蚯蚓原位堆肥提升番茄连作土壤质量研究
2022-03-16曹云娥吴泽帅张美君李建设田永强
曹云娥,尹 翠,吴泽帅,张美君,李建设*,田永强,3*
(1 宁夏大学农学院,银川 750021;2 宁夏共享人力集团有限公司,银川 750021;3 中国农业大学园艺学院,北京 100193)
连作障碍是长期困扰国内外农业生产的复杂问题之一,寻求克服或缓解连作障碍措施已迫在眉睫。目前生产上解决连作障碍的对策主要包括轮作换茬、间作套种、土壤消毒、抗性品种栽培和生物防治等方法[1]。近年来,大量研究表明,添加土壤改良剂可以缓解连作障碍的发生,其中以有机物料和蚯蚓粪的效果较为明显[2]。由于在改善土壤物理、化学和生物学性质等方面的优势以及较强的实际可操作性,蚯蚓粪目前已被成功应用于作物连作障碍防治。有研究表明,在连作土壤上添加蚓粪,可显著增加土壤中细菌和放线菌数量,降低真菌丰度尤其是尖孢镰刀菌的种群数量,提高土壤酶活性和养分含量,促进植株根系健康生长,提高作物的生物量,进而缓解作物连作障碍[3–4]。
番茄 (Lycopersicon esculentum Mill.) 是设施栽培中种植最广泛的蔬菜作物之一[5]。因其口感适宜,营养价值高,备受消费者喜爱,市场需求量大,经济效益较高,番茄总产量在我国设施农业中位居首位[6]。为追求高产量和高效益,加之规模化和集约化种植,菜田复种指数越来越高,导致菜田土壤微生物种群结构失衡、有害微生物种群密度上升、质量下降、理化性状恶化、养分缺失、农药残留、病虫害加重等一系列连作障碍问题[7–8]。这不仅造成蔬菜大幅减产,而且使得产品品质下降,不利于蔬菜产业的可持续发展[9–10]。连作障碍已成为制约我国番茄可持续生产的主要瓶颈问题之一[11]。
蚯蚓堆肥(vermicompost)是近几年兴起的普适性较好的土壤改良剂,其在克服连作障碍方面的效果显著[12]。蚯蚓堆肥的施用可以减少养分流失,增加土壤肥力,提高碳封存力,保持土壤碳平衡,调节土壤pH等[12–13]。此外,蚯蚓堆肥可通过改变土壤的pH和微量营养元素的可利用性来改善微生物的活性和结构,促进土壤有益微生物的增加[14–15]。例如,蚯蚓堆肥的施用可以明显促进微生物介导的有机质循环,提升土壤细菌群落多样性和微生物碳源利用水平,进而改善土壤整体质量。此外,蚯蚓堆肥的施用可以提高植株的抗性。如有研究表明,施加蚯蚓堆肥会显著提升土壤质量(维持酸碱平衡、增加微生物活性),并增加植株根系累积的防御物质,从而使感病植株表现出较强的根结线虫抗性[16]。蚯蚓堆肥的上述优良性状使得其在蔬菜栽培上的应用前景广阔。
目前, 已有的研究多集中在利用蚯蚓异位处理有机废弃物料形成的蚯蚓粪(蚯蚓异位堆肥)对土壤质量的提升和连作障碍的克服效果,而对利用蚯蚓原位堆肥(在田间栽培畦原位同时投入有机废弃物料和蚯蚓)克服土壤连作障碍的研究报道较少。为此,本研究从土壤微生态角度出发,探讨蚯蚓原位堆肥对连作5年设施番茄土壤细菌群落结构、土壤理化性质的影响,为蚯蚓生物治理番茄连作障碍提供理论支撑,并为设施蔬菜产业的可持续发展提供支持。重点研究细菌群落是因为细菌在设施番茄土壤中占主导作用,是调控土壤质量体系的关键[17–18]。
1 材料与方法
1.1 试验设计
试验于2017年8月6日至2019年1月10日在宁夏银川贺兰山园艺科技示范区 (106.3°E, 38.6°N)光伏温室内进行。供试土壤为连作5年(2012—2016)番茄的沙壤土,2017年土壤的理化性质:全氮0.15 g/kg、全磷 0.12 g/kg、速效氮 13.8 mg/kg、速效磷11.2 mg/kg、速效钾 524 mg/kg、pH 7.76、电导率(EC) =0.89 mS/cm);供试番茄品种为‘粉宴 1号’,起垄种植,株距为30.0 cm,每垄布置两条滴灌管,滴头间距 0.30 m。
试验采用单因素随机区组设计,共设置3个处理:施用化肥对照(CK);蚯蚓异位堆肥(T1;将蚯蚓消解牛粪制备蚯蚓粪均匀撒施在栽培垄上);蚯蚓原位堆肥(T2;将牛粪均匀撒施在栽培垄上,放入蚯蚓消解牛粪)。每个处理设置3个重复,随机排列。小区面积为31.5 m2,每个小区种植3垄,每垄长7.50 m,宽 0.70 m,垄间距 0.70 m。3个处理的氮素投入量相同,两个蚯蚓粪处理包含同样的牛粪和蚯蚓量。CK处理基施尿素0.69 t/hm2;T1处理基施蚯蚓粪 130.4 t/hm2,T2处理直接施入牛粪 179.4 t/hm2、蚯蚓 1.67 t/hm2。蚯蚓粪全氮含量 2.38 g/kg,牛粪全氮含量为1.73 g/kg。供试蚯蚓为赤子爱胜蚓(Eisenia foetida)。在苗期、开花期和坐果期分别随水施入配方水溶肥(尿素183.00 mg/L,磷酸二氢铵77.05 mg/L,硝酸钾 202.00 mg/L,硫酸钾 174.00 mg/L)。每茬拉秧结束后,重复施用上述尿素、蚯蚓粪、牛粪,蚯蚓不再增加(蚯蚓可自身繁殖)。
1.2 样品采集
在每茬番茄盛果期采集根际土壤。每个处理随机选择5株,将番茄连根拔起后,把根际土抖落到报纸上,去除石子、根系等杂物,混匀后过筛。将每个处理样品分成两份,一份装入离心管中,迅速放入液氮中保存,用于土壤微生物基因组 DNA 提取;另一份自然风干,用于土壤理化性质和酶活性测定。
1.3 试验方法
1.3.1 DNA 提取、PCR 扩增和 Illumina Miseq 测序
根据 Power Soil DNA 试剂盒 (MoBio Laboratories Inc.,CA, USA)说明书对土壤总 DNA 进行提取,DNA浓度和纯度利用Nano Drop 2000进行检测,利用1%琼脂糖凝胶电泳检测DNA质量;用338F(5'–ACTCCTACGGGAGGCAGCAG–3')和 806R(5'–GGACTACHVGGGTWTCTAAT–3')引物对细菌DNA的 16 SrRNA V3–V4区段进行 PCR扩增,然后在Illumina MiSeq平台上进行PCR反应,PCR产物纯化并测序。在97%的序列相似度水平上,将质量过滤后的序列进行分类单元(operational taxonomic units, OTUs)的分类。用QIIME进行稀疏分析[19],以区分OTU丰富度、Shannon丰富度和总物种丰富度的Chao1水平[20]、Coverage指数和Simpson指数[21]。为了预测微生物群落的代谢功能,使用PICRUSt从16S rRNA 数据生成功能图谱[22]。
1.3.2 土壤理化性质测定土壤理化性质测定方法参照《土壤农化分析》[23]进行。在土水质量比为1∶10时测定样品pH和EC值;用凯氏定氮法测定全氮含量;用碱解扩散法测定碱解氮含量;用重铬酸钾-硫酸氧化法测定土壤有机质含量;用NaHCO3(0.5 mol/L)浸提—钼锑抗比色法测定速效磷含量;采用高氯酸–硫酸法测定土壤全磷;采用氢氟酸–高氯酸法测定土壤全钾;用 NH4OAc (1 mol/L) 浸提—火焰光度法测定速效钾含量。
1.4 数据处理与分析
采用Excel 2010软件进行数据整理和相关图表制作,采用SPSS 20.0软件进行显著性(P<0.05)和Pearson相关性分析。采用单因素分析(one-way ANOVA),当数据间达到显著性差异(P<0.05)时,采用Tukey’s HSD 进行方差分析。利用北京奥维森基因科技有限公司的测序平台(https://cloud.allwegene.com) 进行 α 多样性分析。使用 CANOCO 5.0软件分析土壤细菌门水平群落结构、细菌相对丰度。对于高通量原始测序数据,去除接头序列后,根据overlap将双端测序序列拼接成单条序列,并对序列质量进行质控和过滤。参考已知数据库GreenGenes去除嵌合体序列,得到最终可用的序列,并将质量过滤后的序列在97%的序列相似度水平上进行分类单元(OTUs)的分类。由于微生物的群落功能组成与环境的关系比物种组成更为密切,因此基于16 SrRNA的测序结果,采用PICRUSt进行了功能预测。为了准确预测功能构成,首先对原始测序数据的种属数量进行标准化处理,然后将其映射在构建好的已测序基因组的种属功能基因构成表上,获得生化代谢途径和三级KEGG预测的功能结果。
2 结果与分析
2.1 不同蚯蚓堆肥处理对连作土壤理化性质的影响
由图1可知,连续3茬蚯蚓异位堆肥(T1)和原位堆肥(T2)处理下土壤EC和pH均显著降低,且前两茬T2处理EC显著低于T1,第三茬T2和T1处理间无显著差异;土壤全氮、全磷、全钾、总碳、有效氮和速效钾含量三茬基本表现为T2>T1>CK(P<0.05)的趋势。这说明,相较于蚯蚓异位堆肥,蚯蚓原位堆肥更有利于提升土壤肥力水平。而且,由于原位堆肥无需每次加入蚯蚓,其堆肥的制作(无需占地和二次运输)和使用成本也大大降低。
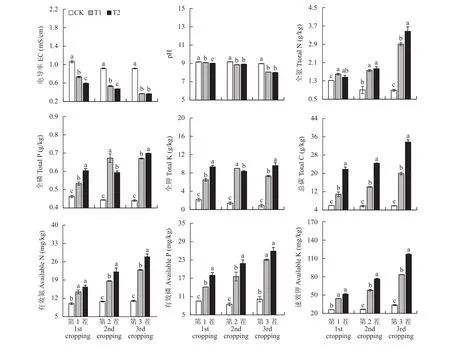
图1 不同处理下土壤理化性质Fig. 1 Physiochemical properties of soils under different treatments
2.2 不同蚯蚓堆肥处理对连作土壤细菌群落数量、多样性及丰富度的影响
由表1可知,3个茬口各处理的Coverage指数均在0.9233~0.9567,表明微生物数量分析的可靠性均很高。从多样性指数看,CK处理的OTUs数量显著低于T2和T1处理,平均下降了17.41%。OTUs数量、Chao1指数和Shannon指数T1处理在第一茬与CK没有显著差异,从第二茬起显著高于CK,而T2处理从第一茬起即显著高于CK,且在第三茬显著高于T1 (P<0.05)。Simpson指数反映的是物种的优势度,3个处理的Simpson指数无显著性差异。说明蚯蚓堆肥处理主要显著增加了土壤细菌群落的多样性,但没有改变细菌的构成,且原位堆肥提高细菌多样性的效果显著好于异位堆肥。
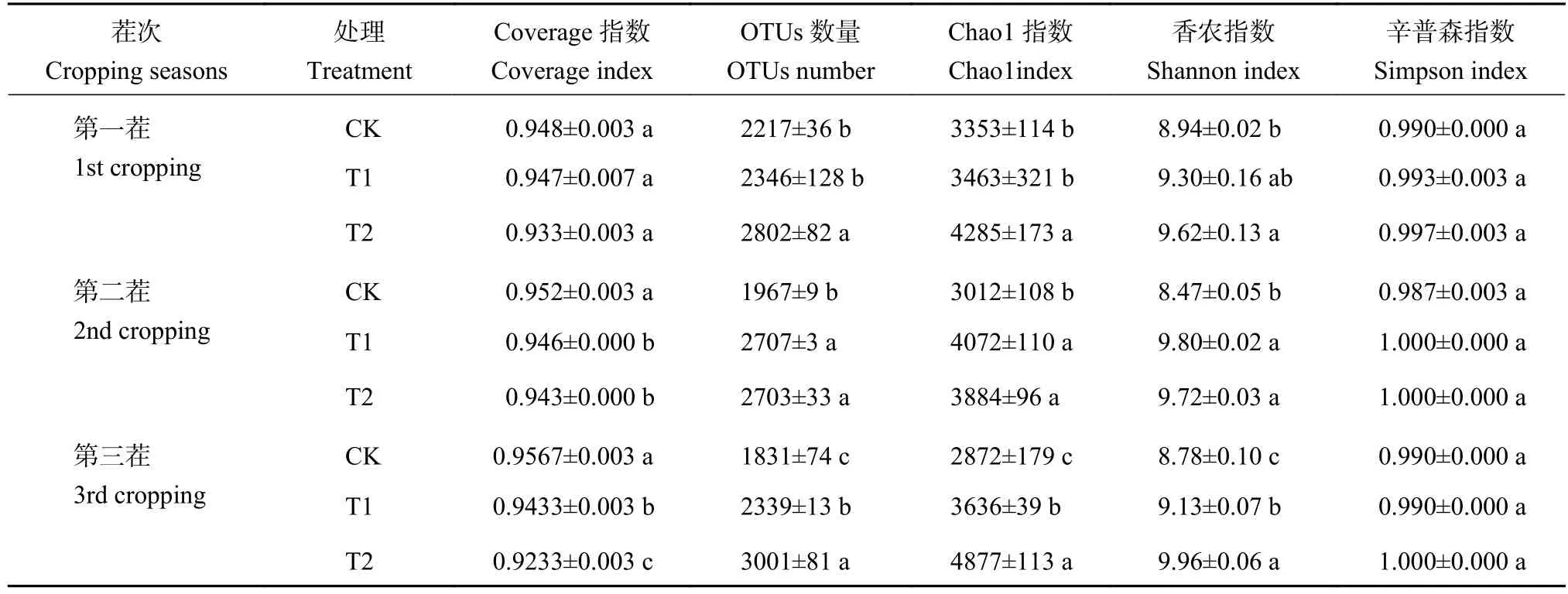
表1 土壤细菌群落多样性和丰富度指数Table 1 Soil bacterial community diversity and richness index
2.3 不同蚯蚓堆肥处理对连作土壤细菌群落组成的影响
在门水平上,共获得48个相对丰度>0.5%的菌群(图2)。其中,平均相对丰度>2%的菌群在3个处理土壤中的相对丰度为:变形菌门 (Proteobacteria)29.00%~59.06%、放线菌门 (Actinobacteria) 13.63%~34.65%、厚壁菌门(Firmicutes) 3.92%~13.80%、绿弯菌门 (Chloroflexi) 5.64%~13.34%、酸杆菌门(Acidobacteria)2.00%~10.00%、拟杆菌门(Bacteroidetes) 1.79%~16.02%、芽单胞菌门(Gemmatimonadetes) 2.00%~4.32%、硝化螺旋菌门(Nitrospirae) 0.52%~3.21%,占到全部菌门的 92.71%~97.44%。平均相对丰度均< 2%的其他7类菌群仅占整个菌门的2.56%~7.29%。蚯蚓处理土壤相对丰度较高的菌群随种植茬数的延长有变化,在第1茬相对丰度最高的优势菌门是变形菌门,第2茬是厚壁菌门和芽单胞菌门,第3茬则是变形菌门绿弯菌门和异常球菌–栖热菌门(Deinococcus-Thermus)。
连续3茬施肥处理后,相较于CK,T1和T2处理的变形菌门、放线菌门、厚壁菌门、绿弯菌门、芽单胞菌门和硝化螺旋菌门均显著增加,而与连作障碍相关的酸杆菌门和拟杆菌门显著降低。这说明,蚯蚓异位堆肥(T1)和蚯蚓原位堆肥(T2)处理均能够抑制这些菌在土壤中的相对丰度。此外,相较于CK和T1,T2处理显著增加了绿弯菌门、芽单胞菌门的相对丰度,说明这两类菌可能是蚯蚓原位堆肥(T2)处理抑制连作障碍相关菌的特殊菌门。
在属水平上,相对丰度大于0.5%的菌属如图2所示。变形菌门中的假单胞菌属(Pseudomonas)、鞘氨醇单胞菌属(Sphingomonas)、亚硝酸菌属(Nitrosomonas),厚壁菌门的芽孢杆菌属(Bacillus),放线菌门的链霉菌属(Streptomyces),污物假节杆菌属(Pseudarthrobacter)和硝化螺旋菌门的硝化螺旋菌属(Nitrospira)平均相对丰度均大于1%,为优势菌属。与CK相比,T1和T2处理的假单胞菌属、链霉菌属、硝化螺旋菌属和污物假节杆菌属的相对丰度无显著差异,而鞘氨醇单胞菌属、亚硝酸菌属和芽孢杆菌属的相对丰度显著增加。这说明,鞘氨醇单胞菌属、亚硝酸菌属和芽孢杆菌属可能是蚯蚓异位堆肥(T1)和蚯蚓原位堆肥(T2)处理抑制连作障碍的优势菌属。
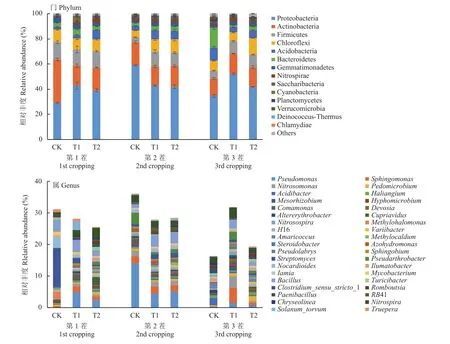
图2 不同处理土壤优势细菌门和属的相对丰度Fig. 2 Relative abundance of dominant soil bacteria at the phylum and genus levels under different treatments
2.4 不同蚯蚓堆肥处理对连作土壤代谢功能的影响
微生物群落的变化直接或间接地改变了代谢通路。为更直观了解此种趋势,本部分基于KEGG代谢通路数据库的代谢功能各个处理间存在显著性差异。由图3可知,关于代谢途径序列的相对丰度解释了81.16%。在相对丰度大于5%的代谢通路中,第2茬蚯蚓改良处理下碳水化合物代谢、辅酶与维生素代谢、萜类和聚酮类化合物的代谢和其他氨基酸的代谢途径的相对丰度显著高于CK处理。第3茬蚯蚓改良处理下碳水化合物代谢、氨基酸代谢的相对丰度显著高于CK处理。
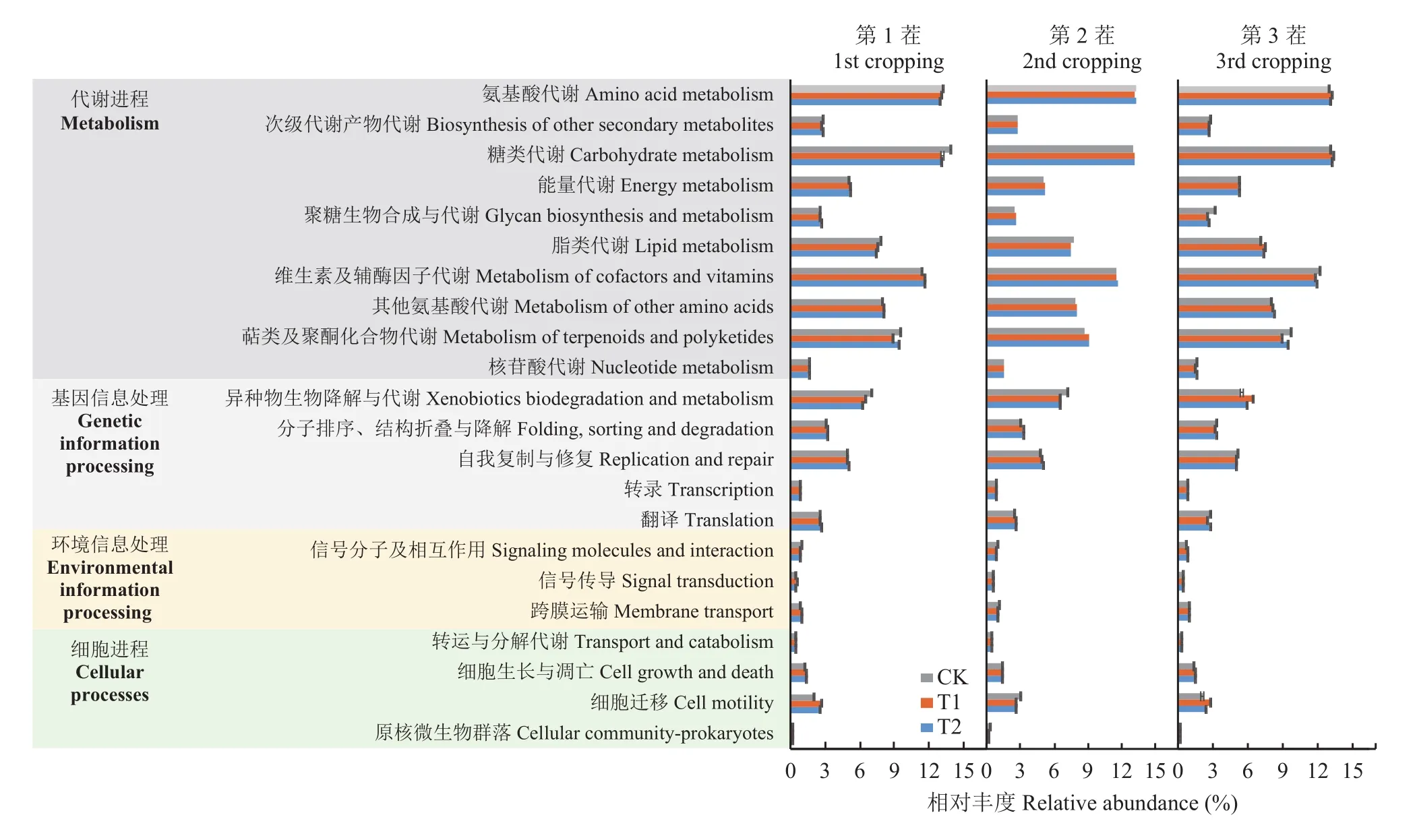
图3 不同处理土壤细菌群落生化代谢途径Fig. 3 Biochemical metabolic pathways of soil bacterial community as affected by treatments
图4列出了相对丰度排名前20的3级KEGG同源功能预测。3级KEGG同源功能预测结果显示,在排名前20的代谢途径中,3茬蚯蚓改良处理下D-精氨酸与 D-鸟氨酸代谢、C5-Branched dibasic acid metabolism 和 Cell cycle - Caulobacter的相对丰度显著高于CK处理,蚯蚓处理(T2)的C5-Branched dibasic acid metabolism、Sulfur relay system 和 Cell cycle-Caulobacter显著高于其他处理。说明C5-Branched dibasic acid metabolism 代谢途径在改良连作土壤过程中具有重要作用。C5-Branched dibasic acid metabolism属于二级代谢途径中碳水化合物代谢途径。再次验证了碳水化合物代谢途径在蚯蚓(粪)改良连作土壤中有重要作用。

图4 不同处理土壤细菌群落3级KEGG同源功能的预测Fig. 4 The level 3 KEGG homologous function predictions of soil bacterial community as affected by treatments
2.5 土壤环境因子对门分类水平的细菌群落结构或代谢功能的影响
采用RDA分析了土壤环境因子对门分类水平的细菌群落结构或代谢功能的影响(图5a)。由图5a可知,第一排序轴和第二排序轴分别解释了细菌菌落变化的59.94%和22.78% (合计82.72%)。经过蒙特卡罗检验,TC (F=5.0,P=0.014)、EC (F=4.1,P=0.028)、TK (F=4.7,P=0.012)、AN (F=4.5,P=0.024)、TP (F=6.6,P=0.008)是解释菌落差异的关键性因素。再次证实了蚯蚓能够加速有机物质的降解,提高土壤的总碳含量,进而影响细菌群落结构。由相关性热图分析可知(图5b),EC、AN、TN、TP、AP、AK、TC和TN与门分类水平的细菌群落结构有显著或极显著正负相关关系。EC与Actinobacteria呈显著正相关关系;AN和Bacteroidetes、Verrucomicrobia、Chlamydiae呈显著负相关关系,与Peregrinibacteria呈极显著负相关关系;TP与Armatimonadetes、WS6、Gracilibacteria和 Hydrogenedentes呈显著性正相关关系;TC与Chloroflex、Tectomicrobia、Armatimonadetes和 Gracilibacteria呈显著正相关关系;Chloroflexi与AK、TC和TN呈显著负相关关系。在平均相对丰度大于1%的菌门中,EC、AN、TN、TP、AP、AK、TC和 TN与Actinobacteria和Chloroflexi呈显著正相关关系,说明土壤特性会显著影响Actinobacteria和Chloroflexi菌门。这与前面蚯蚓处理对连作土壤细菌群落组成的影响结果一致,Actinobacteria、Firmicutes、Gemmatimonadetes、Deinococcus-Thermus、Chloroflexi和Nitrospirae在蚯蚓改良过程中起到了重要的作用。
为进一步分析不同土壤环境因子对KEGG代谢功能的影响,对土壤环境因子与KEGG代谢功能进行RDA分析。由图5a可知,第一排序轴和第二排序轴分别解释了代谢功能的66.83%和25.94% (合计92.77%)。经过蒙特卡罗检验,EC (F=4.1,P=0.028)、TP (F=6.3,P=0.006)、TK (F=3.6,P=0.034)、AP (F=5.6,P=0.004)、AN (F=4.4,
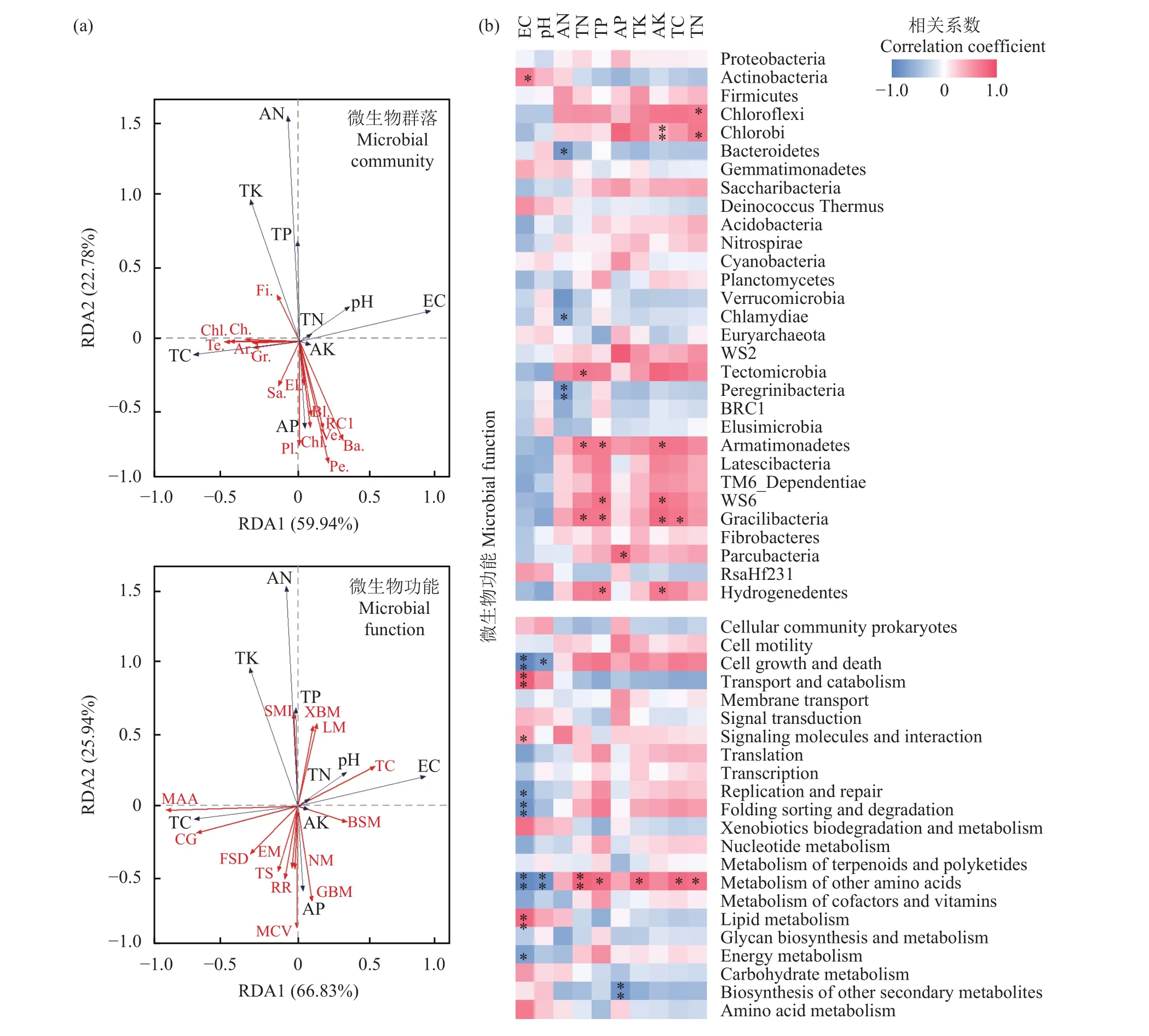
图5 微生物群落、功能与土壤理化性质的RDA冗余分析(a)及相互关系热图(b)Fig. 5 RDA plots (a) and heatmap visualizing relationships (b) between microbial communities, metabolic functions and physiochemical properties
P=0.028)是解释代谢功能的关键性因素。由相关性热图(图4b)分析可知,EC、pH、TN、TP、AP、TK、TC和TN与代谢功能有显著或极显著正负相关关系。EC与细胞生长与死亡、大分子化合物折叠、降解和氨基酸代谢呈极显著负相关关系,与遗传物质翻译、复制与修复呈显著负相关关系;与物质转运与分解代谢和类脂化合物代谢呈极显著正相关关系,与氨基酸代谢呈显著正相关关系。氨基酸代谢与EC和pH呈极显著负相关关系;与TN呈极显著正相关关系;与TP、TK和TC呈显著正相关关系。说明EC是影响KEGG代谢功能的关键土壤因子。
2.6 土壤细菌群落多样性与土壤肥力指标的Person相关性分析
由土壤细菌群落多样性与土壤肥力指标的Person相关性分析(表2)表明,除有效磷与Simpson指数没有相关性外,TN、AN、TP、TK、AK和TC均有显著(P<0.05)或极显著(P<0.01)正相关关系(表2),表明蚯蚓活动和蚯蚓粪可能通过增加土壤微生物多样性进而提升土壤养分含量。因为在土壤养分循环过程中,土壤细菌占有十分重要的地位,因此当土壤细菌的数量降低时会引起土壤微生物功能的失调,从而降低土壤养分。
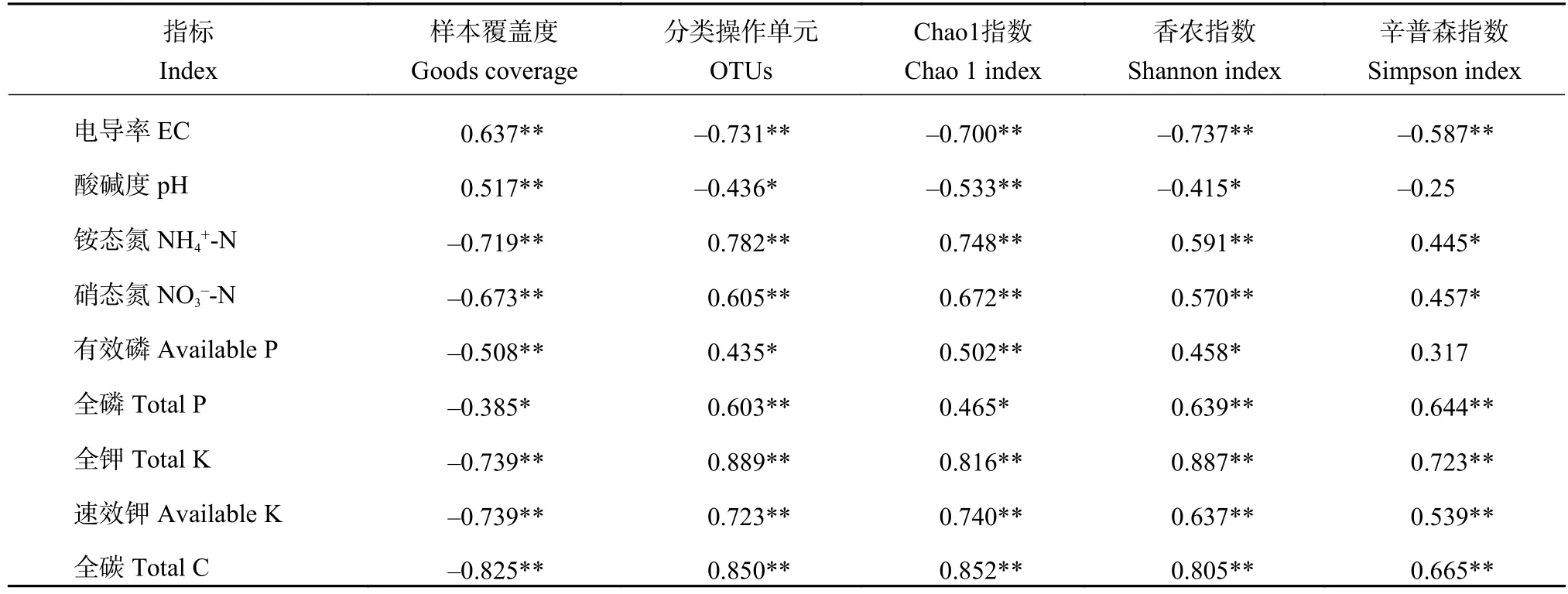
表2 土壤细菌群落多样性与土壤理化性质的相关性Table 2 Correlation between microbial diversity indices and fertility of soils
3 讨论
3.1 蚯蚓堆肥可提高连作土壤的微生物多样性,抑制病原菌的数量
随着集约化生产的进一步发展,复种指数高和茬口单一等原因,设施大棚土壤连作障碍问题日益突出。连作常会导致土壤板结和次生盐渍化、作物生理病害加重、植株生长发育异常、果实品质变劣和产量下降等[24]。连作障碍已成为制约我国一些地区种植业可持续发展的瓶颈问题。蚯蚓作为"生态系统工程师",通过取食、消化、排泄(蚯蚓粪)、分泌(粘液)和掘穴等活动方式(蚯蚓处理)促进植物残枝落叶的降解及有机物质的分解和矿化,进而改善土壤结构,增加土壤养分的可利用性,调控土壤微生物群落,提高土壤中有益微生物的种群数量和土壤肥力[25–26]。蚯蚓粪处理废弃物形成蚯蚓粪,是一种将植物营养、生物活性激素和植物保护融为一体的优质高效有机肥料和土壤改良剂,蚯蚓粪内含有大量的有益微生物,具有促进根系生长的作用[27]。连作土壤有其不同于其他耕作地的特点,例如连作草莓会发生尖孢镰刀菌引起的枯萎病[32]。蚯蚓粪改良连作土壤的研究报道已有很多,目前已成功应用于连作障碍防治,并取得了一定的研究成果。比如施用蚯蚓粪提高韭菜基质中细菌和真菌的多样性,增加养分有效性[28],添加蚓粪或灭菌蚓粪显著增加了连作土壤中的细菌和放线菌数量,降低了真菌特别是尖孢镰刀菌数量,进而减轻苹果连作障碍带来的危害[4]。蚓粪处理番茄连作土壤显著降低了优势真菌的相对丰度,从而缓解了番茄连作障碍[3]。施用蚯蚓粪较牛粪能减轻连作障碍中的酸化感效应,促进脲酶等土壤相关酶活性提高,是缓解连作障碍及连作土壤灭菌对草莓植株生长发育影响的有效措施[2]。
在草莓连作土壤灭菌后施用具有生物活性的蚯蚓粪,可以促进根系生长,缓解土壤灭菌对草莓植株生长发育的影响,是值得推荐的有效措施[29]。通过温室盆栽试验,研究了在不同番茄种植历史的土壤中,全生育期用有机肥代替矿质肥对土壤肥力、番茄产量和品质的影响。结果显示,蚯蚓粪可作为改善连作番茄果实质量、产量和土壤质量的肥料[30]。利用温室盆栽试验研究0、5、和20年种植番茄,蚯蚓粪对土壤微生物特性、酶活性和番茄产量的影响,结果显示蚯蚓能改善土壤微生物的功能,可以代替农家粪肥改善温室番茄连作质量[31]。本试验结果表明,蚯蚓原位堆肥(T2)处理的土壤细菌OTUs数量、Chao1指数和Shannon指数整体上高于蚯蚓粪异位堆肥(T1)和CK (表1),且其通过增加绿弯菌门、芽单胞菌门的相对丰度抑制了连作障碍相关的酸杆菌门和拟杆菌门(图2)。这说明与蚯蚓异位堆肥相比,蚯蚓原位堆肥更能够改善细菌群落组成、增加多样性,且大大降低了前期农业废弃物原位还田后有害菌群增殖的风险,从而抑制了连作障碍的发生。
3.2 蚯蚓堆肥可改善连作土壤的肥力和理化性状
在本试验中,施用蚯蚓化解的牛粪显著降低了碱性土壤的EC和pH, 提高了土壤中有机碳、全量氮磷钾和速效氮磷钾含量,在番茄种植垄上施用牛粪,将蚯蚓直接接种于牛粪的效果比使用蚯蚓腐解牛粪的效果更好(图1)。Wang等[30]、Zhao等[34]的研究也得出类似的结果。蚯蚓原位堆肥处理能够改善连作土壤的肥力和理化性状,其原因可归结为有机碳投入和蚯蚓活动两方面:一方面,牛粪富含有机碳,其在土壤中的投入量多,不仅能直接提高土壤肥力,而且能改善土壤物理性状(如降低容重、增加孔隙度、提高持水持肥力[30,34];另一方面,活蚯蚓的吞食与挖掘活动,不仅可以疏松土壤并提高通气性,而且能够使土壤有机质呈斑块状分布,并使有机质与矿质土混合形成富含有机质的土壤微粒[35]。连作土壤施用适量蚯蚓粪可调节土壤酸碱度,降低总盐分和电导率,一定程度上缓解土壤酸化、盐渍化及养分比例失衡等问题。本试验中,蚯蚓改良处理,均显著提高了连作土壤的总氮、总磷、总钾、速效氮、速效磷和速效钾含量,这与商丽荣等[36]的研究结果一致。值得注意的是,蚯蚓原位堆肥处理在全部3茬的总有机碳(TOC)、速效氮(AN)、速效磷(AP)与速效钾(AK)含量均显著高于蚯蚓异位堆肥处理。此外,由相关RDA分析进一步表明,TOC、AN、AP与AK与放线菌门呈现显著正相关关系。虽然放线菌门与变形菌门同样有着较宽的生态幅[37],但由于放线菌属于富营养型革兰氏阳性菌[38],能够分解蚯蚓堆肥中的残存纤维素、木质素,随着有机质含量的增加,蚯蚓原位堆肥的放线菌门会在第二、三茬多于蚯蚓异位堆肥发酵(图2)。此外,蚯蚓原位堆肥形成的蚯蚓粪中有益微生物丰度高于蚯蚓异位堆肥,其能够促进有机质的分解和养分的矿化,是植物营养元素的活性库[39]。
3.3 不同蚯蚓堆肥处理对连作土壤细菌群落组成的影响
有研究报道,赤子爱胜蚓肠道内的细菌类群主要为放线菌门、拟杆菌门、绿弯菌门 、厚壁菌门、浮霉菌门[40]。当蚯蚓摄取土壤后,土壤在蚯蚓肠道内的运输过程是使土壤细菌群落结构发生改变的关键[41]。本研究发现,在门分类水平上,变形菌门、放线菌门 、厚壁菌门、硝化螺旋菌门等在连作土壤改良过程中起潜在的作用。微生物的生长活性通常受到土壤理化性质的限制,在土壤理化性质不同的区域微生物群落组成也不同[42],本研究中不同蚯蚓堆肥处理均显著提升了土壤养分含量,之前报道属于富营养型细菌的变形菌门的绝大多数、拟杆菌门[43]在本研究中也都成为了优势种,这一点和前人的研究较为类似,即细菌在空间上分布主要由养分所驱动[44]。在属水平上,芽孢杆菌属丰度较高,与这个属大多是需氧或兼性厌氧型的光合细菌有关[45],而蚯蚓由于自身摄食、运动所形成的蚓触圈,能够在原位堆肥过程中形成疏松多孔的蚯蚓粪。这可能是蚯蚓原位堆肥处理的芽孢杆菌属丰度高于蚯蚓异位堆肥的原因。前期研究发现,变形菌门、放线菌门和厚壁菌门在有机质矿化和营养循环中发挥着重要作用[42]。尽管部分研究认为土壤细菌在门水平的大体组成非常类似,均以变形菌门、酸杆菌门和放线菌门相对丰度为最高,总相对丰度达到56.6%~73.8%[43],但是深入调查发现土壤和蚯蚓肠道样品之间大约有33.3%的共享OTUs,其中放线菌门(占总序列的39.2%)、变形菌门(26.9%)和厚壁菌门(18.7%)是蚯蚓肠道内最主要的3类细菌群落,土壤中的优势菌群主要是变形菌门(48.7%)、厚壁菌门(14.3%)、拟杆菌门(10.1%)和放线菌门(9.1%)[44]。本研究结果较有力地支持了上述结论。此外,部分研究所报道的“特殊性抑制”作用中的球孢链霉菌、丁香苷链霉菌等[46]本研究并未检测到,仍需后期的深入研究。
3.4 不同蚯蚓堆肥处理对连作土壤细菌群落和代谢功能的影响
前期研究结果表明,连作土壤施用适量蚯蚓粪可提高果实维生素C和可溶性糖含量,降低硝酸盐含量及总酸度,增大糖酸比,明显改善品质[29, 47–48]。本研究基于KEGG代谢通路数据库的代谢功能分析,在连作土壤改良过程中,碳水化合物代谢途径起到重要作用。同时对相对丰度排名前20的3级KEGG同源功能进行预测,得出C5-Branched dibasic acid metabolism不仅在蚯蚓改良处理,而且在蚯蚓处理中具有重要的作用。C5-Branched dibasic acid metabolism属于二级代谢途径碳水化合物的代谢途径。再次验证了碳水化合物代谢途径在蚯蚓(粪)改良连作土壤中有重要的作用。值得注意的是,本研究发现蚯蚓原位堆肥处理富集到的萜类化合物的代谢路径远多于异位堆肥处理(图3),而萜类物质一直都是重要的化感物质,如Ladhar等[49]确定了两个新的达马烷型三萜化合物抑制醉花蝶属植物种子萌发和幼苗生长,直至最近人们从放线菌中分离得到了一系列新颖的萜类化合物[50]。这可能是蚯蚓原位堆肥较蚯蚓异位堆肥改善土壤细菌群落组成和提升连作土壤质量的原因之一。此外,Šimek和Pill的室内试验研究表明,在蚯蚓的体表和肠道中有固氮菌的出现[51];而在红壤中添加不同量的蚯蚓粪时也发现,NO3–含量随蚯蚓添加量显著升高[52]。而本研究代谢功能分析也检测到了关于氨基酸的代谢路径。该部分结果从侧面印证了蚯蚓堆肥平衡了由于连作造成的土壤养分失调,改善了土壤理化条件,有效促进了土壤微生物生长,进而改善了土壤微生物量碳氮比例[35]。这其中土壤中的蛋白酶和蔗糖酶活性、含量变化则需要更进一步地探究。
4 结论
蚯蚓异位堆肥和原位堆肥均显著降低了土壤EC值,提高了土壤有机质、全量氮磷钾和速效氮磷钾含量,增加连作番茄土壤细菌多样性,改善土壤细菌群落结构和代谢功能。
蚯蚓原位和异地堆肥显著提高了土壤中变形菌门、放线菌门、厚壁菌门、绿弯菌门、芽单胞菌门和硝化螺旋菌门的数量,降低了与连作障碍相关的酸杆菌门和拟杆菌门的数量。相较于化肥和蚯蚓异位堆肥,蚯蚓原位堆肥显著增加了绿弯菌门、芽单胞菌门的相对丰度,因而缓解连作障碍的效果更佳。因此,在实际生产中,推荐蚯蚓原位堆肥以获得更好的缓解连作番茄土壤次生盐渍化与碱化的效果,并降低有机肥的使用成本。
