马尾松混交林的超产效应及其影响因素
2022-03-22林润东张进如李石一宁姚晓东王小红陈光水
林润东,张进如,李石一宁,姚晓东,王小红,陈光水*
(福建师范大学 a.湿润亚热带山地生态国家重点实验室培育基地,b.地理科学学院,福州350007)
中国亚热带地区是中国乃至全球最重要的人工林基地之一。中国第9次森林资源清查(2014—2018年)结果[1]显示,全国森林面积达到2.20亿hm2,森林覆盖率为22.96%,其中人工林面积达0.80亿hm2,占林地面积的36.45%,占全球人工林总面积的33%,人工林面积全球排名第一。其中马尾松(Pinusmassoniana)人工林是中国人工林重要的组成部分,面积达0.6×107hm2,是主要的松树人工林树种,耐干旱、耐贫瘠、适应性强,是水土保持的主要造林树种[2],在林业生产和森林碳汇中发挥着重要作用。然而,90%的马尾松是以单一人工林进行栽培的,由于自然或人为的因素,往往会引起包括土壤肥力和生物多样性的降低、对环境干扰的耐受程度低、线虫等病虫害频繁发生、火灾风险高等一系列生态问题[2]。因此如何科学营造人工林,在兼顾生产力增加的同时能够更好地满足生态系统的服务功能需求,是当前中国林业生产所急需解决的重大问题。
相比之下,许多研究表明,营造混交林是提供更多样化的生态服务和林产品的理想方法[3]。然而混交林是否可以增加林分生产力一直是一个核心问题,混交林是否能够超产(Overyielding),即混交林的生产力是否比各自的纯林的预期生产力高,目前仍存在争议。因此很有必要对混交林超产效应进行系统的meta分析。目前国际上已有很多学者对混交林的生产力进行大尺度的meta分析,均发现存在不同程度的超产效应[4-6],并且会因树种组成[7]、树龄[8]、混交比例[9]、气候因子[10]等不同林分和环境影响因素的差异而产生很大的变化。
然而目前研究都主要集中于处在温带和寒温带地区的欧洲国家的少数几个树种组合,同一个目标树种与多样化树种混交林混交效应的meta分析仍然缺乏,并且有关亚热带地区混交林的超产效应研究很少,特别是缺少以马尾松等中国特有的乡土树种为目标树种的混交林的综合分析。由于亚热带与温带的水热条件和树种组成存在显著的差异,温带地区混交林的超产结论能否在亚热带地区的混交林得到验证仍缺乏相关研究。此外已有大多数混交林对生产力的影响研究主要基于蓄积量(树干)的评估,而未能准确体现其他器官如根、枝以及叶的贡献。这些器官占了生物量和生产力的很大比例,很可能会影响超产的结果以及对超产原因和机制的解释[11],且不同混交林中树种间的相互作用引起的不同器官生物量分配比例变化,也会影响其超产效应的评估[12],因而仅仅基于蓄积量的评价指标可能对评估混交林是否能够超产带来一定的不确定性。
作为中国南方主要的造林树种,马尾松造林面积大,是重要的森林资源。随着林业发展战略的转变,作为中国森林面积排位第5的优势树种,大部分马尾松林的功能定位已由商品林转为生态公益林,传统的马尾松林经营模式已无法满足这一定位转变的需要,必需做出相应的调整[13],所以对马尾松混交林的超产效应及其影响因素进行meta分析十分有必要。本研究所要回答的科学问题有:(1)马尾松混交林的生产力是否能够获得超产效应?(2)马尾松混交林是否会影响马尾松各器官生物量的分配?(3)树种组成、树龄、混交比例、气候因子、小地形、混交树种高度差等因素是否会影响超产效应?由于乔木层总生物量可代表不同树种对资源利用、碳分配和碳固持的贡献,且包括了树木的各个器官,因而可以作为比蓄积量更好评价混交林生产力的关键指标。本研究以森林乔木层总生物量和各器官生物量作为生产力的评价指标,揭示马尾松混交林生产力的超产效应及其影响因素。研究结果可为科学评估马尾松混交林的生态和经济效益提供科学依据,进而为中国马尾松林的可持续经营发展以及混交林的培育和管理提供借鉴和参考。
1 研究区与数据采集
1.1 研究区概况
数据来源于中国知网和Web of Science数据库。以中文关键词“马尾松”“混交林”“生物量”“生产力”“生产量”,在中国知网和万方数据库中检索至2020年的期刊论文或学位论文;同时以相同的英文关键词在Web of Science中检索至2020年的期刊论文。对检索出来的论文按以下条件进行进一步筛选:(1)研究中必须同时包括马尾松混交林及其对应纯林(至少要有马尾松纯林);(2)混交林应包括各树种树高、胸径、总生物量或各器官生物量,或可以间接计算出生物量的数据;(3)必须为人工种植,混交林和纯林初植密度相同,且立地条件一致;(4)与马尾松混交的树种限定为乔木树种,排除竹类、藤本等;(5)如果同一个混交林试验有不同时期的报道结果,仅选择林龄最大的研究结果。
根据上述的条件进行筛选,最终一共得到26篇文献,包括50个马尾松混交林。同时收集马尾松混交林和纯林的林龄、初植密度、初植混交比例、林分和各树种保留密度、各树种林木生长状况数据;以及气候、土壤类型、小地形等数据。所收集的研究位于福建、江西、广西、广东、安徽、湖南6个省份,福建省最多。混交林试验中,如果同时设计有混交林及混交林中各树种的纯林,称为三元组。本研究中具备完整的三元组(Triplets,即同时包含马尾松混交林、混交林中各树种纯林)的试验共21例。三元组的设计已被证实在定量树种的混交效应上十分有用[14],只有三元组的混交林试验,才可计算出林分水平的超产效应。
1.2 数据分析
采用对数化响应比(lnR)作为评价混交效应的衡量指标。根据不同混交林与纯林的比较目的,采用3种不同的lnR:
(1)马尾松混交林相对于纯林的响应比(lnRp):
lnRp= ln(Xm/Xp)
(1)
式(1)中:Xm为马尾松混交林的变量(混交林林分或某个树种的生物量、树高、胸径、器官分配比例等),Xp为马尾松或混交林中其他树种纯林相对应的变量;下标p和m分别代表纯林和混交林。
(2)林分水平马尾松混交林相对于纯林加权平均值(或预测值)的响应比(lnRw):
lnRw= ln(Xm/Xw)
(2)
式(2)中:Xw为马尾松混交林中某个变量的预测值,为两个纯林的加权平均值(下标w代表加权平均值),由下式计算为:
Xw=Xp, PM×PPM,(OS)+Xp, OS×P(PM),OS
(3)
式(3)中:Xp, PM和Xp, OS分别是马尾松纯林和其他树种纯林的某个变量的值,PPM,(OS)和P(PM), OS分别是混交林中马尾松和其他树种的初植混交比例。下标PM代表马尾松,OS代表其他树种;PM,(OS)代表混交林中马尾松,(PM), OS代表混交林中其他树种。当lnRw>0时,如果变量为生物量,则表示产生超产效应。
(3)树种水平马尾松混交林中某树种相对于纯林加权平均值(或预测值)的响应比(lnRw′):
lnRw′= ln(Xm/Xw′)
(4)
式(4)中:Xw′为马尾松混交林中某个树种某个变量的预测值,为其对应纯林的加权平均值(下标w代表加权平均值),由下式计算为:
Xw′=Xp, PM×PPM,(OS)
(5)
式(5)中:Xp, PM是马尾松纯林某个变量的值,PPM,(OS)是混交林中马尾松的初植混交比例。下标PM代表马尾松,OS代表其他树种;PM,(OS)代表混交林中马尾松。当lnRw>0时,如果变量为生物量,则表示产生超产效应。
在meta分析中,为了对不同研究中的效应值进行整体评估,往往根据不同研究结果中的方差进行加权。但国内已报道的混交林文献中很少报道数据的样本大小和变异数据,因而对不同研究的结果很难根据方差进行加权。所以只能对所有的研究赋予相同的权重,即不进行加权[15],因而本研究中没有对不同研究进行加权。本研究采用非权重平均法来计算出各指标的效应值,效应值的显著性由999次重取样进行检验,95%置信区间通过MetaWin 2.1软件所整合的自助法(Bootstrap)非加权数据来产生。如果不同组间的95%置信区间没有重叠,那么不同组间的效应值具有显著差异。如果置信区间包含0,则没有显著效应。若置信区间全部>0,则有显著增加;反之,则显著减小。同时,由于马尾松与常绿阔叶树种混交较多,与其他生活型混交较少,因而单独对该混交类型进行分析。通过SPSS 26.0分析混交效应与混交林林龄、初植混交比例和密度、林分保留密度、年均温、年均降雨量、海拔、坡度间的相关关系。
2 结果与分析
2.1 林分水平和树种水平马尾松混交林生物量的超产效应
在林分水平上,与预测值(纯林的加权平均值)相比,马尾松混交林乔木层总生物量和各器官生物量的效应值均显著大于0,表现出超产现象(图1)。总体上,乔木层总生物量95%(21个样本中20个lnRw为正值)表现出超产现象,超产44.8%;地上部分生物量89%(18个样本中16个lnRw为正值)表现出超产现象,超产49.4%。树枝生物量超产最多,为57.7%;树根超产最少,为27.3%。
在树种水平上,从图2可见,与林分水平相似,马尾松在混交林中也都表现出了超产效应。总体上,马尾松总生物量90%(29个样本中26个lnRw为正值)表现出超产现象,超产31.3%;地上部分生物量87%(30个样本中26个lnRw为正值),超产36.9%,超产幅度相比林分水平较小。从图3可见,其他树种总生物量和地上部分生物量89%(18个样本中16个lnRw为正值)表现出超产现象,分别超产45.5%和50.6%。虽然与常绿阔叶树种混交,马尾松也基本获得超产效应,但是在林分和树种水平上根生物量都没有获得超产,并且各器官生物量的超产幅度相较于总体水平也减小。从图4可见,马尾松混交林中马尾松平均胸径和平均树高相对于马尾松纯林的响应比lnRp值均显著大于0,分别提高了13.4%和7.3%。其他树种的胸径和树高分别提高了11.3%和7.3%(图5)。这些结果均表明了马尾松混交林可以更好地促进马尾松和其他树种的生长和发育。

注:Total:马尾松总体水平混交,EG-BL: 马尾松与常绿阔叶树混交。杠表示响应值的95%置信区间,数字表示样本数,下同。
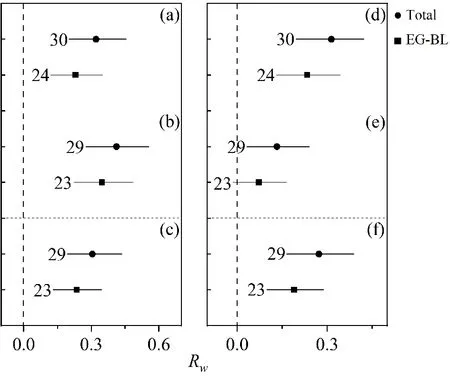
图 2 树种水平上马尾松混交林中马尾松树干(a)、树枝(b)、树叶(c)、地上(d)、树根(e)、总生物量(f)相对于预测值的响应比
2.2 混交林对生物量分配的影响
从图4和图5可见,马尾松在混交时器官生物量的分配比例发生了显著变化,这可能是影响混交林产生超产效应的一个重要因素。马尾松树枝的地上部分生物量分配比例的lnRp值显著大于0,表明树枝的地上部分生物量分配比例的增加,分别增加了13.8%和3.1%;树根生物量分配比例和根茎比的lnRp值显著小于0,表明树根分配比例减少,根茎比降低,分别减少了13.1%和15.7%。而其他树种地上生物量分配增加3.5%,树根生物量分配比例和根茎比分别减小6.4%和9.6%。这些结果指出在混交时树种更倾向于将资源投入到地上部分,尤其是树枝部分。此外,从图4可见,马尾松和其他树种在混交时的干形(树高/胸径2)相对于纯林的lnRp值显著小于0,分别减小了10.9%和7.8%。

图 3 树种水平上马尾松混交林中其他树种树干(a)、树枝(b)、树叶(c)、地上(d)、树根(e)、总生物量(f)相对于预测值的响应比

图 4 马尾松混交林中马尾松树干(a)、树枝(b)、树叶(c)、地上(d)、树根(e)分配比例、根茎比(f)、树高(g)、胸径(h)、干形(i)对于纯林的响应比
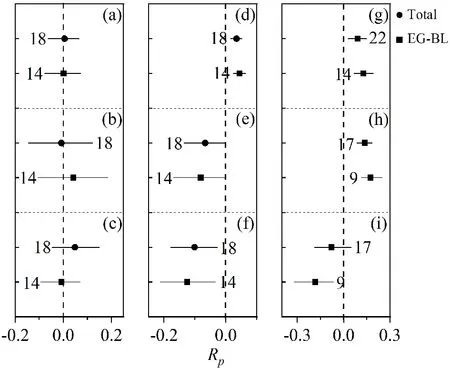
图 5 马尾松混交林中其他树种树干(a)、树枝(b)、树叶(c)、地上(d)、树根(e)分配比例、根茎比(f)、树高(g)、胸径(h)、干形(i)对于纯林的响应比
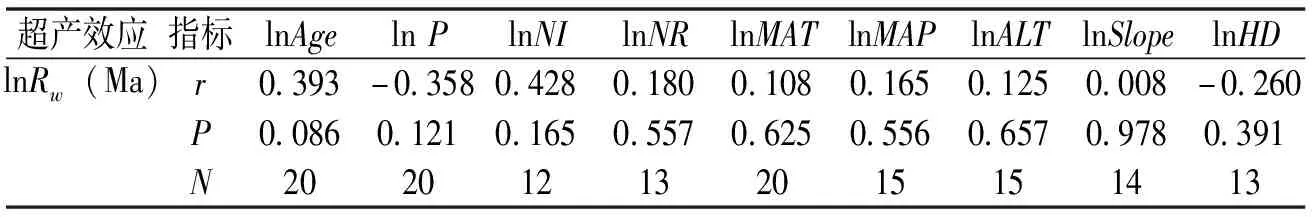
表 1 马尾松混交林的超产(lnRw(Ma))与混交林的林龄(Age)、初植比例(P)、初植密度(NI)、保留密度(NP)、年均气温(MAT)、年均降雨量(MAP)、海拔(ALT)、坡度(Slope)、高度差(HD)的相关关系
2.3 马尾松混交林生物量超产效应的影响因素
从表1可见,马尾松混交林的超产与林龄有边缘显著效应,表明与林龄正相关。然而马尾松混交林的超产效应与初植密度与比例、保留密度、气候因子、小地形、高度差等因素均没有显著的相关性,表明在一定的条件范围内,马尾松混交林的超产效应不受各种环境因素的影响,更多是由于受到混交树种相互作用后,自身资源竞争策略改变而造成。
3 讨论
3.1 马尾松混交林的超产效应
本研究发现,总体上马尾松混交林乔木层生物量超产44.8%,地上生物量超产49.4%,超产的幅度超过已有的一些报道。Hulvey等[4]对8个研究(其中5个位于热带、3个位于温带)中的48个混交林的meta分析表明,混交林的碳贮量(以地上生物量作为近似指标)平均可超产38%。Pretzsch和Schütze[16]对欧洲中部42个三元组(包括欧洲赤松,挪威云杉, 花旗松和欧洲山毛榉4个树种的两两混交组合,共126个混交林和纯林样地)进行了meta分析发现,混交林的林分蓄积量可超产3%~16%。Pretzsch和Forrester[17]基于长期试验和资源清查数据的研究表明,温带和寒温带地区商品林树种混交的蓄积量超产可达10%~30%。Jactel等[11]利用分布在世界各地的60个样地上的126个混交林试验数据进行meta分析发现,总体上混交林的生产力比各自纯林的平均值高15%。Jonsson等[6]利用2012个挪威云杉、欧洲山毛榉和桦树(Betulapendula)两两混交和纯林样地进行整合分析发现,与纯林相比,35%的混交林生物量可以超产,而只有9%的混交林低于纯林。
然而这些研究都主要集中在处在温带地区的欧洲国家以及少部分热带地区,有关亚热带地区混交林对生产力影响的报道仍然很少,并且可以看出相较于温带各树种混交林的超产效应的meta分析所得出的结果,处于亚热带的马尾松混交林有着更大幅度的超产。
由于马尾松与常绿阔叶树种混交数据较多,单独对其进行分析,虽然其与总体上的超产效应基本相似,但是超产幅度降低,并且在根生物量上没有响应,表明不同树种组成对马尾松的超产效应机理可能存在差异。
马尾松为强阳性树种,不耐庇荫,喜光,而常绿阔叶树种大部分为中性或具一定耐阴性,两者对光的需求存在差异。所以马尾松在与常绿阔叶树混交时,如何处理好林冠的垂直结构是其能否超产的关键。混交林中马尾松与常绿阔叶树的相对树高差异,会对马尾松的超产效应产生影响。如果马尾松树高大于混交树种,位于林冠最上层,可以满足其对高光的需求,影响超产效应;而当前者矮于后者时,位于林冠下层,则可能限制了其对光的需求,从而超产效应降低。在本研究中,与常绿阔叶树混交时只有50%的马尾松树高大于混交树种,这可能是超产幅度相较于马尾松总体水平降低的原因。Zenner等[18]也指出,当耐荫树种和阳性树种的混交组合,只有在阳性树种树高大于耐荫树种,且能在林分发育过程中一直得以维持时才能发挥互补性。而混交时其根生物量没有发生超产效应的原因很可能也是因为生物量更多地分配到地上,去竞争光照的结果。
3.2 马尾松混交林对生物量分配的影响
目前虽然有关混交林对森林生产力影响的研究已引起了越来越多的关注,但混交效应的内在机制仍没有被很好地理解。林冠生长空间的互补利用以及生长分配[19]被认为是一个重要的影响因素。在本研究中,也发现了马尾松器官分配的改变和形态的变化。在总体水平上,虽然马尾松混交林中地上和地下部分都表现出了显著的超产效应,但是相比于纯林,马尾松在混交时更倾向于将更多的生物量分配在地上部分,尤其是树枝部分,而树根的分配比例降低,总体上根茎比降低。根据最优分配理论[20],混交林中树种生物量的分配策略可以根据资源的可利用性进行调整,林木生物量的分配具有可塑性,混交林中不同树种之间的相互作用会影响不同器官的分配比例,以提高对最限制生长资源(光、水分、养分)的获取[23]。冠幅一般与胸径(或胸高断面积)呈正比。混交林中树枝分配比例增加,可能意味着冠幅增加。若冠幅增加,胸径也需要增大,才能保证水力的平衡。
对于地上部分生物量来说,马尾松为强阳性树种,不耐庇荫,而光照又是光合作用的潜在限制性资源[8]。在混交时马尾松将更多的生物量分配给树枝,可以更好地调节冠层空间和物理生态位划分,以调节自我遮蔽的程度,改变冠层内部的光照条件(即光照质量和光照数量),允许更多的叶面积进行光的捕获,提高光的利用效率,进而提高马尾松的可利用资源数量。而树干变粗也是为了支撑更大的树冠并且提高养分、水分运输效率,所以马尾松优化了自身的结构,在有效利用树冠空间和机械稳定性要求之间找到权衡。Guillemot等[21]在巴拿马中部对6种16年生热带树种的混交和纯林进行激光扫描和破坏性取样研究发现,树的生物量分配和结构受到树种间的相互作用(可塑性效应)的影响,生物量在树枝间分配的增加解释了所观察到的大量超产效应,并且混交树种往往具有较高的木质生物量和较大的树干直径,试验得出地上部分的超产和树枝结构的可塑性会相互影响,使生物多样性高的林分中的树木能够更均匀、更完整地占据树冠的空间。这也与先前对林龄较低的温带和亚热带森林的研究发现混交林中结构互补性增加的结论类似,所以混交林地上生物量的超产和分配间的正反馈对生态系统产生了积极的影响。
对于地下部分生物量来说,根生物量分配减小可能与混交林中树种对地下生物量的中性响应[22]有关。马尾松更倾向于优化获取器官(即细根)的特征和支持器官(即粗根)的空间构型来强化对地下资源的获取。Martin-Guay等[23]通过对位于加拿大魁北克的研究农场的14个两物种混交、10个四物种混交以及纯林的研究进行分析,也发现树种细根和粗根的生物量没有产生超产效应,并且发现对于地下资源获取效率的提高可以减少对地下生物量分配的需求,同时导致地上部分更大的超产效应。
3.3 树龄、气候、地形等其他因素对马尾松混交林超产效应的影响
本研究发现马尾松混交林的超产效应与林龄有边缘显著效应,表明与林龄正相关,这与已有的一些报道相似,如Thurm和Pretzsch[8]通过对德国不同生态区域、不同年龄(30~120年生)的18个混交林的三元组的研究表明,混交林的超产效应随林龄增加而增加。Mina等[10]发现混交树种间的互补作用随林分发育阶段的增加而增加。然而Lu等[24]的研究没有发现林龄对两种常绿针叶树-落叶阔叶林混交林(花旗松-山毛榉和欧洲赤松-橡树)的超产效应有显著影响。本研究所收集到的马尾松混交林主要为幼龄到近成熟林,因而还无法代表林龄更大的、更为成熟的马尾松混交林的生产力特征。同时,本研究所报道的混交效应仅代表特定马尾松混交林的某个林分发育阶段;混交林中树种间的竞争关系可能随林龄的变化而变化,因而还无法探知同一个混交林在林分发育过程中混交效应的变化规律。
然而超产效应与初植密度与比例、保留密度、气候因子、小地形等因素无关。Pretzsch等[25]的研究表明,虽然其研究的混交林的降雨量、气温等有很大梯度,但其超产效应与气候因子也均不相关。
4 结论
通过meta分析表明,整体上,马尾松混交林的乔木层生物量和地上生物量显著高于纯林,林分水平乔木层生物量超产44.8%,地上生物量超产49.4%,树种水平马尾松总生物量超产31.3%,地上生物量超产36.9%,树高和胸径相比纯林也得到了提升,这表明马尾松混交林可以同时促进林分生产力和马尾松的生长,并且混交会改变树种生物量的分配比例,混交时马尾松的树枝生物量分配比例增加,根生物量分配和根茎比减小,树高/胸高断面积比减小。与林龄呈正相关,但是和初植密度与比例、保留密度、气候因子、小地形等因素没有关系,可能与混交林改变资源竞争策略有关。未来研究应进一步深入揭示马尾松混交林超产的主要机制,并提出优化的马尾松混交林树种组合和栽培管理措施。
