贺兰山不同海拔植被下土壤微生物群落结构特征
2022-03-10马进鹏庞丹波万红云陈高路李学斌
马进鹏,庞丹波,陈 林,万红云,陈高路,李学斌,*
1 宁夏大学西北退化生态系统恢复与重建教育部重点实验室, 银川 750021 2 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地, 银川 750021 3 宁夏大学农学院, 银川 750021 4 宁夏大学生态环境学院,银川 750021
土壤微生物作为生态系统中的分解者,是土壤生态系统的重要组成成分,在生态系统的初级生产、物质循环、能量流动等过程起着重要的作用,同时也与地上植被的多样性有着密切的关系[1—2];作为连接土壤植物之间的纽带,微生物是生物地球化学循环的重要驱动因素[3—4]。在森林生态系统中,土壤微生物的分布受空间因素的影响有较为明显的差异。研究表明,土壤表层因其物种交互作用强,资源丰富,是微生物群落最为丰富、群落特征最明显的区域[5]。同时,土壤微生物是生态系统功能的敏感指标,其组成和数量可以反映土壤的质量,其多样性及群落结构可以有效反映土壤系统的稳定,能很好地指示森林生态环境和系统功能的变化[6]。
早在2008年,Bryant等[7]研究,首次发现土壤酸杆菌多样性随海拔上升而线性减小,后续越来越多的学者研究土壤微生物随海拔的分布格局[8—9]。近年来,随着土壤微生物测定技术的发展,土壤微生物随海拔的分布格局研究不断涌现,研究焦点不断创新,研究结果更加多样[10]。在长白山苔原带,Shen等[11]研究发现土壤细菌丰富度随海拔升高而降低;同时,人类活动也会影响土壤微生物随海拔分布格局,张倩等[12]在云和梯田研究发现细菌多样性在未受干扰的自然条件下随海拔升高而减小,而在水稻土中随海拔升高先升高后减小。在山地森林生态系统中海拔的变化对植被组成和土壤性质的影响尤为明显,而土壤又是微生物赖以生存的场所[13]。土壤微生物与气候、植被类型及土壤性质等密切相关,而海拔变化往往在较短范围内引起温度、降雨、植被类型和土壤性质等生物和非生物因子的显著变化,因此与海拔相关的环境条件强烈影响着土壤微生物的组成及群落结构[3,14]。在全球变化的背景下,研究土壤微生物的群落随海拔变化的特征,对于探索森林生态系统物质循环和能量流动对环境因子变化的响应有着重要的作用[15]。
贺兰山位于我国温带草原区和荒漠区的分界处,在地理位置上具有典型性,在植物区系和植被带上具有过渡性,植被类型具有复杂多样性,生态系统有较高的脆弱性[16]。贺兰山植被类型具有明显的垂直地带性,随海拔的升高,植被类型依次为荒漠化草原、山地疏林草原、山地针叶林和亚高山灌丛草甸或高山草甸。现今,贺兰山植物多样性的研究越来越受到重视,但是对其不同海拔梯度土壤微生物的群落结构和分布特征研究较少[16—17]。磷酸脂肪酸(PLFA)是磷脂的构成成分,自然生理条件下相对恒定,在土壤中的多样性和丰度可以表示特定生物类群的多样性和丰度[18]。不同类群的微生物能通过不同的生化途径合成不同的磷酸脂肪酸,部分磷酸脂肪酸可以作为分析微生物生物量和微生物群落结构等变化的生物标记[19—20]。本文采用PLFA图谱法分析贺兰山不同海拔梯度土壤微生物群落结构特征,探讨土壤微生物群落随海拔的变化规律及影响因素,意在为贺兰山荒漠-森林生态系统的养分循环提供依据。
1 材料与方法
1.1 研究区概况
贺兰山位于银川平原和阿拉善高原之间(地处38°27′—39°30′N,105°41′—106°41′E之间),山体弧立,成南北走向,主峰海拔3556m。贺兰山西边和北边为阿拉善戈壁荒漠,东北为宁夏平原。贺兰山处于典型大陆性气候区域范围内,具有山地气候特征。气候变化大,年均气温-0.8℃,年均降雨量420mm,年均蒸发量2000mm,贺兰山降雨量有明显的垂直分布规律,平均每上升100m,降雨量增加13.2mm,其降水主要集中在6—8月份,占全年降水量的60%—80%[17,21]。
1.2 样品采集
在贺兰山东坡1300—2300m海拔范围内物种丰富度最高,林型变化较大,因此沿海拔高度选取以短花针茅为建群种的荒漠草原、以蒙古扁桃为建群种的山地旱生灌丛、以油松为建群种的温性针叶林,以油松、杜松、山杨为优势种的针阔叶混交林,以青海云杉为建群种的寒温性针叶林等5种代表性的典型植被带,在每个植被带内按等高线设置3个取样点(样地基本情况见参考文献[22]),每个取样点去除地表的枯落物和腐殖质或剥离表土后用直径4cm的土壤取样器采用五点取样法取0—10cm的土样5个,混合后作为该取样点的土壤样品并用自封袋密封之后放入冰盒带回实验室。去除植物根系和石块,过2mm筛后分成两份,一份自然风干进行土壤理化性质测定,另一份放入-80℃冰箱进行土壤微生物测定。
1.3 测定方法
1.3.1土壤理化性质测定
采用pH计测定土壤pH(水土比2.5∶1);土壤容重采用烘干法;土壤含水量用环刀法测定;土壤有机碳采用重铬酸钾外加热法测定;土壤全氮采用凯氏定氮法;全磷采用HClO4-浓H2SO4外加热消煮法、分光光度法[23]。
1.3.2土壤微生物群落结构测定
土壤微生物群落结构多样性采用磷酸脂肪酸法(PLFA)[24],采用修正的Bligh-Dyer方法进行脂类提取和磷脂脂肪酸分析:土样用体积比为1:2:0.8的氯仿:甲醇:柠檬酸缓冲液振荡提取总脂类,然后经SPE硅胶柱分离得到磷脂脂肪酸,将得到的磷脂脂肪酸进行碱性甲醇化,用Agilent 6890N气相色谱仪分析PLFA的成分,内标为正十九烷酸甲酯(19:0)[25]。本研究选取了8种指标,其中表征的磷酸脂肪酸如下:非特异性细菌(12:0,14:0,15:0,16:0,17:0,18:0,19:0,20:0,24:0);革兰氏阳性细菌(a12:0,i13:0,a13:0,i14:0,a14:0,i15:0,a15:0,i16:0,a16:0,i17:0,a17:0,i17:1ω9c);革兰氏阴性细菌(16:1ω7c DMA,17:1ω8c,17:0cycloω7c,19:0cycloω7c,22:1ω9c);真菌(18:2ω6c);放线菌(10Me16:0,10Me17:0,10Me18:1ω7c,10Me17:1ω7c);原生动物(20:4ω6c,20:5ω3c);16:1ω5c,18:1ω9c)。因为16:1ω5c在G-和AMF(丛枝菌根真菌)中同时存在[26],18:1ω9c在G-和真菌中同时存在[27],且所占比重较大,同时,16:1ω5c是表征AMF的一个重要磷酸脂肪酸,18:1ω9c是表征真菌的重要脂肪酸,所以选取16:1ω5c和18:1ω9c作为独立的指标来表征微生物的数量。另外我们用磷酸脂肪酸的相对丰度分析反映微生物群落组成[28]。
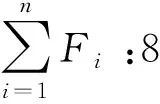
1.4 数据处理
用SPSS25.0进行数据处理,选择单因素方差分析(ANOVA)和最小差异性显著(LSD)进行差异性显著检验,用Origin2019制图;采用Canoco5进行主成分分析(PCA)和冗余分析(RDA)。
2 结果分析
2.1 不同海拔梯度对土壤理化性质的影响
随海拔的变化(如图1),土壤养分在不同海拔之间存在显著差异(P<0.05),随海拔的升高土壤有机碳、土壤全氮含量显著增加,土壤全磷含量变化随海拔升高先升高再降低再升高,在海拔2139m处最小,海拔1650m和海拔2438m处高于其他海拔;土壤化学计量比在不同海拔之间也存在差异(P<0.05),C/N在海拔2139m处最高,N/P、C/P在海拔2249m处较高;土壤理化指标在不同海拔之间存在显著差异(P<0.05),土壤pH随海拔的升高而降低,土壤容重随海拔升高显著降低,土壤含水量随着海拔升高而增加。土壤理化各指标的最大值和最小值均不出现在同一海拔高度。

图1 不同海拔梯度土壤理化性质Fig.1 Physical and chemical properties of soil at different altitudes 不同小写字母表示不同海拔间的差异显著(P<0.05)
2.2 不同海拔梯度对土壤微生物的影响
2.2.1不同海拔梯度对土壤微生物群落组成的影响
随海拔的变化,特征土壤微生物含量在不同的海拔之间存在差异(如表1)。其中细菌、真菌、放线菌、革兰氏阳性细菌、革兰氏阴性细菌均表现出在海拔2139m处含量最高,在海拔1380m处含量最低,而原生动物则在海拔1650m处含量最高。不同海拔梯度土壤微生物总量整体表现为,海拔2139m>2438m>1650m>2249m>1380m。
不同海拔下土壤微生物真菌/细菌,革兰氏阳性细菌G+/革兰氏阴性细菌G-存在差异(如图2)。其中真菌/细菌在海拔2139m处最高,在海拔1380m处最低;而G+/G-则在海拔1380和1650m处最高,在海拔2139m处最低。真菌/细菌随着海拔升高先升高后下降,而G+/G-随着海拔升高先下降后升高。
2.2.2不同海拔梯度对土壤微生物群落丰度的影响
随海拔的变化,土壤微生物群落丰度在不同海拔之间存在差异(如图3)。非特异性细菌和真菌在不同海拔的相对丰度存在显著差异(P<0.05),非特异性细菌在海拔1650m处丰度要高于其他海拔,真菌(18:2ω6c)在海拔2139m处丰度最高。革兰氏阳性细菌丰度在不同海拔之间无显著差异(P<0.05),在海拔1380m和海拔2249m处丰度最高,在海拔2139m处丰度最低;而革兰氏阴性细菌在不同海拔之间存在差异(P<0.05),在海拔2139m和海拔2438m处的丰度要高于其他海拔。18:1ω9c在不同海拔之间无显著差异(P<0.05),但相比而言,在海拔2139m处的丰度要高于其他海拔。16:1ω5c在不同海拔之间存在显著差异(P<0.05),在海拔1380m和海拔2438m处的丰度显著低于其他海拔。放线菌和原生动物在不同海拔之间无显著差异(P<0.05),但相比而言,在海拔1380m处放线菌的丰度高于其他海拔,在海拔1650m处原生动物的丰度高于其他海拔。总体而言,革兰氏阳性细菌和放线菌丰度随着海拔升高先下降后升高;非特异性细菌、真菌和16:1ω5c随着海拔升高先升高后下降。

表1 不同海拔梯度特征土壤微生物的含量

图2 不同海拔下真菌/细菌、G+/G-Fig.2 Fungi/bacteria、G+/G- at different altitudes不同小写字母表示不同海拔之间的差异显著(P<0.05)
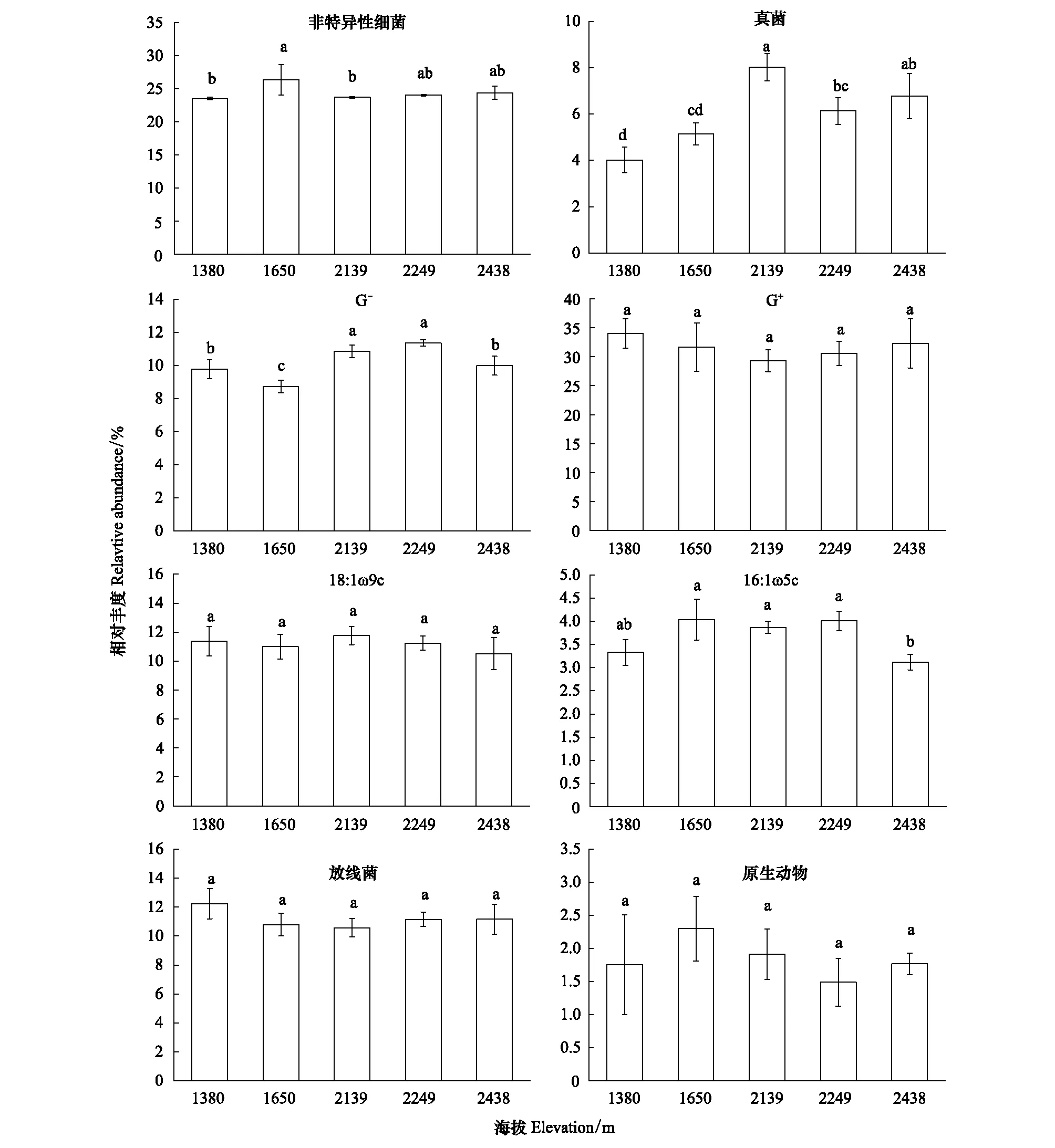
图3 不同海拔梯度不同微生物类群的相对丰度Fig.3 Relative abundance of different microbial groups at different altitude gradients不同小写字母表示不同海拔间的差异显著(P<0.05)

图4 土壤微生物群落相对丰度的主成分分析Fig.4 Principal component analysis of relative abundance of soil microbial community
对不同海拔的微生物相对丰度进行主成分分析,结果见图4。根据土壤微生物相对丰度二维排序图,4个轴对微生物相对丰度差异的总解释高达95.46%,第1排序轴贡献率为52.76%,第2排序轴贡献率为28.72%,前两个轴累计贡献率为81.48%,对微生物相对丰度的差异解释能力强。与第1排序轴相关性较强的微生物为G+、G-、真菌和18:1ω9c,相比之下,16:1ω5c的相关性较弱。与第2排序轴相关性较强的微生物为放线菌、原生动物和非特异性细菌。海拔取样点距离的大小表示样点间微生物群落结构的相似程度,距离越近则相似程度越高,可以看到,海拔1380m和海拔2139m处微生物群落结构异质性相比海拔1380m与其他海拔之间要高,而海拔2249m、2438m和海拔1650m处土壤微生物群落结构相似性较高。
2.3 土壤微生物群落特征与土壤理化性质的关系
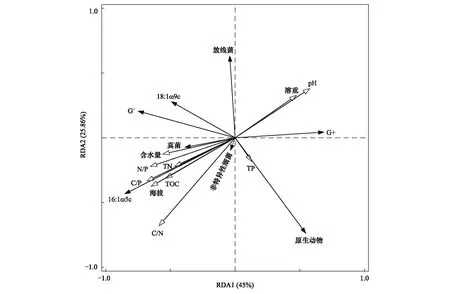
图5 不同海拔土壤微生物群落结构与土壤因子冗余分析Fig.5 Analysis of soil microbial community structure and soil factor redundancy at different altitudes
对不同海拔土壤微生物群落组成与土壤因子进行冗余分析(RDA)(图5),各因子的解释量和显著性检验结果(表2)。RDA轴1和轴2共同解释变量的81.81%,分析可知,土壤微生物群落结构组成与土壤理化性质之间相关性较好。真菌与G-、非特异性细菌正相关,与G+、放线菌和原生动物负相关;G+与原生动物正相关,与其他微生物负相关;G-与放线菌正相关,与原生动物负相关,与非特异性细菌相关性小;作为AMF的特征脂肪酸16:1ω5c,其与真菌、G-及非特异性细菌正相关,与原生动物的相关性小,与放线菌和G+负相关。土壤容重和pH与排序轴1和2呈正相关;土壤含水量、土壤有机碳、土壤全氮及化学计量比与排序轴1和2呈负相关;土壤全磷与排序轴1呈正相关,与排序轴2呈负相关。海拔的变化引起土壤环境因子的变化,在土壤微生物群落组成中,真菌、非特异性细菌及16:1ω5c受土壤环境因子的影响较大,海拔的变化主要影响了真菌、非特异性细菌、16:1ω5c、G+的相对丰度,其与真菌、非特异性细菌、16:1ω5c、G-及18:1ω9c呈正相关,与原生动物的相关性较小,与放线菌和G+呈负相关。
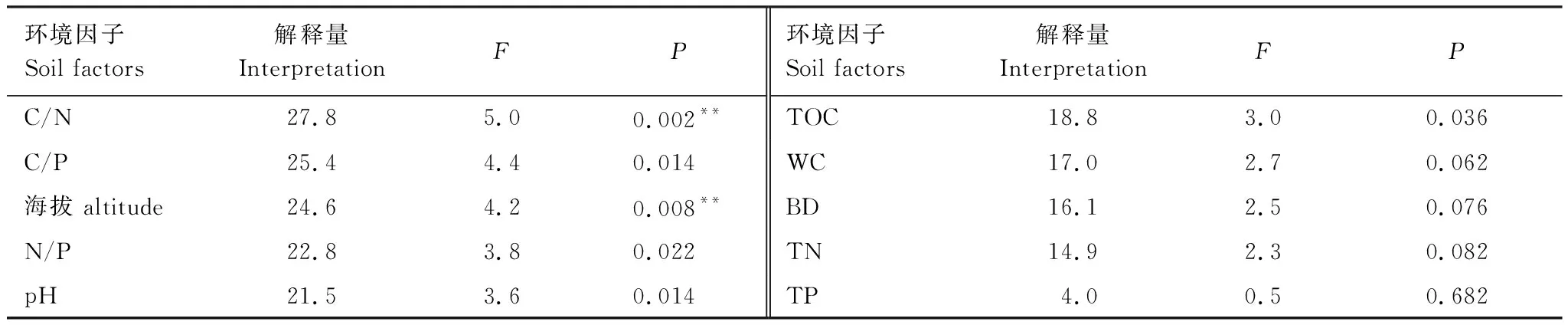
表2 环境因子对土壤微生物的解释量和显著性检验结果
3 讨论
森林生态系统中,土壤微生物与植物关系缜密,植物的根系分泌物及形成的凋落物会直接或间接地影响土壤微生物,而微生物参与土壤养分循环从而影响植物的生长[29]。海拔的变化会在较短时间内影响气候环境因子的改变,土壤微生物作为土壤生态系统中的敏感指标会对环境的改变产生剧烈的响应[30]。研究贺兰山不同海拔下土壤微生物含量可以看出,细菌含量最高,接下来是放线菌,其次是真菌,原生动物的含量最低;同时海拔2139m处微生物的含量高于其他海拔,土壤微生物的含量整体呈先增加后减小的趋势,这与谷晓楠等[31]研究长白山土壤微生物群落结构发现随着海拔的升高土壤微生物含量呈现先增加后减少的变化趋势相同。与吴则焰等[32]研究武夷山不同海拔下土壤微生物随着海拔的升高微生物的种类和含量逐渐下降的结果不同;孟苗婧等[33]研究发现在凤阳山针阔混交林地土壤微生物含量在不同海拔有所不同,由此可见在不同的研究区域土壤微生物含量随海拔变化有所不同。有研究发现较高的植物群落多样性伴随有较高的土壤微生物多样性[34]。贺兰山海拔2139—2438m处分布大部分阔叶树种,其丰富的凋落物可以为微生物提供大量的营养物质,这是该海拔土壤微生物含量较大的原因之一。
真菌和细菌是土壤中两种重要的微生物种类,真菌/细菌可以表征两个种群相对丰度的变化状况,比值越高说明生态系统越稳定,研究发现真菌和细菌在海拔2139m处的含量高于其他海拔,这与刘秉儒等[17]在贺兰山研究发现海拔1900—2100m处真菌含量最高的结果一致,说明该海拔的温度、水分、有机质等条件更适合真菌生长。G+和G-的含量随海拔的变化与其他微生物含量变化类似,研究发现G+/G-在海拔2139m和2249m处最小,而G+/G-可以指示土壤营养状况,比值越大则营养胁迫越强烈[31],可见在海拔2139m处土壤肥力较好,这与该海拔下丰富的物种多样性密切相关,所产生的凋落物为微生物提供高营养、易分解的底物,促进了细菌的繁殖。8种微生物指标的相对丰度分析结果表明,不同海拔梯度土壤微生物的相对丰度存在差异,在海拔2139m处相对丰度最大,由于该海拔有着较高的生物多样性,这也符合山地生态学中的“中部膨胀”理论,在山地低海拔地区受水分的限制,高海拔地区受热量限制,而中部区域生产力大,物种多样性高[35]。革兰氏阳性细菌在海拔1380m处丰度最大,革兰氏阴性细菌在海拔2249m处丰度最大,这说明革兰氏阴性细菌更适合高海拔环境生存[36]。放线菌一定程度上会促进土壤有机物的分解,改善土壤结构,同时放线菌在农业、工业和医药等领域有重要价值,与人类生活密切相关。研究发现放线菌丰度最大值出现在海拔1380m处,这与朱文杰等[37]对太白山北坡土壤放线菌垂直分布的研究发现在海拔800—1845m范围内,土壤放线菌最高含量出现在1845m的结果有所差异,在贺兰山海拔1380m处人类的生产和活动可能是造成这种现象的原因之一。
山地森林生态系统中,海拔的变化造成物种丰富度的不同及土壤性质的差异,对微生物群落产生影响[38]。RDA分析结果表明,贺兰山海拔和C/N对土壤微生物影响显著。有研究认为土壤养分含量是影响土壤微生物群落的重要因素,土壤的N、P含量一定程度上影响土壤细菌群落多样性[39];有研究表明土壤的物理性质对土壤微生物群落结构作用显著,良好的物理性质一定程度上增加了土壤微生物丰富度[40];也有研究发现温度是影响不同海拔土壤微生物群落结构差异的主要因素[41]。由此可见,海拔引起环境因子的改变不同程度的影响着土壤微生物,土壤微生物的群落结构及多样性乃是多因素的综合结果。
4 结论
土壤有机碳和土壤全氮随海拔的升高而增加,土壤全磷随海拔变化不明显;土壤含水量随海拔的升高而增加,土壤容重随海拔升高而降低,土壤pH随海拔变化不明显。
海拔是影响微生物分布的重要因素,特征微生物的含量和相对丰度随海拔的升高先升高后降低,在海拔2139m处达到最大值,符合山地生态学中的“中部膨胀”理论。
