刚竹属3个竹种根际土壤微生物群落结构
2020-11-11况小宝曾庆南黄思远卢玉生
李 怡 程 平 余 林 况小宝 曾庆南 黄思远 卢玉生
(江西省林业科学院 南昌 330013)
植物根际微生物参与植物与土壤间的物质循环,因此其对于植物生长以及土壤生态环境有着十分重要的意义[1]。根际微生物能够通过分泌激素来调控植物生长代谢,此外它们也可通过多种方式抵御根际有害微生物[2]。植物的根际通过产生分泌物逐渐改变土壤的理化性质,进而影响根际微生物群落结构,提高植物的抗逆性[3-5]。每种植物都能形成其特异性的根际微生物群落,特别是菌根菌、固氮菌等一些与植物联系较为紧密的微生物[6-7]。李菁等[8]研究发现,3种杓兰属植物菌根真菌群落结构差异显著,且指示物种互不相同;叶文雨等[9]研究发现,2种菌草根际土壤细菌群落结构也有较明显的差异。植物物种间的系统发育学距离越大,根际微生物群落组成差异就越显著[10],如农泽梅等[11]研究发现不同甘蔗品种的根际细菌群落主要差异表现在优势菌属丰度上。Hartmann等[12]研究几种豆科植物发现,豌豆与羽扇豆根际微生物群落结构相似,而与蚕豆却有显著差异。
江西省林科院竹种园的竹林成林后施肥量低,土壤微生物群落结构因施肥所产生的影响较小,故本研究选择竹种园内生长情况良好的刚竹属竹种,以毛竹、雷竹以及黄秆乌哺鸡竹这3个竹种的根际土壤作为研究对象,采用Illumina Hi Seq高通量测序方式对3个竹种的根际微生物群落进行分析,通过比较3种不同竹种的根际微生物群落的差异,为竹种专用微生物菌肥的开发与利用提供研究基础。
1 试验地概况
研究试验地位于江西省林业科学院竹种园,地处28°44′41′′N,115°48′46′′E,属亚热带季风湿润型气候,年平均气温19 ℃,年平均降雨量1 713.5 mm。试验地为红壤土,土层厚60 cm以上,pH值约4.1~5.0,有机质含量2.0%~3.5%。成林后每年施复合肥50 kg/667 m2、尿素25 kg/667 m2。林分密度为6 100~9 300株/hm2。
2 研究材料与方法
2.1 研究材料
2019年12月,在江西省林业科学院竹种园采集毛竹(Phyllostachysedulis)、雷竹(Ph.praecox)和黄秆乌哺鸡竹(Ph.vivaxf.aureocaulis)的根际土壤样品,挖取15~20 cm左右竹鞭根,用钥匙将附在根上刮下的土视为根际土壤(剥落分离法)[13],分别标记为毛竹(MR)、雷竹(LR)、黄秆乌哺鸡竹(JR),以附近非竹林土壤作为空白对照(YK)。采样当天将土样带回实验室,过2 mm筛后,置于-80 ℃冰箱保存,供检测使用。
2.2 土壤样品测定
1) DNA提取。取0.5 g土壤样品,用试剂盒提取土壤样品总DNA。
2) 土壤细菌、真菌序列的PCR扩增及测序。以各土壤样品微生物总DNA为模板对细菌16S rRNA V3+V4区域、nifH功能区域及真菌ITS1区域进行PCR扩增。委托上海派森诺生物科技有限公司对PCR产物进行高通量分析测序。
2.3 数据处理
对序列按相似性≥97%水平进行归类操作及OTU划分,筛选OTU的代表序列。对OTU结果应用QIIME软件计算土样的Alpha多样性,再利用R语言工具绘制群落结构图。
3 结果与分析
3.1 3个竹种根际土壤微生物OTU水平
在3个竹种根际土壤总DNA的16S rDNA V3+V4区得到656 203条有效序列,在ITS1区得到1 225 304条有效序列。
基于相似性≥97%的水平,对样品的细菌及真菌有效序列进行聚类,如图1。通过聚类,从3个竹种根际细菌中共获得4 512个OTUs,毛竹、雷竹、黄秆乌哺鸡竹根际土壤细菌OTUs数量分别为2 149、2 449、2 536个,三者特有OTUs数量分别为635、1 118、815个,三者共有OTUs为723个。竹种根际真菌共获得922个OTUs,毛竹、雷竹、黄秆乌哺鸡竹根际土壤真菌OTUs数量分别为409、470、421个,三者特有OTUs数量分别为179、258、193个,三者共有OTUs为86个。可见,根际土壤细菌OTUs数量远高于真菌。
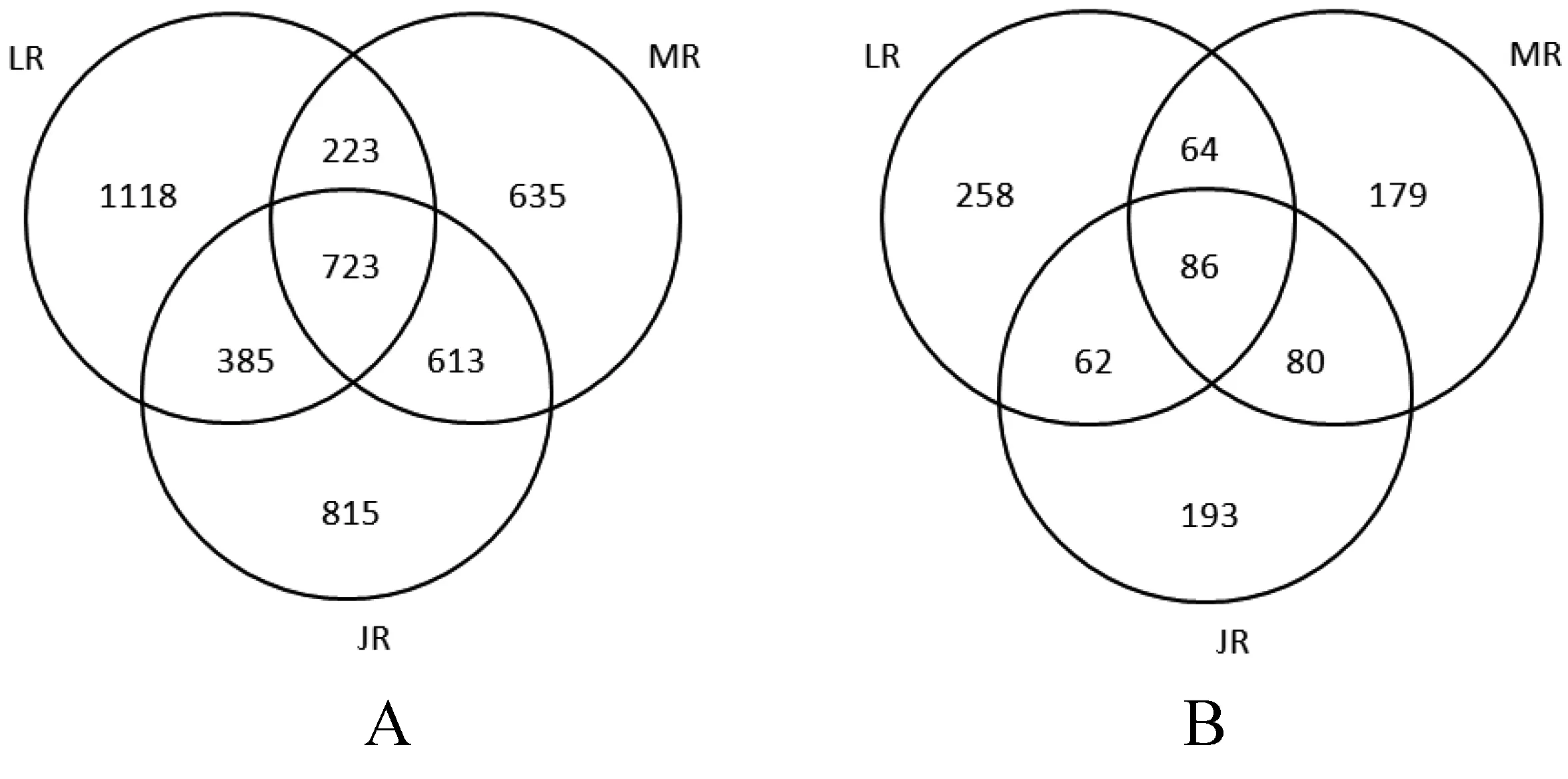
注:A:细菌OUT;B:真菌OUT。图1 竹种根际菌落OTUs分布Venn图
3.2 3个竹种根际土壤微生物Alpha多样性
毛竹、雷竹及黄秆乌哺鸡竹根际土壤微生物群落多样性指数Simpson、Shannon及群落丰富度指数ACE、Chao1见表1—表3。可以看出,3个竹种根际土壤细菌、固氮菌群落的丰富度及多样性均高于非竹林土壤;土壤真菌的丰富度高于非竹林土壤,而多样性低于非竹林土壤。其中黄秆哺鸡竹根际土壤细菌丰富度最高,雷竹根际土壤细菌多样性最低;雷竹根际土壤真菌丰富度及多样性均最高,毛竹根际土壤真菌丰富度及多样性均最低;雷竹根际土壤固氮菌丰富度及多样性均最高,毛竹根际土壤固氮菌丰富度最低,黄秆哺鸡竹根际土壤固氮菌多样性最低。
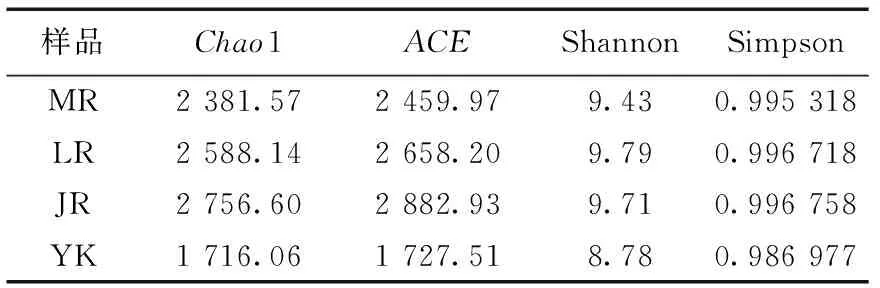
表1 不同竹种根际土壤细菌多样性指数
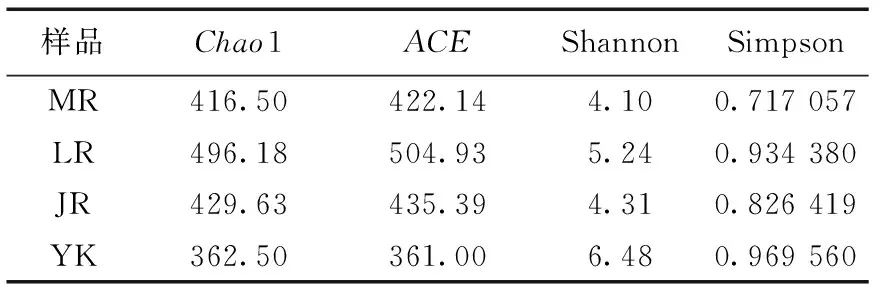
表2 不同竹种根际土壤真菌多样性指数
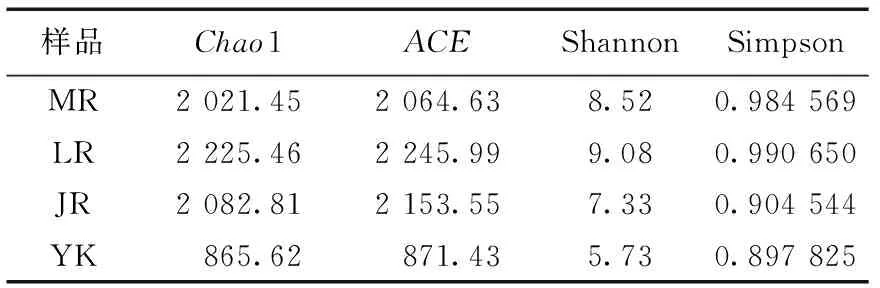
表3 不同竹种根际土壤固氮菌多样性指数
OTU水平及Alpha多样性指数都表明根际土壤细菌群落的丰富度及多样性都远高于真菌,不同竹种间微生物群落存在着一定的独特性和差异性,黄秆乌哺鸡竹根际土壤具有相对更丰富的细菌群落种类,雷竹根际土壤具有相对更丰富的真菌群落种类。
3.3 3个竹种根际微生物群落组成
3.3.1 根际细菌群落组成
3个竹种根际土壤细菌群落在各级分类水平上组分相似,构成比例差异不显著,如图2。3个竹种优势菌门为变形菌门(Proteobacteria)、绿弯菌门(Chloroflexi)、放线菌门(Actinobacteria)、酸秆菌门(cidobacteria),在毛竹根际土壤中占比分别为25.5%、20.2%、23%、20.7%,在雷竹根际土壤中占比分别为34.3%、6.8%、21.4%、28.1%,在黄秆乌哺鸡竹根际土壤中占比为28.7%、12.6%、11.9%、32.9%。
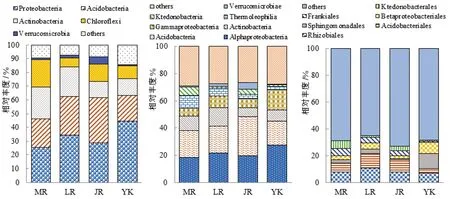
图2 3个竹种根际细菌在门(左)、纲(中)、目(右)水平上的相对丰度
3个竹种根际土壤优势菌纲均为α-变形秆菌纲(Alphaproteobacteria)及酸秆菌纲(Acidobacteria),在毛竹根际土壤中占比分别为18.4%、19.5%,在雷竹根际土壤中占比分别为21.7%、19.6%,在黄秆乌哺鸡竹根际土壤中占比分别为19.8%、28.4%;毛竹根际土壤优势菌目为根瘤菌目(Rhizobiales),占比8.2%,雷竹和黄秆乌哺鸡竹根际土壤优势菌目为根瘤菌目和酸秆菌目(Acidobacteriales),占比分别为11.1%、10.7%和8.1%、9.1%;雷竹根际土壤优势菌科为黄色秆菌科(Xanthobacteraceae)。
非竹林土壤与竹种根际土壤比较,细菌群落组分及构成比例差异并不显著。非竹林土壤优势菌群为变形菌门,α-变形秆菌纲,鞘脂单胞菌目(Sphingomonadales),鞘脂单胞菌科(Sphingomonas)。其中非竹林土壤根瘤菌目、酸秆菌目及弗兰克氏菌目(Frankiales)占比远低于竹种根际土壤,鞘脂单胞菌目及β-变形菌目(Betaproteobacteriales)占比高于竹种根际土壤。
3.3.2 根际固氮菌群落组成
在3个竹种中,毛竹和雷竹根际土壤固氮菌群落组分及构成比例都更接近,如图3和图4。毛竹、雷竹及黄秆乌哺鸡竹根际土壤主要构成菌门为变形菌门、蓝藻(blue-green algae)、厚壁菌门,这与刘彩霞等[14]对毛竹林固氮微生物群落研究结果一致;3个竹种的优势菌门均为变形菌门,占比分别为97.2%、97.5%、98.2%;在3个竹种中蓝藻在雷竹根际土壤占比最高,厚壁菌门在毛竹根际土壤占比最高,而雷竹不含厚壁菌门;3个竹种的优势菌纲均为α-变形秆菌纲,占比分别为77.2%、86.8%、93.8%,3个竹种中δ-变形菌纲、γ-变形菌纲(Gammaproteobacteria)、β-变形菌纲(Betaproteobacteria)、芽孢秆菌纲(Bacilli)在毛竹根际土壤占比均最高,雷竹根际土壤不含芽孢秆菌纲,黄秆乌哺鸡竹根际土壤不含δ-变形菌纲(Deltaproteobacteria);3个竹种的优势菌目均为根瘤菌目,占比分别为76.8%、86%、93.6%,3个竹种中雷竹根际土壤不含芽孢秆菌目,黄秆乌哺鸡竹根际土壤不含除硫单胞菌目(Desulfuromonadales)及假单胞菌目(Pseudomonadales);3竹种的优势菌科均为慢生根瘤菌科(Bradyrhizobiaceae),占比分别为76.4%、85.2%、89.7%;3竹种的优势菌属均为慢生根瘤菌属(Bradyrhizobium),占比分别为76.3%、85.2%、89.7%,黄秆乌哺鸡竹相较于毛竹与雷竹无地秆菌属(Geobacter)与假单胞菌属(Pseudomonas),而眉藻属(Calothrix)也远少于毛竹与雷竹。
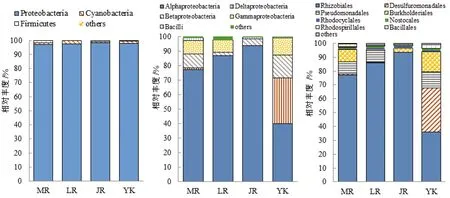
图3 3个竹种根际固氮菌在门(左)、纲(中)、目(右)水平上相对丰度
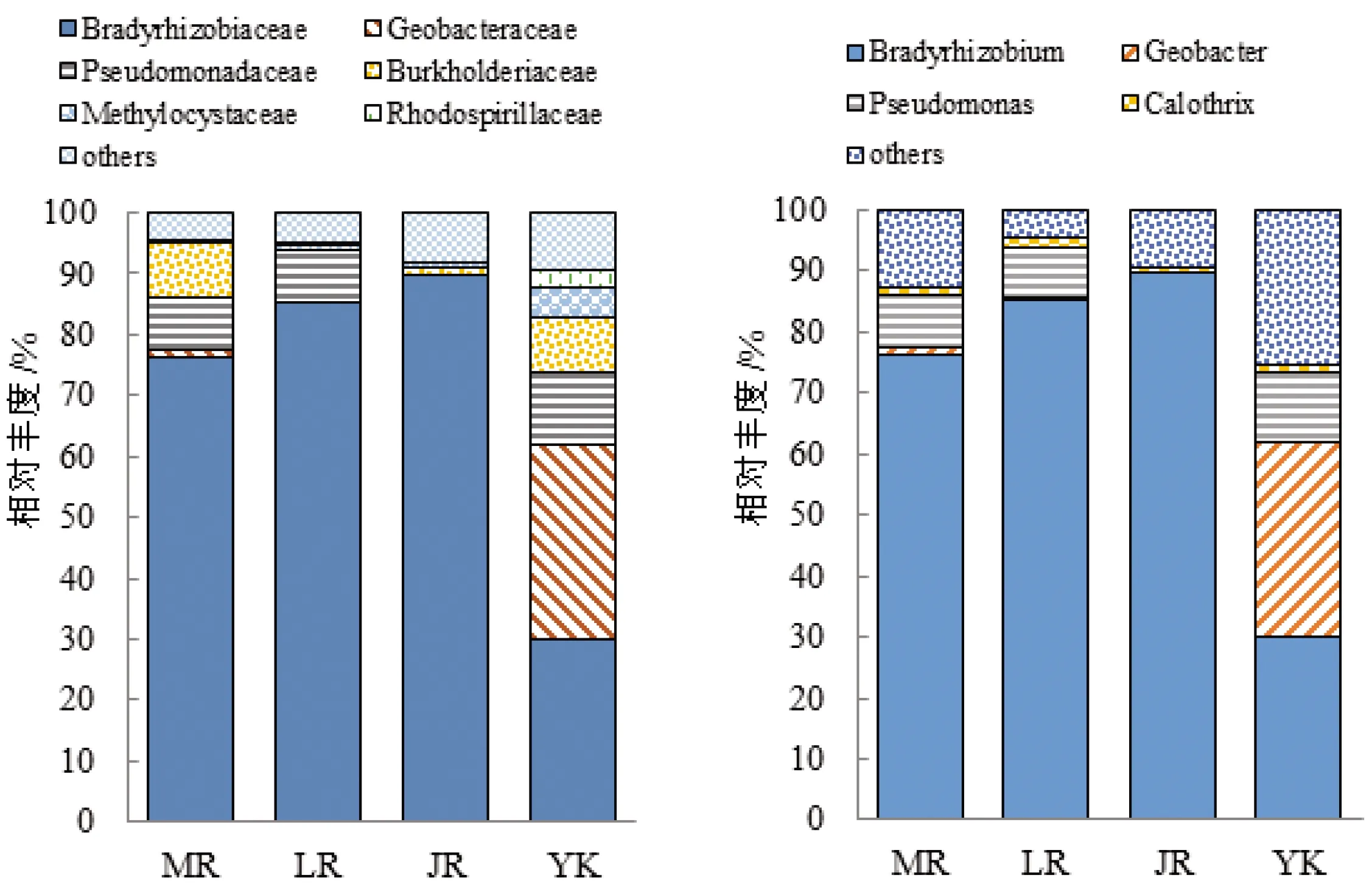
图4 3个竹种根际固氮菌在科(左)、属(右)水平上相对丰度
竹种根际土壤与非竹林土壤细菌群落比较,从纲水平上开始表现出较大差异,非竹林土壤α-变形秆菌纲及其下属级别占比均远低于竹种根际土壤,而δ-变形菌纲、β-变形菌纲、γ-变形菌纲及他们下属级占比都高于竹种根际土壤。
3.3.3 根际真菌群落组成
3个竹种根际土壤真菌群落在各级分类水平上组分相似,构成比例差异显著,如图5和图6。3个竹种根际土壤主要构成菌门为子囊菌门(Ascomycota)、担子菌门(Basidiomycota)、被孢霉门(Mortierellomycota),其中毛竹和雷竹根际土壤优势菌门均为子囊菌门,占比分别为83.8%、88.6%;黄秆乌哺鸡竹根际土壤优势菌门为担子菌门,占比46.8%。毛竹根际土壤优势菌纲为锤舌菌纲(Leotiomycetes),占比56.3%;雷竹根际土壤优势菌纲为散囊菌纲(Eurotiomycetes)和座囊菌纲(Dothideomycetes),占比分别为26.4%、32.1%;黄秆乌哺鸡竹根际土壤优势菌纲为伞菌纲(Agaricomycetes),占比43.6%。毛竹根际土壤优势菌目为柔膜菌目(Helotiales),占比56%,雷竹根际土壤优势菌目为散囊菌目(Eurotiales),占比26%,黄秆乌哺鸡竹根际土壤优势菌目为伞菌目(Agaricales),占比39.5%。毛竹根际土壤优势菌科为柔膜菌科(Helotiaceae),占比52.9%,是毛竹特有的菌科;雷竹根际土壤优势菌科为曲霉科(Aspergillaceae),占比24.1%,占比远高于毛竹及黄秆乌哺鸡竹根际土壤;黄秆乌哺鸡竹根际土壤优势菌科为鹅膏科(Amanitaceae),占比38.8%,是其特有菌科。毛竹根际土壤优势菌属为Claussenomyces,占比为52.8%;雷竹根际土壤优势菌属为青霉菌属(Penicillium)、曲霉菌属(Asperquillus),占比分别为13.5%、10.7%。
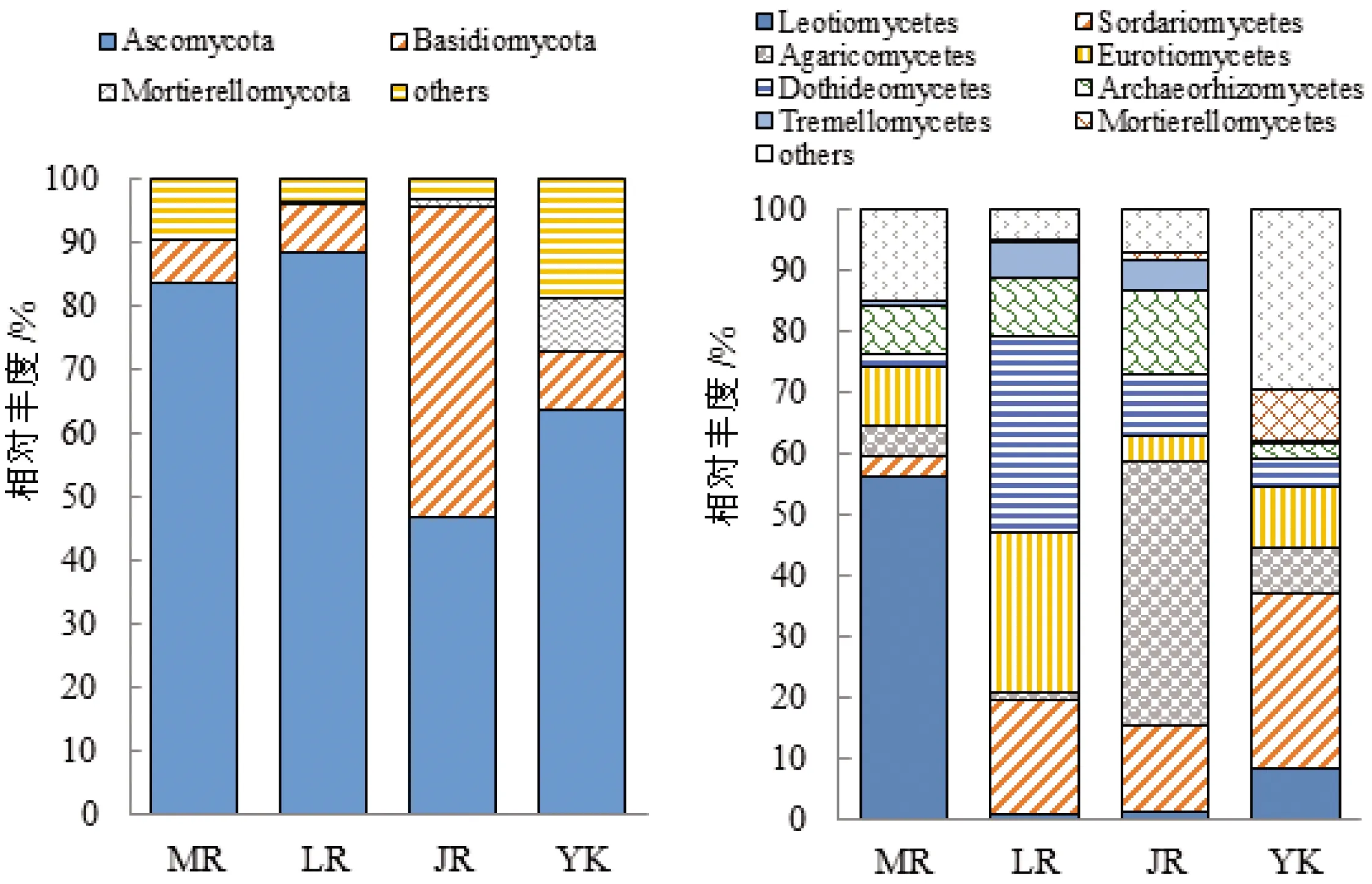
图5 3个竹种根际真菌在门(A)、纲(B)水平上相对丰度
非竹林土壤与竹种根际土壤比较,从门水平上开始表现出较大差异,其被孢霉菌门(8.3%)及其下级分类占比都远高于竹种根际土壤;粪壳菌纲(Sordariomycetes)(28.6%)占比也远高于竹种根际土壤,而古根菌纲(Archaeorhizomycetes)及银耳纲(Tremellomycetes)占比则低于竹种根际土壤。
4 讨论和结论
在本研究中根际土壤细菌群落的优势菌门与此前本课题组研究的东乡红星垦殖场雷竹林[15]、瑞金的毛竹林[16]土壤细菌群落相一致,同样以变形菌门、酸秆菌门、放线菌门、绿弯菌门这4个菌门为优势菌门,翟婉璐[17]发现未覆盖雷竹林土壤细菌群落以变形菌门、绿弯菌门、酸秆菌门为优势菌门,这些研究表明竹子这一物种的土壤细菌群落的优势菌门具有高度一致性。冯玮娜等[18]研究发现,四川牡丹根际细菌以变形菌门、厚壁菌门(Firmicutes)、酸秆菌门、硝化螺旋菌门(Nitrospirae)为优势菌门。Lundberg等[19]发现,在拟南芥根际土壤细菌以放线菌门、芽单胞菌门(Gemmatimonadetes)、变形菌门、拟秆菌门(Bacteroidetes)、厚壁菌门、酸秆菌门为优势菌门。周柳婷等[20]研究发现连栽木麻黄根际土壤细菌以酸秆菌门、放线菌门、疣微菌门(Verrucomicrobia)、变形菌门为优势菌门。乔清华[21]研究发现,棉花根际细菌以中变形菌门、疣微菌门、浮霉菌门(Planctomycetes)、酸秆菌门、放线菌门、拟秆菌门为优势菌门。综上可见,不同植物根际细菌的优势菌门存在一定程度的差异,放线菌门、变形菌门、酸秆菌门是竹类与以上植物间共有的优势菌门,说明这几个菌门可能是大多数植物的共同菌门,其中绿弯菌门是竹种较其他植物的特有优势菌门。
对竹种根际土壤与非竹林土壤细菌群落比较发现,慢生根瘤菌属在竹种根际土壤占比显著高于非竹林土壤,可能由于试验竹林成林后施肥量低,竹林生长需要一定量的氮,而慢生根瘤菌属为化能异养菌,可寄生植物根部固定大气氮,有的也可以游离状态固定氮,可为竹林供给一定量的氮,故慢生根瘤菌属在竹种根际土壤中占比较高。在3个竹种根际土壤真菌群落中青霉菌属、曲霉菌属在雷竹根际土壤中占比均最高,且这2个菌属在3个竹种根际土壤中占比都高于非竹林土壤,可能由于试验竹林施肥量低,而青霉菌属、曲霉菌属作为常见的溶磷真菌,能将土壤中植物难以吸收利用的磷转化为可吸收利用的形态,供给竹种利用,故他们在根际土壤中占比高,这一研究发现为溶磷菌筛选提供了菌株资源,也为后期研制生物肥料提供了理论基础。
本研究运用Illumina Hi Seq高通量技术,通过分析毛竹、雷竹及黄秆乌哺鸡竹根际微生物(细菌、真菌、固氮菌)群落结构,结果表明不同竹种根际土壤的微生物群落多样性具有一定差异性。从3个竹种Alpha多样性指数来看,雷竹根际土壤真菌及固氮菌丰富度及多样性均最高;毛竹根际土壤真菌丰富度及多样性均最低。从微生物群落组成来看,3个竹种根际土壤细菌群落在各级分类水平上组分相似,构成比例差异不显著,而非竹林土壤与竹种根际土壤比较,细菌群落组分及上构成比例也并无显著差异。在3个竹种中,毛竹和雷竹根际土壤固氮菌群落组分及构成比例都更接近,非竹林土壤与竹种根际土壤比较,从纲水平上开始表现出较大差异。3个竹种根际土壤真菌群落在各级分类水平上组分相似,构成比例差异显著。综上初步认为,不同竹种的根际微生物群落都有其特异性,因此在生产上应根据竹种的不同,选择施用合适的高效微生物肥料,促进竹种根际土壤优势菌群的形成,从而促进竹种生长,此研究结果为今后研制竹种专用生物肥料提供理论基础。
