制干辣椒果实辣椒素对干旱、盐及其双重胁迫的响应
2022-03-12吕慧吉雪花张中荣朱冉冉王世宁谢雪果袁雷
吕慧 吉雪花 张中荣 朱冉冉 王世宁 谢雪果 袁雷



摘 要:为明确干旱、盐等环境因素对制干椒辣椒素含量的影响,试验以红龙23号板椒为材料,从现蕾期开始设置不同土壤含水量65%(W1)、45%(W2)和NaCl(S1=120 mmol·L-1)及盐旱复合处理(W1S1、W2S1),利用高效液相色谱法分析转色期果实辣椒素素含量,并送样进行转录组测序。结果表明,W1处理下辣椒素含量较对照增加107.11%,W2处理时辣椒素含量下降73.01%;含盐处理均能提高辣椒素含量,其中W2S1处理的较对照高392%。盐、旱胁迫主要影响苯丙烷生物合成途径,其中HCT、4CL、CAD等基因表达受影响较大;聚类分析表明BCKDHE2、ENRb等基因主要响应干旱胁迫,CCR、GS2 主要响应盐胁迫。由上可知,适度干旱和盐胁迫均能提高辣椒素含量;干旱加剧时与盐分的复合效应更显著,对辣椒素积累的促进作用更大。
关键词:辣椒素;干旱胁迫;盐胁迫;双重胁迫;转录组
中图分类号:S641.3 文献标识码:A 文章编号:1673-2871(2022)02-078-07
Capsaicin of dry pepper fruit grown under drought, salt and combined stress condition
LÜ Hui, JI Xuehua, ZHANG Zhongrong, ZHU Ranran, WANG Shining, XIE Xueguo, YUAN Lei
(Agricultural College of Shihezi University/Key Laboratory of Special Fruits and Vegetables Cultivation Physiology and Germplasm Resources Utilization, Shihezi 832000, Xinjiang, China)
Abstract: Honglong 23 pepper was cultivated in pots stressed with drought and salt from budding to fruit color break to understand the effects of stress on capsaicin in pepper fruit for drying. Soil water contents were 65% (W1) and 45% (W2) of saturated content, NaCl concentration was 120 mmol·L-1(S1), and combined stresses were W1S1 and W2S1. Capsaicin contents were measured with HLPC, transcriptional data was analyzed by Personal Biotechnology Co Ltd. The results showed that the capsaicin content of pepper fruit under W1 treatment was 107.11% higher than that of the control, and the capsaicin content with W2 treatment was 73.01% lower than that of the control. Salt treatments (S1, W1S1, W2S1) increased the capsaicin content, and the capsaicin content in W2S1 treatment was 3.92 times of the control. Salt and drought stresses mainly affected the phenylpropane biosynthesis pathway of capsaicin, and had a great impact on the expression of HCT, 4CL, CAD genes in phenylpropane pathway. Cluster analysis showed that CPR, pAMT2 genes responded to the three stresses, while BCKDH E2, ENRb mainly responded to water stress, CCR and GS2 mainly responded to salt stress. Moderate drought and salt stress can increase the content of capsaicin in fruits. The combination of severe drought and salt had the most significant effect on capsaicin, while the mild drought and salt had little effect.
Key words: Capsaicin; Drought stress; Salt stress; Combined stress; Transcriptome
辣椒因其特殊的風味,备受人们的喜欢,而制干辣椒是新疆新兴的特色经济作物,随着种植规模的不断扩大,新疆已成为全国制干椒的原料生产基地。制干辣椒可用于提取辣椒素、辣椒红素等高附加值产品[1-2]。辣椒素类化合物是辣椒产生辛辣味的主要来源物质,同时也是评价制干辣椒果实品质的重要指标。研究表明辣椒素类物质有30多种,其中辣椒碱和二氢辣椒碱是辣椒素主要成分,占果实辣椒素总含量的90%[3-4]。辣椒素作为一种次生代谢产物,在农药研发、军事防御、食品加工、医学药理、临床研究等方面被广泛使用[5-7]。辣椒素主要在果实胎座中形成,其含量较低,供不应求,限制了其在市场中的大量使用[8]。因此,提高辣椒素含量是辣椒栽培和育种环节的重要目标。
辣椒素类物质的合成途径主要有两条:苯丙氨酸途径和支链脂肪酸途径[3,9-10]。多个基因参与了上述两条途径。目前已克隆的辣椒素代谢相关基因有PAL、C4H、CoAOMT、pAMT、KAS、ACL、FAT、ACS和AT3 等基因[11-12]。研究表明果实胎座中的C4H、CoAOMT、pAMT、KAS、AT3等基因的表达量与辣椒素含量呈正相关关系[13]。
辣椒素的含量不仅取决于基因型,还受到外界环境的影响。张海英[14]研究表明碱性盐胁迫可以显著提高红熟果中辣椒素的含量。新疆气候干旱、水资源短缺,而地表蒸腾强烈,因此盐碱土面积较大。实际生产中制干辣椒生长期面临着盐碱和干旱的双重胁迫[15]。目前,对辣椒素的环境影响研究主要集中在单一因素,且多在生理方面开展,对复合胁迫下的变化报道较少。
笔者以制约新疆制干辣椒生长的两个关键环境因子干旱和盐为出发点,采用高效液相色谱和转录组测序技术,分析辣椒素代谢相关基因在干旱、盐及双重胁迫下的表达,结合辣椒素含量的变化,明确其对干旱、盐及复合胁迫的响应差异,为农业生产中提高制干椒辣椒素含量提供理论指导。
1 材料和方法
1.1 材料
试验材料为加工辣椒品种红龙23号羊角椒,由新疆天椒红安农业科技有限公司提供。
1.2 試验设计
试验采用完全随机方式,2020年3—8月于新疆石河子大学农学院试验站日光温室进行(85°59′E,44°18′N)。将催芽后的红龙23号种子播种到体积比草炭∶蛭石=2∶1的穴盘中(尺寸为540 mm×280 mm),待其至6片真叶时定植于盆土体积比为草炭和蛭石(2∶1)∶园土(杀菌消毒)∶沙子=2∶4∶1的花盆(30 cm×45 cm)中,每盆质量8 kg。试验设置3个水分梯度:正常水分W0(土壤相对含水量为100%)、轻度干旱W1(土壤相对含水量为65%)、中度干旱W2(土壤相对含水量为45%);2个盐浓度:无盐S0(NaCl=0 mmol·L-1)、重度盐S1(120 mmol·L-1),共6组处理,分别为CK(W0S0)、干旱胁迫(W1、W2)、盐胁迫组(S1)、盐旱双重胁迫(W1S1、W2S1)组(具体见表1)。试验于花后7 d进行处理,每处理10盆,每盆3株重复。采用称质量法控制水分含量,每2~3 d称质量1次;盐分通过电导率控制,每2~3 d测定1次。处理后20 d于转色期采果实测定。各处理选第2~4层果实2~3个置于液氮速冻,-80 ℃保存。
1.3 指标测定
1.3.1 辣椒素含量的测定方法 参照厉志伟等[16]和张海英[14]的方法稍加改进。将辣椒果实置于105 ℃杀青30 min,而后于50 ℃烘干至恒重,在60 ℃水浴中超声辅助甲醇提取样品中辣椒素及二氢辣椒素。用LC-2010AHT高效液相色谱仪进行辣椒碱及二氢辣椒素的HPLC 测定。流动相体积比为甲醇∶超纯水=80∶20,检测波长为280 nm,流速为1.0 mL·min-1,柱温为30 ℃,进样量为10 μL。总辣椒素含量=(辣椒素含量+二氢辣椒素)/90%。
1.3.2 Illumina测序及测序数据的分析 委托南京派森诺基因科技有限公司进行Illumina测序。原始下机数据(Raw Data)经过过滤,去除带接头、低质量的Reads后得到高质量序列(Clean Data),将过滤后的Reads比对到参考基因组GCF_000710875.1_Pepper_Zunla_1_Ref_v1.0_genomic.fna进行对比,对比对上的Reads进行拼接,还原出转录本序列;分析比对所得基因功能注释,所用的数据库为GO(Gene Ontology),KEGG(Kyoto Encyclopedia of Genes and Genomes)。
根据对照组(CK)和各处理组间基因表达量对比进行差异表达分析,以表达差异倍数 |log2 FoldChange| > 1、显著性以p < 0.05作为筛选差异表达基因条件,并对差异表达基因进行代谢通路富集分析。
1.3.3 差异基因的qRT-PCR验证 选取差异表达基因并采用Trizol试剂提取果实的RNA,利用PrimeScript RT reagent Kit With gDNA Eraser试剂盒进行反转录,引物序列的设计及合成均由上海生工生物工程公司完成。每处理的每个基因3次重复,按照2-ΔΔCt相对定量法计算基因相对表达量。
1.4 数据处理
所得的试验数据采用Excel 2010、SPSS17.0软件进行统计分析,用Origin 9.0软件进行作图。聚类分析采用在线软件Clust Vis制作:https://biit.cs.ut.ee/clustvis。
2 结果与分析
2.1 干旱、盐及双重胁迫对辣椒果实中辣椒素含量的影响
由图1 可知,轻度干旱、盐及双重胁迫均有利于果实辣椒素的积累,且果实中辣椒素含量与对照(CK)相比差异显著。单一胁迫时,S1中辣椒素增加幅度较W1小;且辣椒素含量随着干旱程度的加剧而急剧减少,W1较对照显著提高了107.11%,W2较对照显著减少了73.01%。双重胁迫下,W1S1处理的辣椒素较对照显著增加了43.55%,较W1显著下降了30.69%,但与S1间差异不显著。W2S1的辣椒素含量较对照显著提高了3.92倍,较W2显著提高了12.69倍,较S1显著提高了2.61倍。上述结果说明适度的干旱和盐胁迫均能提高辣椒素含量,重度干旱时盐旱互作明显。
2.2 干旱、盐及双重胁迫下辣椒果实差异表达基因分析
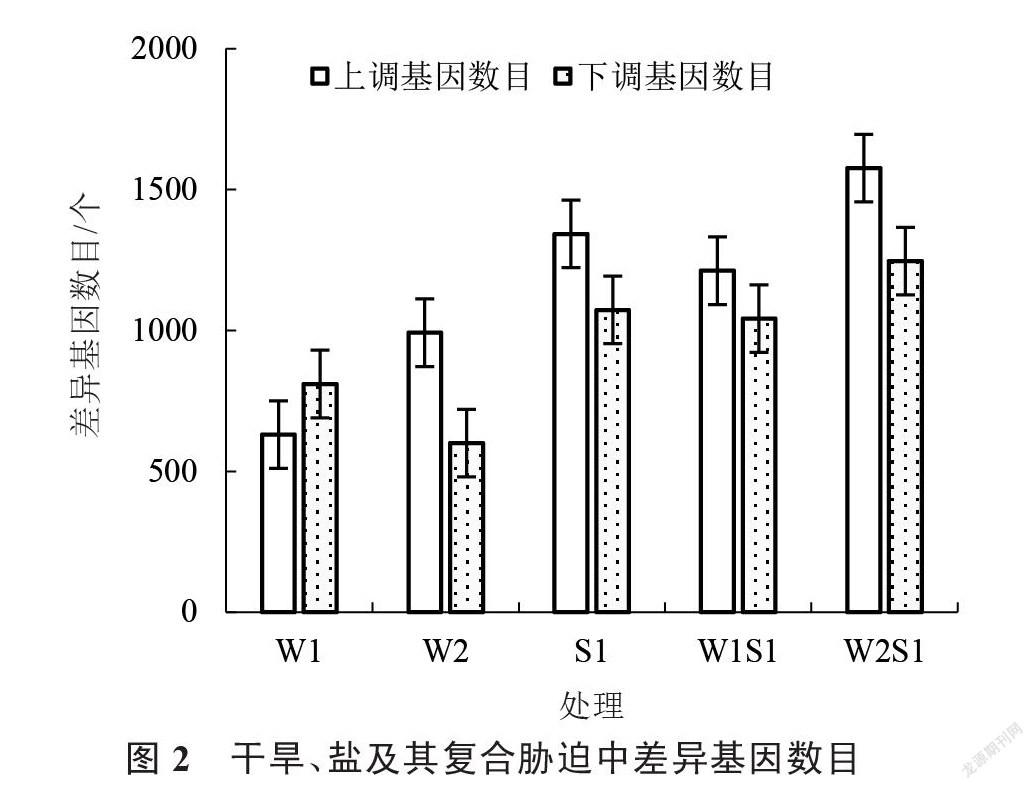
由图2可知 W1处理中有1440个差异基因,其中630个基因表达量上调,810个基因表达量下调。W2有1592个差异基因,其中992个基因表达量上调,600个基因表达量下调。S1检测出2416个差异表达基因,其中1343个上调,1073个下调。W1S1处理有2254个差异基因,其中1212个基因上调,1042个基因下调。W2S1处理有2822个差异基因,1576个基因表达量上调,1246个表达量下调。以上结果表明差异基因数目随干旱程度的加剧而增加。含盐处理(S1、W1S1、W2S1)的差异基因数量高于干旱处理(W1、W2)。
2.3 差异表达基因KEGG pathway分析
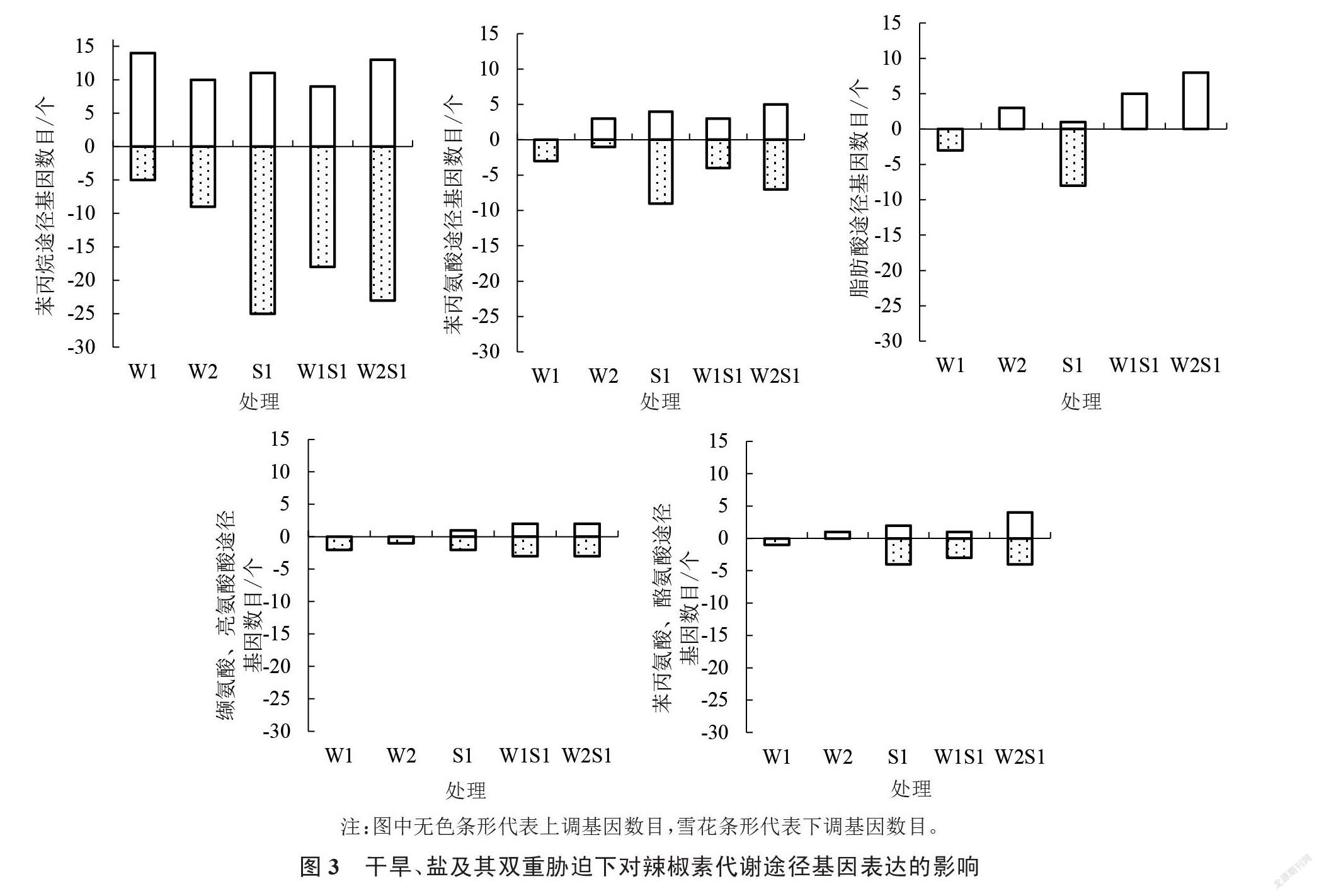
笔者集中对苯丙烷生物合成,苯丙氨酸代谢,脂肪酸生物合成,缬氨酸、亮氨酸和异亮氨酸的合成,以及苯丙氨酸、酪氨酸和色氨酸生物合成等与辣椒素代谢有关的途径进行分析。如图3所示,干旱、盐及其双重胁迫下与辣椒素有关的途径主要集中在苯丙烷生物途径上,其次为苯丙氨酸途径和脂肪酸途径。整体上W2S1对5条途径都较敏感,W2处理对各途径的响应较缓慢。在苯丙烷合成途径中,W1、W2S1的上调基因数目较多,而W2、W1S1上调基因个数基本一致;S1和W2S1下调基因数目较多。S1、W2S1在苯丙氨酸代谢途径中上调和下调的差异基因均较多。在脂肪酸合成途徑中,W1S1和W2S1的上调基因数目均较多,W1和S1的下调基因数目较多。在缬氨酸、亮氨酸和异亮氨酸合成途径中,干旱胁迫处理的差异基因均下调;含盐处理S1、W1S1和W2S1的差异基因上调、下调均较多。S1、W2S1处理的差异基因在苯丙氨酸、酪氨酸和色氨酸途径中上调和下调基因较多。以上表明S1、W1S1及W2S1等含盐处理的差异基因在各途径中的富集程度高于W1、W2等单一干旱胁迫,其中W2S1处理的果实辣椒素相关基因在各途径富集程度均较高,除苯丙烷途径外,W1和W2处理的辣椒素差异基因在其他途径富集程度不高。
2.4 辣椒素代谢相关基因的聚类分析
研究分析了43个与辣椒素代谢相关的基因在干旱胁迫、盐胁迫及其双重胁迫下的表达量,并对各处理下的基因进行表达谱聚类分析,由图4可知,可将辣椒素相关基因分为A、B两组。
A组共有13个基因,按照表达模式又分为4小类A1、A2、A3、A4。
A1组:包含BCKDH E2基因。该基因位于支链脂肪酸途径的中游。该基因在W1处理下的表达量高于其他处理,在W2中表达量最低。BCKDH E2在含盐处理S1、W1S1、W2S1的表达量均低于CK。
A2:CAD、SAMSyn、ACS2等,这组基因在CK 和W1S1处理中表达量均较低。单一干旱或单盐胁迫的CAD基因表达量高于CK和复合胁迫。SAMSyn、ACS2基因在W1、W2中的表达量高于CK,同时随着干旱程度的增加而增加。SAMSyn、ACS2基因在S1、W2S1的表达量高于CK,在W1S1中表达量低于CK。在S1、W1S1、W2S1等含盐处理中,SAMSyn、ACS2基因表达量变化较大,先下降后上升。
A3组:该组只包含ACS1基因。该基因在W1S1处理中表达量最高,而在其他处理中表达量较低。
A4:包括pAMT2、PAT、CCR、Fdx-GOGAT、GS2、CPR、ALS、AHRI等基因,上述基因在W2S1复合胁迫下表达量较高,其次为W1S1处理,在CK和W1处理中表达量较低。
B组共有30个基因,按照表达模式又分为5小类B1、B2、B3、B4、B5。
B1:BCAT是支链脂肪酸途径的中游基因,在CK中表达量最高,而在W1中表达量最低,在其他处理组的表达量均较低。
B2:包括ENRb、ADT、IPMI、IPMS、IPMDH基因。这组基因在W1处理下表达量很高,其次为W2处理。在S1、W1S1、W2S1等含盐处理中表达量均较低。
B3:HCT、KasI、4CL等基因在W2中的表达量最高,在W2S1和S1中表达量较低。
B4:KasIIIb、CCoAOMT、BCKDH E1a为一类,其在CK中表达量较高。其中,BCKDH E1a表达量最高,3个基因在W1、S1、W2S1处理中表达量均较低。其中,CCoAOMT在W2S1中表达量最低。
B5:包括α-CT、DH、C4H、FatB、PDH E1a、PDH E1b、KR、PDH E2、ENRa、PDH E3、BC、PAL、BCCP、KasIIIa、MCAT、C3H、BCKDH E3、CM1等18个基因。这组基因在CK、W2、W1S1中表达量较高,在S1、W2S1中表达量较低。在W1、W2处理中,α-CT、DH、C3H的表达量随干旱程度的增加而减少。在含盐处理中,α-CT、DH、C4H、FatB、PDH E1a、PDH E1b、KR、PDH E2、ENRa、PDH E3、BC、PAL、BCCP、KasIIIa、MCAT、BCKDH E3、CM1在W1S1的表达量均高于S1、W2S1处理。α-CT、DH、C4H、FatB、PDH E1a、PDH E1b、KR、PDH E2、ENRa、PDH E3、BC在W1S1的表达量均高于W1,PAL、BCCP、KasIIIa、MCAT、C3H、BCKDH E3、CM1在W1S1的表达量则低于W1。α-CT、DH、C4H、FatB、PDH E1a、PDH E1b、KR、PDH E2、ENRa、PDH E3、BC、PAL、BCCP、KasIIIa、MCAT、C3H、BCKDH E3、CM1基因在W2的表达量均高于W2S1。轻度干旱时,加入盐分后,可以提高α-CT、DH、C4H、FatB、PDH E1a、PDH E1b、KR、PDH E2、ENRa、PDH E3、BC等的表达量,当干旱胁迫加剧后,则会降低基因的表达量。
2.5 辣椒素代谢相关差异基因分析
在43个辣椒素代谢相关基因中以差异倍数 |log2FoldChange| > 1、显著性p < 0.05为标准,筛选各处理组中符合条件的差异基因并作分析(表3)。
在处理W1中筛选出3个表达量显著下调的辣椒素代谢相关基因KasI、BCAT、BCKDH E1a;W2處理中,有4个差异基因,BCKDH E1a显著下调,KasI、HCT、GS2显著上调;S1处理中有13个差异基因,分别为C4H、KasI、KasIIIa、PAL、BCKDH E1a、CM1、GS2、PDH E1a、PDH E1b、PDH E3、BC、ENRa、MCAT。其中,GS2基因表达量显著上调,其余表达量显著下调;BCAT、ACS1、GS2在W1S1处理下差异显著,其中,BCAT表达量显著下调,其余下调;在处理W2S1中,GS2表达量显著上调,KasI、PAL、C3H、BCKDH E1a、CM1、PDH E3、BC、ENRa、MCAT基因表达量下调。
2.6 差异基因的实时荧光定量qRT-PCR
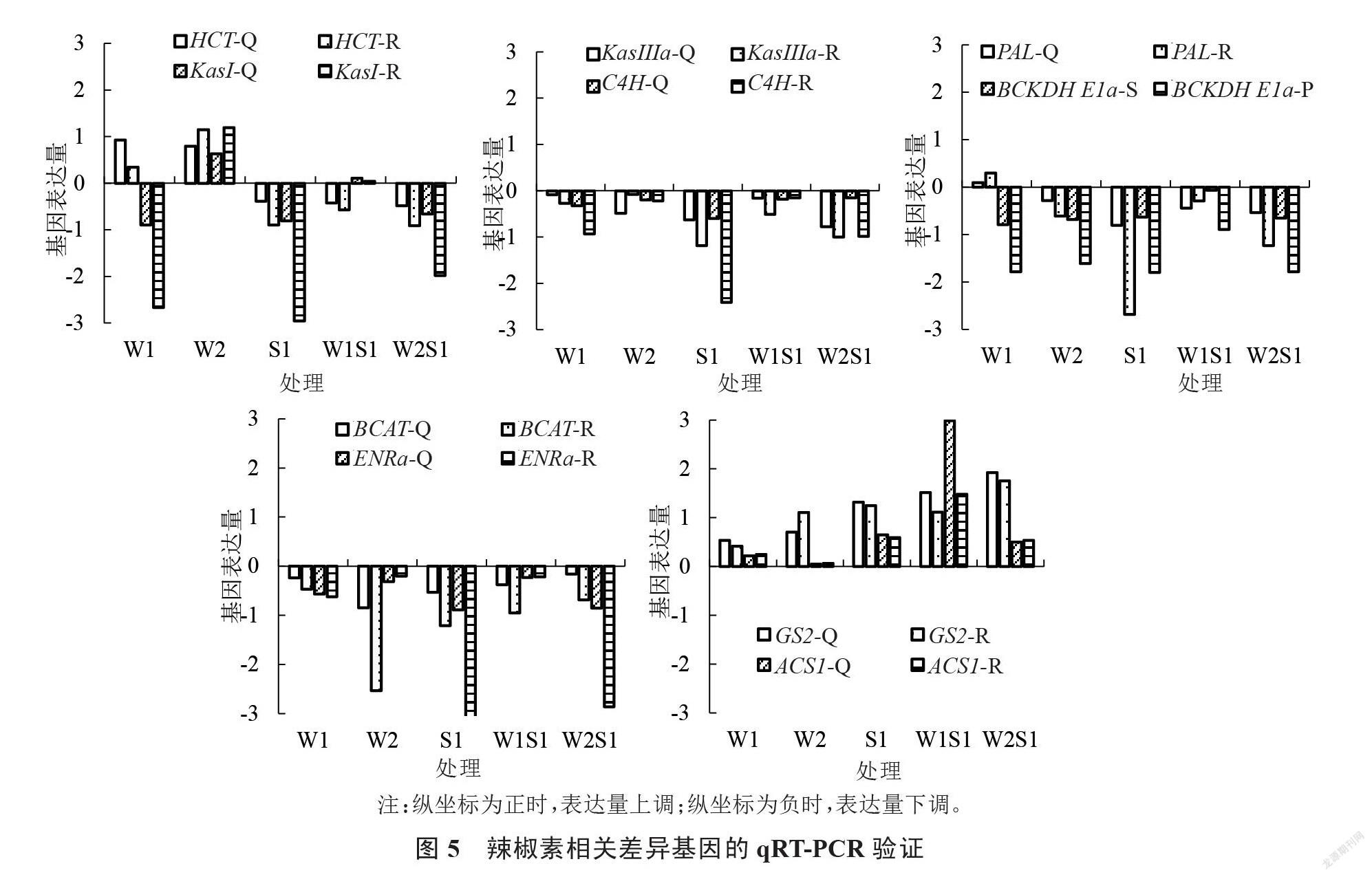
为确保转录组测序数据的准确性,从差异显著的基因中随机选取10个基因HCT、KasI、KasIIIa、C4H、PAL、BCKDH E1a、BCAT、ENRa、GS2、ACS1进行表达量验证。由图5可知,10个辣椒素合成相关基因在干旱胁迫、盐胁迫、旱盐胁迫下均有表达,且表达量有差异。HCT基因在干旱胁迫下表达量上调,在盐胁迫和旱盐双重胁迫时表达量均降低。KASI基因在W2和W1S1处理中表达量上调,其余处理表达量下调。KasIIIa、PAL、C4H、BCKDH E1a、BCAT、ENRa基因在各处理下表达量均降低。GS2、ACS1在各处理下表达量均上升。以上各基因的表达量均与转录组测序得到的趋势一致,确保了测序结果的准确性。
3 讨论与结论
辣椒是广受消费者欢迎的一种蔬菜和调味品,在世界范围内广泛种植。辣椒素类物质作为辣椒中的辣味成分,其含量受品种影响的同时还会受到栽培措施和环境因素的制约[17-19]。干旱和盐碱是新疆制干辣椒栽培过程中面临的主要环境胁迫因子[3,10,20]。本试验结果表明,辣椒素含量在土壤含水量为75%时较对照明显提升,在含水量为45%时显著减少,并且辣椒素含量在盐胁迫及旱盐双重胁迫下均能得到显著提升,这可能是由于植株受到较大程度的干旱胁迫后,气孔关闭,光合下降,碳水化合物的合成不足,从而减少了辣椒素的合成,或者辣椒素在受到盐胁迫后,产生了较多能为辣椒素提供底物的防御物质。这与彭琼等[21]、Arrowsmith[22]对辣椒素在干旱、盐胁迫下的研究结果相似。
辣椒素类物质合成需要在众多的基因和基因家族的共同参与下才能完成,而目前对辣椒素的合成代谢调控认识有限,还需进一步研究。笔者发现含盐处理(S1、W1S1、W2S1)在辣椒素代谢通路中的富集程度要高于干旱处理,且辣椒素代谢相关基因在干旱、盐及旱盐胁迫中的表达量存在差异。其中,PAL在含盐处理中的表达量随水分的减少而降低,说明PAL基因的表达与水分关系密切。有研究表明C4H、ACS、KAS及CCoAOMT基因的表达与辣椒素的积累呈正相关[23-24],但本试验中以上几个基因与辣椒素的积累规律性不强。CCoAOMT基因在胁迫下的表达量均较低,KasI、KasIIIa、KasIIIb在含盐处理中表达量低或几乎不表达,说明以上基因受干旱或盐的影响较大。ENRb、ADT、IPMI、IPMS、IPMDH基因在W1中表达量较高,在W2中表达量较低,说明这些基因对水分较为敏感。HCT、KasI、4CL基因在W2中表达量较高,其余胁迫中较低,说明干旱能刺激基因的表达。SAMSyn、ACS2、pAMT2、PAT、CCR、Fdx-GOGAT、GS2、CPR、ALS、AHRI基因在W2S1中表达量高于其他胁迫,说明在干旱和盐胁迫的共同刺激下促进其表达。但对这43个基因在干旱、盐及双重胁迫下的作用机制有待进一步研究。
综上所述,辣椒素含量在适度干旱、盐胁迫或旱盐胁迫下均能明显提升,重度干旱抑制其合成。含盐处理对代谢通路富集更显著,辣椒素相关基因在干旱、盐及旱盐胁迫中的表达模式存在明显的差异。该结果为农业生产中提高辣椒素含量以及辣椒的栽培管理提供了理论指导。
参考文献
[1] 李宁,杨涛,帕提古丽,等.适于新疆制干类型辣椒种质资源遗传多样性的SRAP和SCoT分析[J].分子植物育种,2017,15(8):3331-3340.
[2] 佚名.《中国线辣椒》简介[J].辣椒杂志,2017,15(1):19.
[3] 张婧,颉建明,郁继华,等.辣椒素类物质的生物合成影响因素及其生理功能研究进展[J].园艺学报,2019,46(9):1797-1812.
[4] ARCE-RODRIGUEZ M L,OCHOA-ALEJO N.Biochemistry and molecular biology of capsaicinoid biosynthesis:recent advances and perspectives[J].Plant Cell Reports,2019,38(9):1017-1030.
[5] CHRISTOPHER A,DENNIS J,GAROLD S,et al.Determination of capsaicin,dihydrocapsaicin and nonivamide in self-defense weapons by liquid chromatography–mass spectrometry and liquid chromatography–tandem mass spectrometry[J].Journal of Chromatography A,2001,912(2):259-267.
[6] YUAN L J ,QIN Y,WANG L,et al.Capsaicin-containing chili improved postprandial hyperglycemia,hyperinsulinemia and fasting lipid disorders in women with gestational diabetes mellitus and lowered the incidence of large-for-gestational-age newborns[J].Clinical Nutrition,2016,35(2):388-393.
[7] CLARK R,LE S H.Anticancer properties of capsaicin against human cancer[J].Anticancer Research,2016,36(3):837-844.
[8] 陳俊琴.辣椒果实中辣椒素生物合成相关物质及外源多胺调控的研究[D].沈阳:沈阳农业大学,2015.
[9] 狄云,蒋健箴,石正强,等.辣椒果实成熟过程中辣椒素的降减[J].食品科学,2000,21(6):19-22.
[10] 张正海,毛胜利,王立浩,等.辣椒的辣味遗传控制与辣椒素生物合成研究进展[J].园艺学报,2014,41(9):1821-1832.
[11] 邓明华,文锦芬,朱海山,等.辣椒素生物合成相关基因研究进展[J].辣椒杂志,2012,10(3):1-5.
[12] 李彦磊.辣椒果实辣椒素积累和生物合成相关基因表达分析[D].长春:吉林大学,2013.
[13] ARCE-RODRIGUEZ M L,OCHOA-ALEJO N.An R2R3-MYB transcription factor regulates capsaicinoid biosynthesis[J].Plant Physiology,2017,174(3):1359-1370.
[14] 张海英.盐胁迫和碱胁迫对制干辣椒生长发育及果实品质的影响[D].新疆石河子:石河子大学,2019.
[15] 罗廷彬,任崴,谢春虹.新疆盐碱地生物改良的必要性与可行性[J].干旱区研究,2001,18(1):46-48.
[16] 厉志伟,王盛莉.高效液相色谱法测定辣椒中辣椒素、二氢辣椒素[J].中国调味品,2017,42(11):123-126.
[17] YU S,YU Y C,NI L T.Capsaicin biosynthesis in water-stressed hot pepper fruits[J].Botanical Bulletin of Academia Sinica,2005,46:35-42.
[18] 袁雷,杨涛,张国儒,等.辣椒果实中辣椒素的研究进展[J].中国瓜菜,2021,34(11):1-9.
[19] 苏丹,蓬桂华,韩世玉,等.复合型肥料对辣椒素及其相关次生代谢物质积累的影响与相关分析[J].中国瓜菜,2016,29(11):19-22.
[20] 胡明芳,田长彦,赵振勇,等.新疆盐碱地成因及改良措施研究进展[J].西北农林科技大学学报(自然科学版),2012,40(10):111-117.
[21] 彭琼,童建华,柏连阳,等.干旱胁迫对辣椒果实中辣椒素、二氢辣椒素及VC含量的影响[J].中国蔬菜,2015(12):44-47.
[22] ARROWSMITH S,EGAN T P,MEEKINS J F,et al.Effects of salt stress on capsaicin content,growth and fluorescence in a Jalapeo cultivar of Capsicum annuum (Solanaceae)[J].Bios,2012,83(1):1-7.
[23] ZHANG Z X,ZHAO S N,LIU G F et al.Discovery of putative capsaicin biosynthetic genes by RNA-Seq and digital gene expression analysis of pepper[J].Scientific Reports,2016,6(8):253-706.
[24] CURRY J,ALURU M,MENDOZA M,et al.Transcripts for possible capsaicinoid biosynthetic genes are differentially accumulated in pungent and non-pungent Capsicum spp.[J].Plant Science,1999,148(1):47-57.
