青海菜用蚕豆改良效果评价及主要数量性状遗传效应分析
2022-03-21周仙莉张红岩滕长才刘玉皎
林 夕, 周仙莉, 张红岩, 滕长才, 刘玉皎,3
(1.青海大学, 西宁 810016; 2.青海省农林科学院, 西宁 810016;3.青海大学省部共建三江源生态与高原农牧业国家重点实验室, 西宁 810016)
蚕豆(ViciafabaL.)是世界四大食用豆之一,是集粮、菜、饲、绿肥兼用的豆科作物,具有十分重要的商业价值和营养价值[1-3]。菜用蚕豆是我国蚕豆产业的重要结构组成部分,主要分布在江苏、浙江、上海等省(市)。随着蚕豆产业结构调整和人类饮食结构的多元化发展,云南、重庆、四川、甘肃、青海等区域以“干改鲜”模式逐步调整和优化蚕豆产业结构,逐步改变我国蚕豆以粮为主的单一产业结构,构建了多元化产业发展模式。选育专用蚕豆品种是推动鲜食蚕豆产业持续健康发展的重要途径。蚕豆主要数量性状遗传特性研究较多[4-10],不同亲本的配合力差异较大,为了提高鲜食蚕豆的定向选育效果,开展特殊亲本遗传改良效果评价非常必要,明确鲜食蚕豆主要数量性状的遗传效应对于定向选育品种具有理论指导作用。
1 材料与方法
1.1 材 料
以5个菜用蚕豆品种(系)为供试材料,分别是陵西一寸、通蚕鲜7号、TF 42、GF 74、GF 96,各亲本名称与来源见表1。
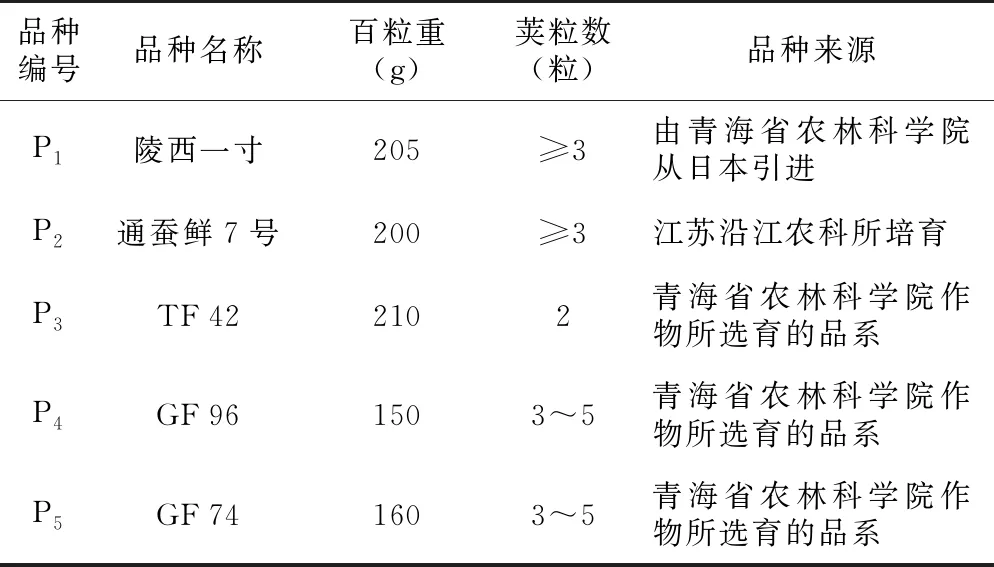
表1 供试亲本来源
1.2 方 法
本试验采用NCII试验设计。以青海本地品系TF 42、GF 74、GF 96为母本,以外来种质通蚕鲜7号、陵西一寸为父本进行杂交,配置6个不完全双列杂交组合。
2018年5月进行杂交组合配置,同年8月收获杂交种。2019年3月,采用随机区组设计种植各组合F1杂交种,一穴一粒点播。同年6月进行表型鉴定,去除假杂种。各组合F1单株成熟后,调查有效分枝数、百粒重、荚层数、单荚粒数、单株荚数、荚长、荚宽等7个主要数量性状。2020年3月,采用随机区组设计种植F2群体,每个组合3次重复,每小区4行,每行20株,共6个小区。F2单株成熟后,调查有效分枝数、百粒重、荚层数、单荚粒数、单株荚数、荚长、荚宽等7个主要数量性状。
1.3 数据处理
利用Microsoft Excel 2016软件对表型性状的杂种优势进行统计分析。利用DPS 19.05数据处理系统进行遗传效应及其相关分析,配合力分析采用NCII设计模型分析。
2 结果与分析
2.1 主要数量性状的改良效果评价
由表2、表3可知,百粒重超亲优势平均值为正值,超越高值亲本。P3×P1、P3×P2超亲优势为10.25%、13.61%,说明这2个组合更有利于培育大粒的菜用蚕豆品种。其余6个性状的超亲优势平均值为负值,但个别组合仍表现出正向超亲优势,可加以利用。P4×P1和P4×P2的有效分枝数超亲优势为2.40%、4.50%,有利于培育多枝品种,从而降低种子使用量;P4×P1和P4×P2的单株荚数的超亲优势为12.36%、15.09%,超亲优势明显,有利于培育多荚的菜用蚕豆品种,杨勇等[11]研究证明,实际产量与单株荚数为正相关关系,培育多荚有利于菜用蚕豆高产。P5×P1和P5×P2荚长的中亲优势为8.81%、9.09%,可培育长荚的菜用蚕豆品种。P3×P2荚宽超亲优势为3.83%,适宜培育宽荚品种。荚层数的超亲优势、中亲优势均为负值,田间观察发现F1杂交后代荚层数均值较双亲有所降低,但单株荚数中亲优势平均值为正值,结荚数超过双亲均值,结荚层数相对集中,不易倒伏。单荚粒数的超亲优势、中亲优势平均值均为负值,P5×P2的中亲优势为1.00%,超越双亲均值,有利于培育多粒品种。P5×P2除荚层数以外的6个性状的中亲优势均为正值,杂种优势较明显。P4×P1和P4×P2除荚层数、单荚粒数2个性状外,其余性状中亲优势均为正值,杂种优势较明显。有效分枝数、荚层数、单株荚数等3个性状变异系数较大,受环境影响较大,变异幅度较大,对后续筛选多枝、多荚层、多荚品种提供了较大的选择空间。综上,获得符合试验预期的菜用蚕豆品种可考虑组合为P5×P2、P4×P2。
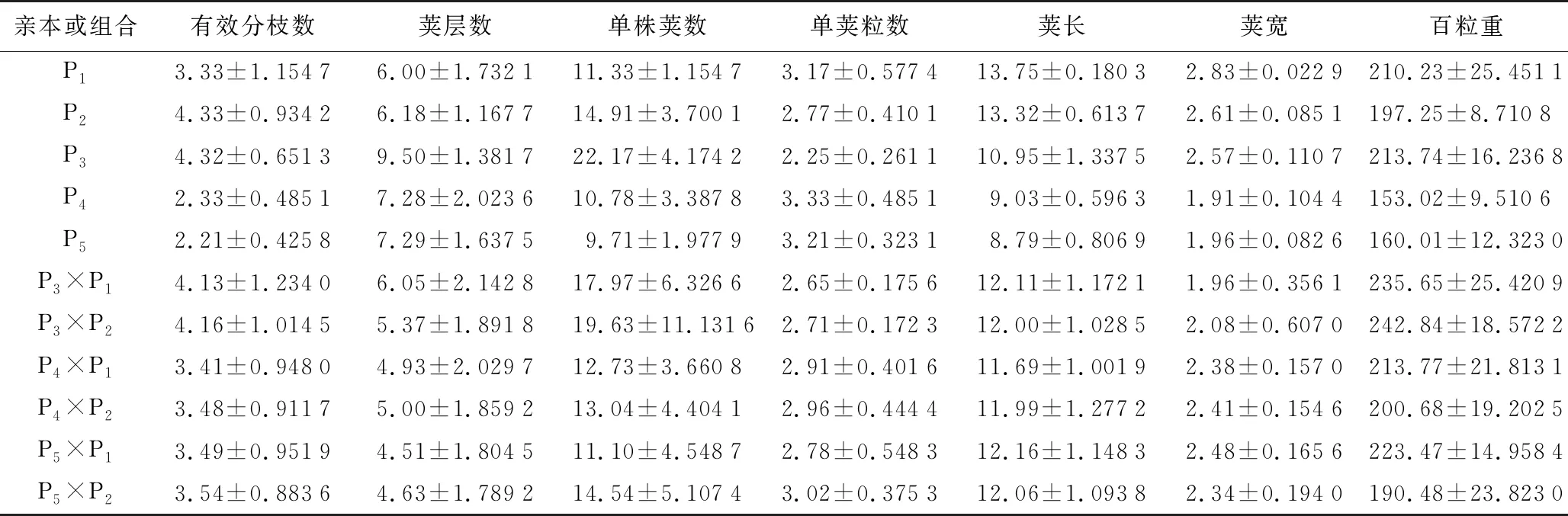
表2 亲本及菜用蚕豆杂交F1代性状表现平均值

表3 菜用蚕豆杂交F1代杂种优势表现
2.2 主要菜用蚕豆品种的配合力分析
2.2.17个数量性状的方差分析
从表4可知,7个性状在亲本或组合间的差异均达到显著或极显著水平,说明进行杂交的两个亲本间的7个数量性状的一般配合力效应和特殊配合力效应对F1杂种后代的影响差异显著,可利用NCII试验模型对菜用蚕豆7个数量性状的一般配合力和特殊配合力效应进行分析。
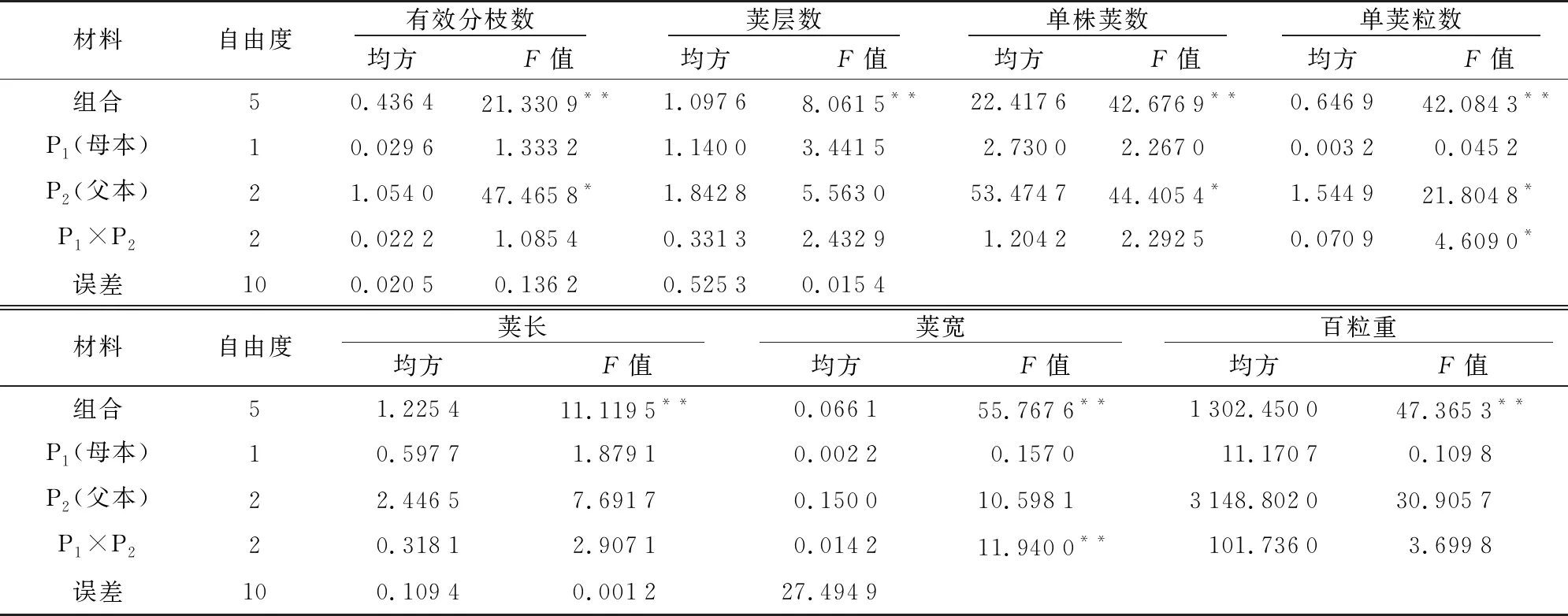
表4 3×2不完全双列杂交菜用蚕豆F1代性状方差分析
2.2.27个数量性状的一般配合力效应分析
由表5可知,同一性状在不同亲本品种之间的一般配合力(General Combining Ability,GCA)效应值的差异较大;而在同一亲本中,不同数量性状的GCA效应值的差异也较大,通常对产量构成因素有影响的性状GCA效应值越大,越易配出符合预期目标的杂交组合。
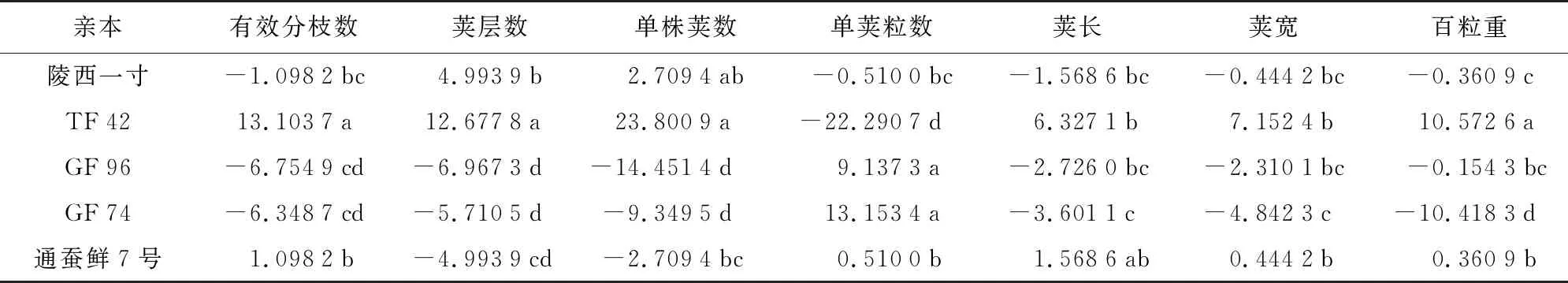
表5 7个数量性状的一般配合力相对效应值
TF 42的有效分枝数、荚层数、单株荚数、荚长、荚宽、百粒重等6个性状的GCA效应值介于6.327 1~23.800 9之间,说明TF 42与其他优良品种杂交后,后代易获得优良表现的性状。TF 42的单荚粒数GCA效应值为负值(-22.290 7),影响其杂交后代的平均单荚粒数。可利用其他单荚粒数较高的品种对TF 42的单荚粒数进行改良时,应注重杂交后代的选择。
GF 96、GF 74、通蚕鲜7号的单荚粒数GCA效应值为正值,GF 74 GCA效应值较高(13.153 4),有利于配出多粒品种。TF 42、通蚕鲜7号的有效分枝数GCA效应值为正值,利用其进行杂交易获得分枝数较多的品种。TF 42、陵西一寸的单株荚数GCA效应值为正值,利用其进行杂交易获得结荚多的品种。TF 42、通蚕鲜7号的荚长、荚宽、百粒重的GCA效应值均为正值,可组配出长荚、宽荚、大粒的品种。预期获得多枝、大粒、多荚、多粒、荚长且宽的菜用蚕豆品种,可选择TF 42、GF 74、通蚕鲜7号、陵西一寸等亲本进行组配。
2.2.37个数量性状的特殊配合力效应分析
由表6可知,同一亲本与不同父本进行杂交,特殊配合力(Specific Combining Ability,SCA)效应值存在差异;而同一数量性状在不同亲本组合中,SCA效应值也具有差异。一般来说,性状的SCA效应值越高,后代表现越好。

表6 7个数量性状的特殊配合力相对效应值
TF 42×陵西一寸的荚层数SCA效应值最高(5.291 6),可用于培育多荚层品种;通蚕鲜7号×TF 42的荚层数SCA效应值为-5.291 6,表现较强的负向效应,F1代中平均荚层数减少,不符合培育多荚层品种的目标。TF 42×陵西一寸、GF 74×通蚕鲜7号的单株荚数的SCA效应值较高,杂交后代中易获得单株荚数较多的单株,符合多荚的育种目标。TF 42×陵西一寸、GF 74×通蚕鲜7号的有效分枝数SCA效应值表现较强的正向效应,符合多枝的育种目标。GF 96×陵西一寸、TF 42×通蚕鲜7号、GF 74×通蚕鲜7号的单荚粒数SCA效应值为正值,表现正向效应,符合多粒的育种目标。TF 42×陵西一寸、GF 74×陵西一寸、GF 96×通蚕鲜7号的百粒重SCA效应值为正值,表现为正向效应,符合大粒的育种目标。本试验预期培育多枝、多荚、大荚、大粒、多粒的菜用型蚕豆,可考虑利用TF 42×陵西一寸、GF 74×通蚕鲜7号等杂交组合,或利用多亲本进行复交。
2.3 7个数量性状亲本配合力基因型方差、群体配合力基因型方差及遗传力分析
由表7可知,在F2杂交组合中,经过基因重组后,VG/VS均大于1,说明7个性状均是加性效应遗传起主导作用,即7个数量性状的表型观测值由双亲多个微效基因的基因型值累加决定。

表7 3×2不完全双列F2杂交组合性状的基因型方差、群体配合力方差及遗传力
广义遗传力由大到小为:单荚粒数、单株荚数、百粒重、荚层数、荚长、荚宽、有效分枝数。狭义遗传力由大到小为:单荚粒数、百粒重、单株荚数、荚层数、荚宽、荚长、有效分枝数。百粒重、单荚粒数、单株荚数的广义遗传力、狭义遗传力皆较高,说明受到环境因素的影响较小,对这3个性状的高值后代可早代选择。荚层数、有效分枝数的广义遗传力、狭义遗传力中等,受到环境因素的影响较大。荚长、荚宽、有效分枝数的广义遗传力、狭义遗传力偏低,应进行多代选择,以提高获得优良后代的可能性。
3 结论与讨论
菜用蚕豆发展前景广阔,而目前菜用蚕豆品种多为适宜浙江、福建、江苏[12-14]等地种植的秋播型品种,适宜青海种植的菜用品种少,研究发展较为缓慢。因此,选育适宜青海种植的优良菜用品种具有较高价值。本试验利用2个外来菜用蚕豆品种(系)与TF 42、GF 74、GF 96进行不完全双列杂交。F1代百粒重超亲优势平均值为正值,超越高值亲本,其余性状超亲优势均为负值,但个别组合性状仍表现出正向超亲优势。单荚粒数、荚层数的超亲优势、中亲优势平均值均为负值,但田间观察发现F1代荚层数下降,结荚位置变得相对集中,3粒及以上荚单株增多。有效分枝数、单株荚数、荚长、荚宽等4个性状的中亲优势为正值,超越双亲均值。有效分枝数、荚层数、单株荚数等三个性状变异系数较大,受到环境影响较大,变异幅度较大,对后续筛选多枝、多荚层、多荚品种提供了较大的筛选空间。玉米、小麦、水稻等禾本科作物上,杂种优势利用已经成熟,利用杂种优势获得了高产。故本试验选择多性状上具有杂种优势的强势组合有利于获得高产品种。综上,预期获得符合试验预期的菜用蚕豆品种可考虑组合为P5×P2、P4×P2。
配合力结果表明,同一亲本不同性状的GCA和SCA表现差异显著,不同亲本同一性状的GCA和SCA表现也有较大的差异,GCA效应值和SCA效应值因亲本及性状不同产生差异,但无明显的规律或对应关系。在大豆中,总配合力与杂种表现的相关性最强[15],菜用蚕豆也可考虑利用TCA效应进行后代选育。TF 42的单荚粒数GCA表现呈负向效应,其余6个数量性状表现为正向效应,可作为杂交组合配置的优良亲本之一,用于选育多枝、大粒、多荚、大荚的菜用品种亲本之一。GF 74单荚粒数的配合力表现相较于其他品种(系)较为突出,有利于培育多粒品种。通蚕鲜7号的GCA表现较为良好,荚层数、单株荚数GCA效应值为负值,其负向效应较弱,可与配合力强的亲本杂交提升结荚数、荚层数。GF 74×通蚕鲜7号的有效分枝数、荚层数、单株荚数、单荚粒数、荚长等5个性状的SCA效应值具有较强的正向效应,荚宽、百粒重表现出负向效应,负向效应较弱,百粒重、荚宽相较低值亲本,仍获得改良,F1代仍表现出较好的杂种优势。TF 42×陵西一寸荚宽、单荚粒数SCA效应值具有负向效应,负向效应轻微,后期筛选宽荚后代进行培育。TF 42×陵西一寸组合的单荚粒数负向效应减弱,3粒荚比例相较于TF 42略有提升,后期筛选多粒荚(3粒及以上)单荚培育多粒后代。综上,TF 42×陵西一寸、 GF 74×通蚕鲜7号的配合力表现较好,后续育种过程中应着重关注单荚粒数、荚宽等数量性状的后代筛选。
百粒重、单株荚数、单荚粒数具有较强的广义遗传力和狭义遗传力,与刘玉皎[16-17]、赵娜等[18]的研究结果相近,对上述3个性状可早代筛选高值后代。刘定富等[19]认为,荚层数(节数)符合加性显性模型,不存在任何方式的母性遗传,荚层数仅受加性效应控制,显性效应不显著,则本试验后续可选择遗传多样性指数高的植株,从F2代中筛选荚层多且具有多个微效基因的单株。易卫平等[20]得出秋播区的15个蚕豆品种的荚宽广义遗传力为73.91%,与本试验广义遗传力值相近,F2代较F1代遗传力水平有所上升,后续试验需多代筛选高值后代。本试验有效分枝数广义遗传力为70.71%,相较其余6个数量性状遗传力最低。郭建华[21]对蚕豆F2代的遗传力研究发现,有效分枝数的广义遗传力较低(均值44.56%),需进行多代筛选多枝后代。崔世友和缪亚梅[22]研究发现,产量构成因素对产量具有较高正向效应,因此改良单株荚数、单荚粒数、荚宽、百粒重等数量性状获得高产、商品性强的菜用蚕豆品种。欧阳裕元等[23]研究认为,荚长对百粒重有较大影响,培育大粒品种需关注荚长,但荚长对产量的影响不大。张宝英和白苇[24]研究发现,有效分枝数、单株荚数、单荚粒数、百粒重对产量影响较大,数量性状相互关联、影响,在满足菜用蚕豆高商品性的同时,想要获得高产应综合考虑各数量性状的影响。本试验对有效分枝数、荚层数、单株荚数、单荚粒数、荚长、荚宽、百粒重等7个数量性状的遗传效应分析后,选择性状具有高正向配合力、杂种优势明显、变异幅度较大的亲本组合为TF 42×陵西一寸、 GF 74×通蚕鲜7号。
