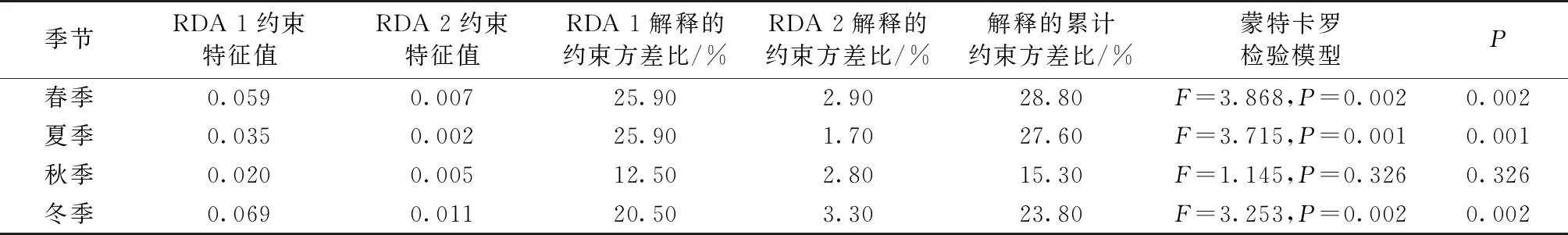
表3 鸟类食性集团与环境因子的RDA分析结果
如图3所示,春、夏和冬季杂食性、肉食性和食谷鸟类数量与草地面积占比呈正相关,食虫和食果鸟类数量与森林面积占比呈正相关;春和夏季杂食性、肉食性和食谷鸟类数量与景观多样性指数呈正相关;冬季杂食性鸟类数量与建设用地面积占比呈正相关;秋季鸟类食性集团个体数与环境因子之间相关关系不显著(图3)。
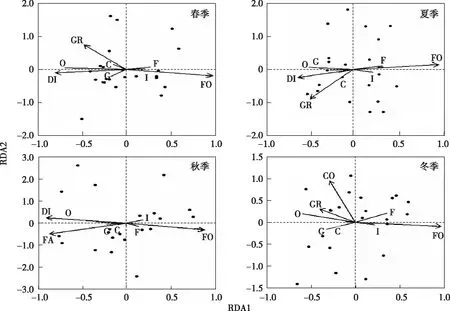
GR为草地面积占比,FA为农田面积占比,FO为林地面积占比,CO为建设用地面积占比,DI为景观多样性指数,F为食果鸟类,G为食谷鸟类,I为食虫鸟类,O为杂食性鸟类,C为肉食性鸟类。
4 讨论与结论
土地覆被组成的不同景观能够为鸟类提供的食物资源有所差异,这些景观对鸟类多样性有显著影响。鸟类个体数和种类与景观多样性存在密切联系,复杂的景观格局有利于提高鸟类物种多样性[6,10]。
在城市景观中,土地覆被格局通常被认为是对鸟类多样性影响最显著的因素之一[10]。笔者研究结果表明,景观多样性指数的提高有利于鸟类物种数和个体数的增加,这与其他城市鸟类的研究结果[7]一致。复杂的景观结构可为鸟类提供更宽阔的生态位和丰富的食物资源,更有利于鸟类生存[2,25]。农田生境为鸟类提供了优越的觅食条件,例如,农田中的谷物、种子和昆虫等,都是能够吸引鸟类的食物资源[11,26-27]。有研究[25]表明,农田面积扩大,对鸟类物种数和多样性增加有积极影响,笔者研究也证实了这一点。在城市景观中适当增加农田景观异质性,提高农田边缘植被复杂度,有利于增加农田鸟类多样性[25]。由于林地景观植被结构复杂,能够为鸟类提供充裕的食物资源和理想的筑巢条件,因此林地景观被认为是最能够吸引鸟类的景观之一[28]。研究区林地面积占比较高的景观中,鸟类物种数也较多。然而,草地面积增加不利于鸟类个体数增加,这可能是由于城市草地植被类型单一,食物资源匮乏[25]。
土地覆被格局不仅对城市鸟类群落物种多样性产生影响,而且对其食性集团组成也有显著影响。在景观多样性指数较高的区域中,有更多的杂食性、肉食性和食谷鸟类。复杂的生境为鸟类提供更多的觅食机会,能够吸引多种食性的鸟类[29]。林地为鸟类提供了理想的庇护所、筑巢点和觅食环境,是鸟类栖息的重要场所[30-32]。在城市景观中,林地是果实和昆虫的重要分布区域,所以能够吸引许多食果和食虫鸟类。草地和农田景观比较开阔,有利于肉食性鸟类的觅食活动,并且这2类景观中的昆虫和谷物,可以吸引较多的杂食性和食谷鸟类[33-35]。随着季节更替,食物资源会发生明显变化,冬季食物资源较为匮乏,鸟类更倾向于依赖人类活动产生的剩余食物[2]。在建设用地区域,频繁的人类活动产生了较多的垃圾和遗弃的食物,可以为杂食性鸟类提供食物资源[14,36]。因此,在建设用地面积占比较高的区域,冬季记录到的杂食性鸟类较多。杂食性鸟类在草地、农田和建设用地等景观中的数量均较多,食虫和食果鸟类仅在林地中数量较多。与食虫和食果等鸟类集团相比,杂食性鸟类适应性强,能够在多种生境中占据优势地位[18-19]。
该研究结果表明,土地覆被格局对城市鸟类群落物种多样性和食性集团组成具有显著影响。土地覆被格局是预测鸟类群落的关键环境因子,各种土地覆被类型对鸟类选择觅食场所和栖息场所均具有重要影响。在城市景观中,较高比例的林地和农田景观有利于鸟类物种数提高,同时,较高景观多样性也是提高鸟类多样性的关键因素。为维持和保护城市鸟类群落物种多样性,建议在城市规划和管理中,增加土地覆被格局的多样性,提高林地面积占比,丰富草地环境异质性;在城市鸟类食物匮乏的冬季,在鸟类经常活动区域,适当增加人工投食,可以为鸟类生存提供食物保障。
