樟子松固沙林生长对土壤磷素变化的影响
2022-03-10周凤艳张欣月赵志浩吴叶礼雷泽勇
周凤艳,张欣月,赵志浩,吴叶礼,雷泽勇,2,*
1 辽宁工程技术大学环境科学与工程学院,阜新 123000 2 辽宁工程技术大学退化土地生态治理研究院,阜新 123000
磷作为生态系统重要养分元素之一,除了可以控制森林生态系统的净初级生产力和碳分配外,还可以控制土壤碳的分解速率和封存过程[1—2],其在森林生态系统功能维持过程中发挥着非常重要的作用[3]。与初始来源于大气的碳氮循环不同,生态系统中的磷初始来源于含磷矿物的风化[4]。风化的磷进入土体后,通过植物根系吸收,然后分配到各个功能器官,参与各种生理生化过程[5]。从沙地自然生态系统看,土壤磷的输入主要有含磷矿化合物的风化、大气沉降、凋落物的分解释放[6];而土壤磷的输出主要有风蚀[7]、降雨淋失(包括地表径流和淋溶损失)[8]、植物吸收利用[9]。其中,磷的最初来源物——含磷矿化合物在沙地土壤中含量低,因此经风化进入土体的磷素少[8];而沙地凋落物受气候干旱的影响分解缓慢,释放到土壤中的磷也较少[8]。沙地风蚀强烈,导致表层土壤磷的损失大[7],再加上沙地土壤粗颗粒含量高,固持磷的能力差[10]。因此,沙地土壤中的磷元素含量相对较低,被认为是沙地森林生态系统中林木生长的限制性养分因子。从土壤磷的输入输出来源可以看出,沙地土壤磷含量除了受气候因素影响(风化速率、风蚀、降雨等)外,植物种类[11—13]、土层深度[14—15]、土壤性质(pH、质地、岩性)[10,16—17]、土地利用方式[17—19]等也是重要影响因素。沙地土壤中的磷随着生物量的去除(或收获)、淋溶和侵蚀(风蚀和地表径流)而逐渐耗竭[20],生态系统的发展将日益受到磷限制[21]。因此,研究土壤中磷的转化和有效性是沙地生态系统稳定性维持的重要一步[4]。
樟子松(Pinussylvestrisvar.mongolica)天然分布于中国内蒙古自治区大兴安岭林区的山地和沙地。由于其优良的抗逆性能和速生特性,在20世纪50年代被成功引种到辽宁省章古台地区后,作为优良的固沙造林树种被迅速扩大栽植[22]。目前该树种不仅是辽宁省西北部沙区主要造林树种,也是我国东北、华北、西北地区营建防护林的主要树种[23]。但由于以往有关沙地草地建立樟子松人工林分后土壤磷变化及有效性研究存在一定的局限性[24],导致樟子松林分的生长对沙地土壤磷的变化及其影响机理还不清晰,限制了人们对沙地营造大面积樟子松固沙林合理经营和管理的全面认识。基于上述缘由,本文以辽宁省章古台地区营建的不同生长阶段樟子松固沙林为研究样地,以各生长阶段林分附近天然草地为对照,研究沙地樟子松人工林的营建及林分生长对土壤磷的转化及磷的有效性的影响,为我国樟子松人工固沙林地土壤质量的改善、樟子松人工固沙林的合理经营管理以及实现沙区林业可持续发展提供理论依据。
1 研究区与研究方法
1.1 研究区概况
研究区位于原辽宁省固沙造林研究所(现辽宁省沙地治理与利用研究所)章古台试验林场(42°39′—42°43′N,122°23′—122°33′E)(图1),海拔高度为(225±5) m。该地区位于中温带,属大陆性季风气候,年均气温4.6—6.3℃,年平均降水量约500 mm,且降雨大部分集中于6—8月份,年平均蒸发量约1600 mm,土壤主要类型为生草风沙土,pH值约6.7,代表性植物有胡枝子(Lespedezadaurica)、蒺藜梗(Agriophyllumsquarrosum)、蕨麻委陵菜(Potentillaanserina)和中华隐子草(Cleistogeneschinensis)等[25]。该地区樟子松人工固沙林分建立前为固定半固定沙地。
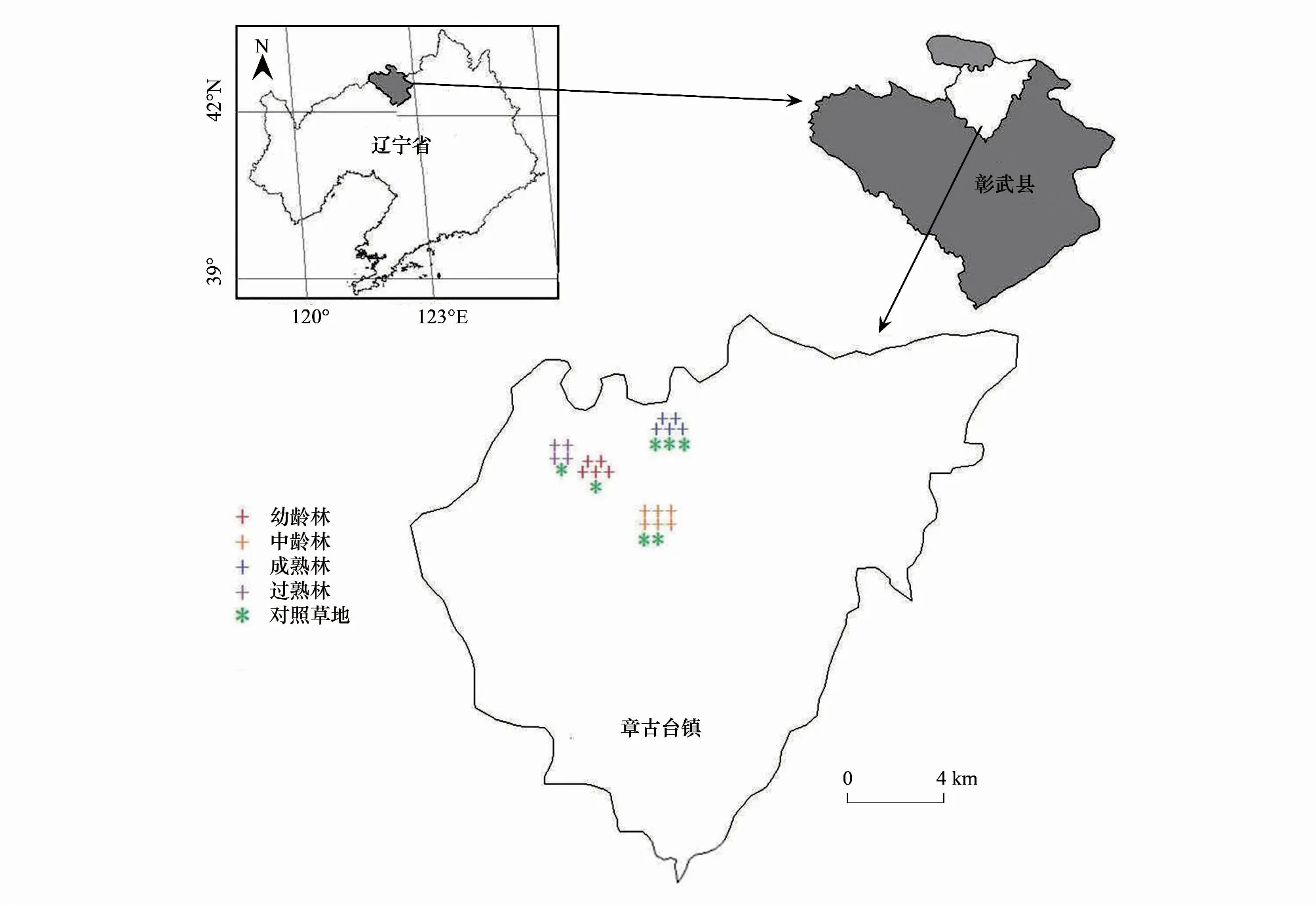
图1 研究区地理位置图Fig.1 Location map of the study area
1.2 试验地选择与土壤样品的采集
2017年8月,通过踏查,基于以下条件(i)标准地林分要覆盖不同生长阶段;(ii)标准地间的距离大于50 m,排除混交林及紧邻道边、农田的林分;(iii)各生长阶段标准地附近需有一定面积的天然草地(作为对照样地);(iv)标准地尽可能平坦,选取了20块樟子松人工纯林作为标准地进行研究,每块标准地的面积为20 m×20 m,其中13年生的幼龄林、27年生的中龄林、42年生的成熟林和56年生的过熟林分别为5、5、6、4块。除了56年生的过熟林造林时的株行距没有查到外,其他各林龄造林时的株行距见表1,各林分经自然死亡或轻微的人为干扰后形成现有林分密度。由于农作物的种植,大面积的天然草地选择比较困难,每个林龄只能就近选择1块天然草地作为对照。2017年8月开始,对标准地林分进行每木检尺,依每木检尺数据计算林分平均胸径、平均高、平均冠幅(表1),依据林分平均高选择标准地的标准木。
标准木选定后,在各标准地内距标准木1 m处分别挖长、宽、深各100 cm的土壤剖面,用环刀(容积200 cm3)分别收集0—10、10—20、20—40、40—60、60—80、80—100 cm 6个层次的原状土,用于测定土壤容重和土壤水分,每层3个重复。同时,在每块标准地内随机选择5个样点收集0—10、10—20、20—40、40—60、60—80、80—100 cm层的土壤样品,同层次的5个样品充分混匀,除去其中的植物根系和石砾,用四分法取500 g样品带回实验室,室温风干处理后碾碎过筛,用于测定土壤化学性质。
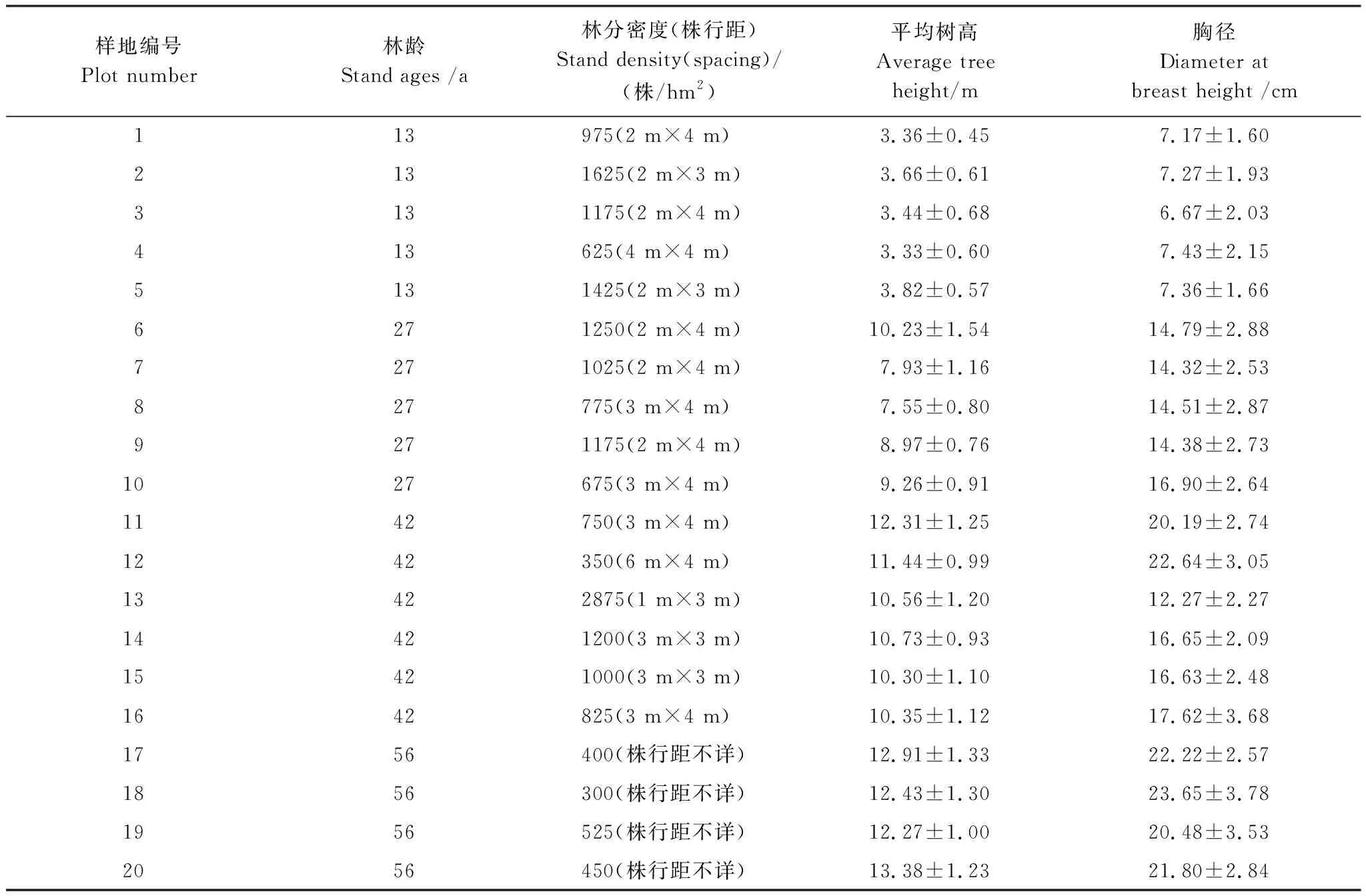
表1 樟子松人工林样地基本情况表
1.3 土壤理化因子测定方法
土壤各理化因子值测定方法如下:土壤全磷含量采用NaOH熔融—钼锑抗比色法,土壤速效磷含量采用NaHCO3法,土壤全氮含量采用半微量开氏法,土壤碱解氮含量采用碱解扩散法,土壤全钾含量采用NaOH熔融—火焰光度法,土壤有效钾含量采用NH4OAc浸提—火焰光度法,土壤有机碳含量采用重铬酸钾容量法—外加热法,土壤含水率采用烘干法,土壤pH值采用电位法,土壤容重采用环刀法,各测定方法及计算详见土壤农业化学分析[26]。土壤机械组成采用激光粒度分析仪测定[18]。
1.4 数据处理与分析
将计算得到的数据按下式计算土壤各理化因子变化量:
ΔXi=Xi-X0
式中,ΔXi为草地造林后各土壤理化因子i的变化量;Xi为樟子松林地各土壤理化因子i的测定值;X0为相应对照草地对应的土壤理化因子i的测定值。
按下式计算土壤理化因子相对变化率:
式中,ΔXi为林分建立后各土壤理化因子i的变化量;X0为相应对照草地对应的土壤理化因子i的测定值;Pi为林分建立后各土壤理化因子i的相对变化率。
以土壤磷(土壤全磷、速效磷)相对变化率为因变量,以其他土壤因子相对变化率为自变量进行逐步回归分析,同时排除各影响因子间共线性关系,得出土壤磷相对变化率回归方程为:
PPi=β0+βiPi
式中,PPi为林分建立后土壤磷的相对变化率;β0为常数;Pi为林分建立后各土壤理化因子i的相对变化率;βi为林分建立后的土壤理化因子i的回归系数。
那么,土壤磷则可以表示为:
式中,Ni为林地土壤磷含量值;N0为相应对照草地土壤磷的测定值;
该式表明土壤磷相对变化率与其影响因子相对变化率的正负效应和土壤各养分因子与其相应的因子正负效应相同,只是系数大小存在差异。
1.5 统计分析
运用SPSS 20.0软件对上述计算的数据进行统计分析,运用最小二乘法(LSD)进行多重比较分析,判断不同林分生长阶段土壤磷垂直分布的差异性,显著水平为α=0.05。利用逐步回归分析建立土壤磷变化率的回归方程式模型。以土壤磷相对变化率为因变量,以土壤有机碳、土壤全氮、土壤碱解氮、土壤全钾、土壤速效钾、土壤含水率、粒径<0.05 mm的土壤细颗粒、粒径0.05—0.25 mm的土壤粗颗粒、土壤容重、土壤pH值的相对变化率为自变量,运用逐步回归分析对自变量进行删选,满足水平P<0.05的变量进入模型。
2 结果与分析
2.1 樟子松固沙林土壤磷素变化
由表2可以看出,樟子松固沙林地的土壤全磷含量在(92.36±4.39) mg/kg—(149.27±14.61) mg/kg之间。土壤全磷含量随林分年龄的增加逐渐递增,至成熟林时达到最高,过熟林地降低,但仍接近于人工幼林地。这一结果表明在幼林至成熟林时期,人工林的生长显著促进了土壤中全磷的积累,而过熟林的生长加速了土壤中全磷含量的下降。另外,林分不论处于哪个生长阶段,各土层间的全磷含量没有显著差异,表明土壤全磷对土层深度不敏感。
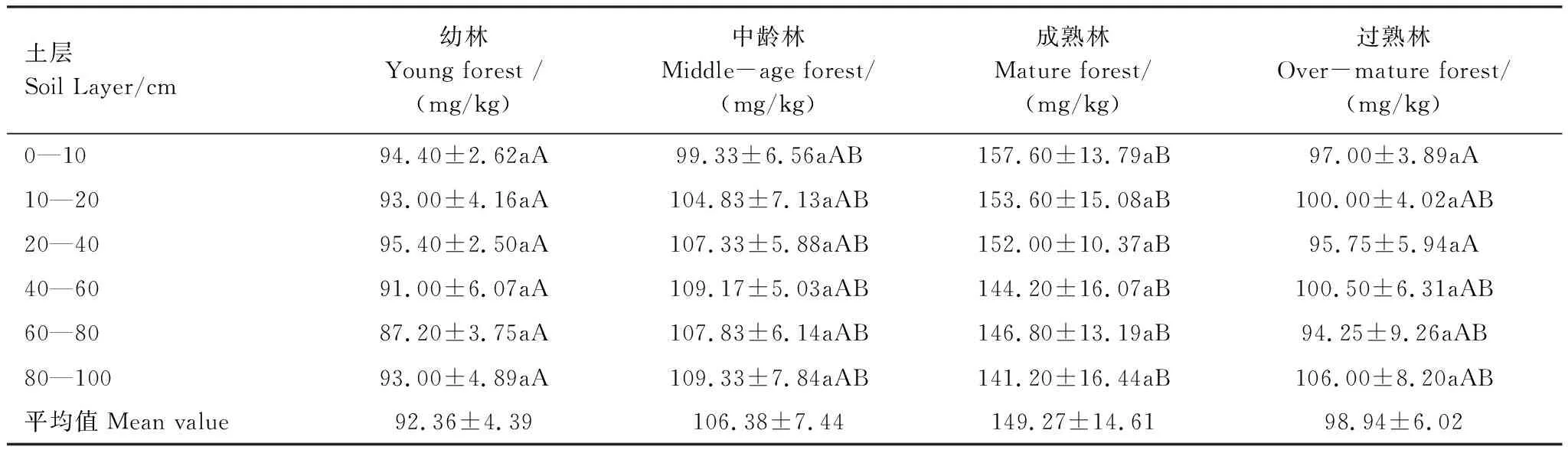
表2 不同林龄的樟子松固沙林地土壤全磷值
表3结果表明,草地营造樟子松固沙林后,总体上看,成熟林地的土壤速效磷含量略高于幼林和中龄林,过熟林最低,但与幼林、中龄林没有显著差异。成熟林地10—40 cm层的土壤速效磷含量显著高于过熟林。速效磷含量从表层到深层有逐渐降低的趋势,但幼林和成熟林地,这种降低的趋势不显著。
2.2 林龄对土壤磷素变化的影响
为量化林分年龄对土壤磷的影响大小,将林分年龄与土壤全磷和速效磷变化量(即相对于草地的增加或减少)分别进行回归分析并建立回归方程。结果表明,不同土层的全磷变化量(增加或减少)均与林分年龄存在显著相关关系。0—80 cm层的土壤全磷量均随林龄的增加而增加,年变化速率值分别是0.0015、0.0035、0.0023、0.0034、0.0028 g/kg;80—100 cm层随林分年龄的增加呈先升高后降低的趋势(图2)。从R2来看,林分年龄对0—10 cm层土壤全磷变化的解释率低于10—80 cm层,表明表层的全磷变化受到更多因素的影响,深层受林分生长影响更大。沙地造林初期,各土层的全磷含量均降低,且随林分的生长逐渐恢复,各土层分别在26、26、13、25、21a恢复到草地水平;在林分成熟期阶段,全磷增加量达到最大(图2)。土壤速效磷的变化在10—20 cm层与林分年龄相关不显著,其余土层的变化均与林分年龄存在极显著负线性相关,年变化率值分别是 -0.0197、-0.0234、-0.0267、-0.011、-0.0225mg/kg(图3)。建立的回归方程表明,除60—80 cm土层外,其他土层土壤速效磷含量自上而下在55、52、43、49年开始低于草地水平。这一结果表明林分的生长加速了土壤中速效磷的减少,和草地相比,林分的生长能够消耗更多的速效磷,樟子松成过熟林的生长将会受到土壤有效磷的限制。
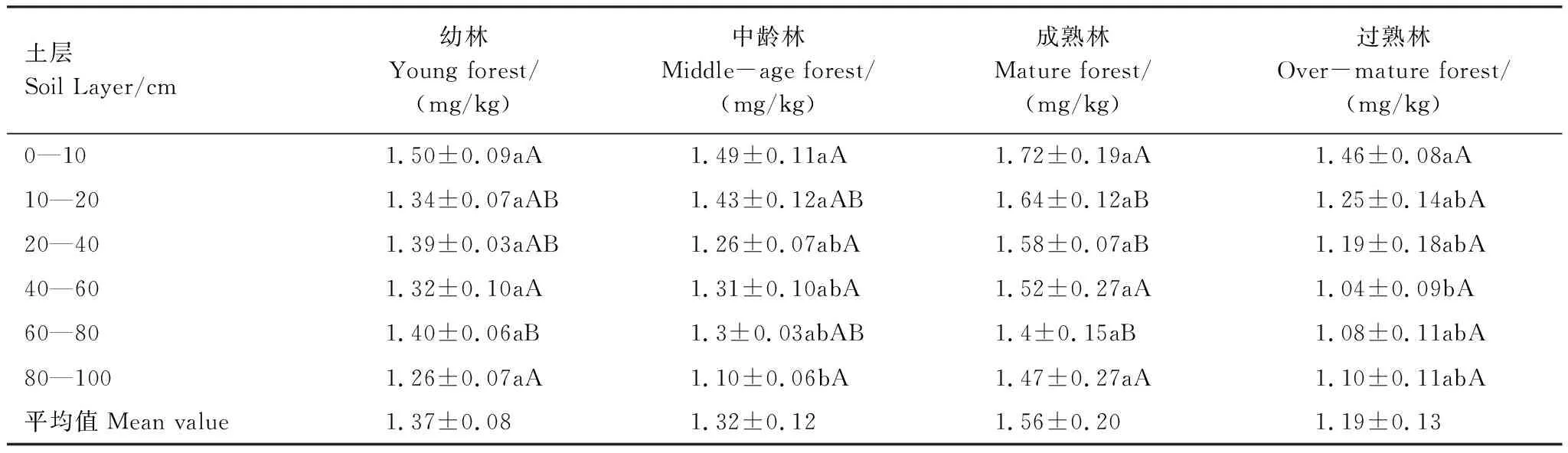
表3 不同林龄的樟子松固沙林地土壤速效磷值
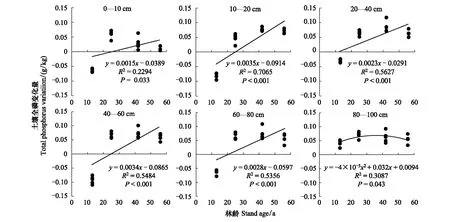
图2 不同土层土壤全磷变化量与林龄的关系Fig.2 Relationship between soil total phosphorus variation and stand ages in different soil layers
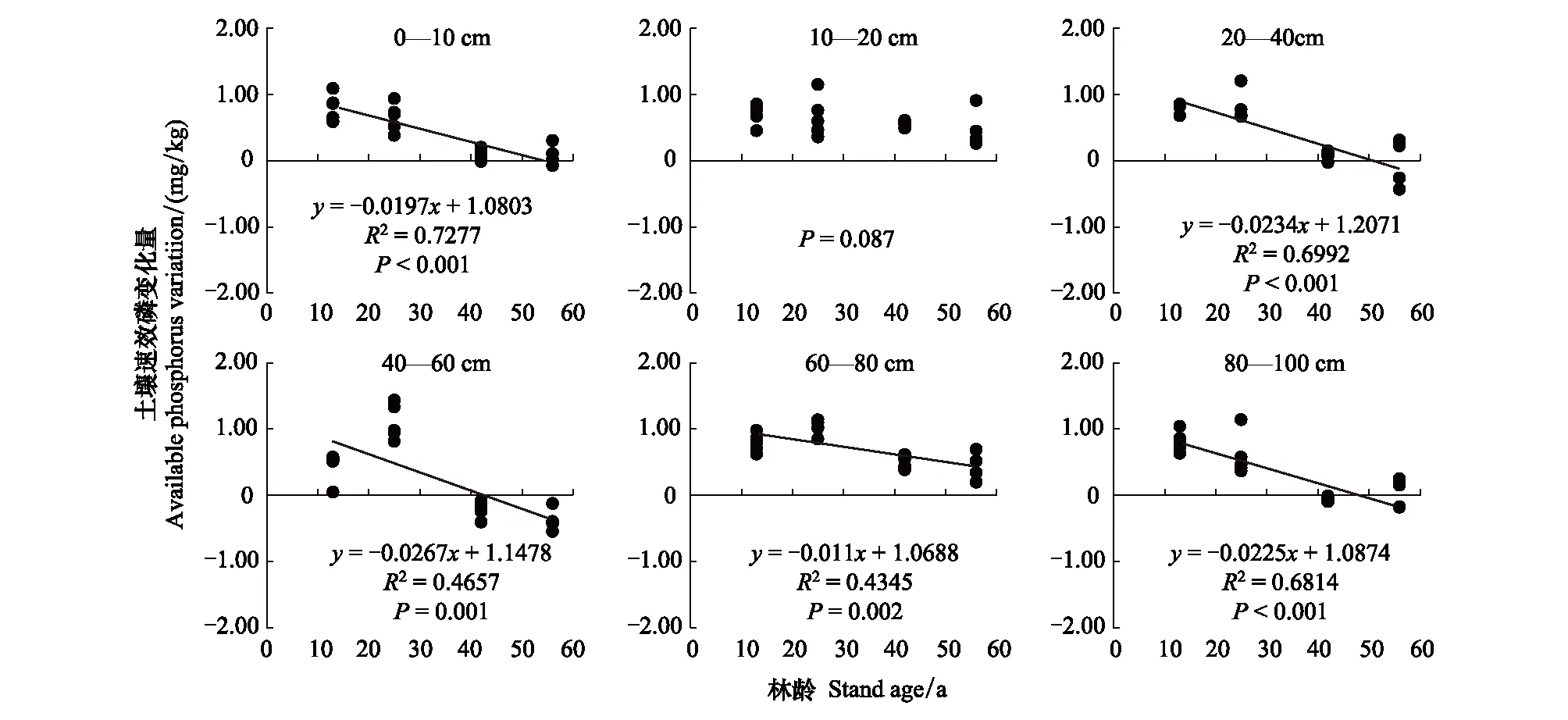
图3 不同土层土壤速效磷变化量与林龄的关系Fig.3 Relationship between soil available phosphorus variation and stand ages in different soil layers
2.3 影响樟子松林地土壤磷素变化的其他土壤理化因子
土壤全磷变化量在不同土层中与其他土壤理化因子关系不一致(表4)。土壤全磷在0—10 cm层与土壤容重和土壤全氮分别呈极显著和显著的负相关关系,与土壤含水率呈极显著的正相关关系,且容重对土壤全磷含量变化的影响大于土壤含水率和全氮的影响;10—20 cm层的土壤全磷与有效氮含量呈极显著负相关,与土壤有机碳和土壤速效钾含量分别呈极显著和显著正相关;20—40 cm层与速效氮和速效磷含量为极显著负相关关系;40—60 cm层与速效氮和速效磷含量分别呈极显著的负相关和正相关关系,与土壤容重和土壤含水率呈显著负相关;60—80 cm层与速效氮、土壤容重呈极显著负相关,与粒径0.05—0.25 mm的土壤粗颗粒含量呈极显著正相关;80—100 cm层仅仅与土壤速效钾含量呈极显著负相关。
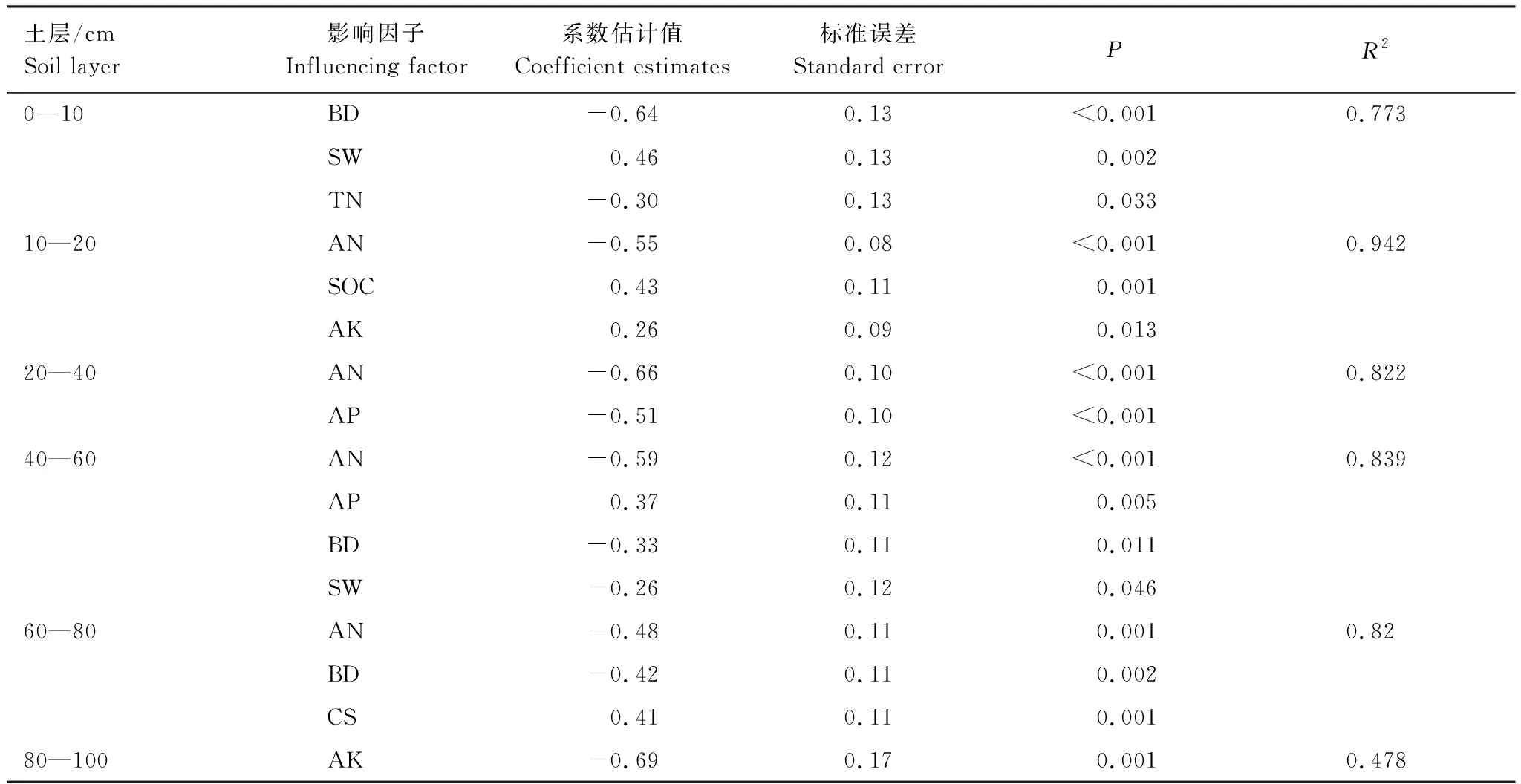
表4 土壤全磷变化率影响因子的回归分析
不同土层影响速效磷含量变化的因子不同(表5)。土壤速效磷含量在0—10 cm层与土壤全氮和土壤有机碳含量呈极显著负相关;在10—20 cm层与土壤有机碳含量和土壤含水率分别呈极显著和显著的负相关,且土壤有机碳对速效磷变化的影响大于土壤含水率;在20—40 cm层与土壤全钾含量和pH值分别存在极显著的正相关和显著的负相关,且全钾含量高低的影响大于pH值;在40—60 cm层与全钾和全磷含量极显著正相关,与粗颗粒含量和全氮含量极显著负相关;在60—80 cm与土壤有机碳含量呈极显著正相关,与土壤全氮含量显著负相关;在80—100 cm层与土壤有机碳含量和pH值分别存在极显著和显著负相关,与土壤含水率呈极显著正相关关系。

表5 土壤速效磷变化率影响因子的回归分析
3 讨论
3.1 樟子松固沙林林分生长对土壤磷素变化的影响
草地营造樟子松固沙林后,任何生长阶段的土壤全磷含量对土层深度均不敏感,这一结果与淑敏等[24]、董雪等[25]的研究结果相似。另外,不同生长阶段樟子松固沙林地土壤全磷含量均低于全国平均水平 (0.65 g/kg)[27],更低于中国温带草原的平均值((0.75±0.54)g/kg)[28],表明沙地土壤中的磷素含量较为贫瘠。
本文结果表明,幼林至成熟林时期,林分的生长显著促进了土壤全磷的积累,而过熟林时期,林分的生长加速了土壤中全磷的减少,即林分的生长促进了土壤全磷含量的变化。研究认为,草地建立人工林分后,林分年龄是引起土壤全磷变化的主要因子[29—30]。如牛沙沙等[31]结果表明,樟子松固沙林地土壤全磷随林分年龄增加呈增长的趋势,Chen等[32]对不同林龄华北落叶松磷的研究也得到了相似结论。樟子松固沙林营建及生长初期,土壤全磷输出增加,输入减少,导致了土壤中全磷的降低。土壤全磷输出增加体现在以下两方面:一方面,由于整地导致地表裸露,提高了土壤温度,增强了土壤微生物活性,促进了土壤有机质的分解和土壤有机磷的矿化,矿化的无机物满足樟子松生长和微生物固定[6];另一方面,裸露的地表,加速了风蚀和季节性的降雨淋失,风蚀和降雨淋失导致土壤表层粘粒含量降低,沙粒含量增加,而粘粒中含有的磷素远高于沙粒[10],同时沙粒中磷的侵蚀损失又大于粘粉粒[33]。土壤全磷输入减少主要在于,林分生长初期,地上地下的凋落物少,通过凋落物分解输入土壤的磷素极少。随着林分的生长,林下植物多样性逐渐增加,表层覆被物亦随之增加,群落结构逐渐复杂,林地凋落物逐渐增多。凋落物的分解引起土壤养分逐渐积累,作为养分元素之一的土壤全磷含量逐渐积累(表2)。同时,覆被物和凋落物增加,还聚集了粘粉粒,提高了表层土壤粘粉粒含量[18—19];和草本植物相比,樟子松的根系在广度和深度两个方面都极大地增加[34],而根及其根系分泌物是土壤剖面有机质的主要来源[35]。因此,根系的增加提高了土壤有机质含量,降低了土壤容重,土壤变得更疏松,增强了土壤固持养分的能力,增加了土壤全磷的含量[19](表4)。而樟子松生长需要的土壤速效磷,大部分依靠根系分泌的磷酸酶、有机酸等物质来促进土壤中有机磷的矿化以及无机态磷酸盐的溶解获得。土壤磷酸酶的活性随林分年龄增加而增加,在30林龄时活性达到最高[36],磷酸酶活性与有机磷矿化速率正相关[37],因此,林分的生长导致土壤中速效磷含量的升高(图3)和全磷含量的降低(图2),促进了凋落物的分解。过熟林时期,林分逐渐衰退[38],地上凋落物逐渐减少,土壤中活的林木根系及根系分泌物也逐渐减少,微生物和酶的活性降低[36],有机质(凋落物和根系及根系分泌物)分解变慢,土壤中全磷的输入逐渐降低,从而导致过熟林地土壤全磷含量明显低于成熟林(表2)。
3.2 樟子松林分生长过程中其他土壤理化因子对土壤磷素含量的影响
其他土壤理化因子对土壤磷素含量的影响也是由林分生长引起的。有机质的分解是土壤养分的主要来源[39]。研究表明,土壤全磷与土壤有机碳含量显著正相关[40],本文的土壤有机碳与土壤全磷的关系在10—20 cm层与这一结论一致。造林初期,由于地表裸露,土壤吸收太阳能辐射增加,地表温度升高,微生物活性增强[6],导致土壤有机质加速分解和有机磷的矿化。分解和矿化释放的磷素一部分保留在土壤中供植物和微生物吸收固定[41],小部分在季节性降雨作用下淋失到深层[42],导致表层土壤全磷含量降低(图2),速效磷含量增加(图3)。随着林分的生长,林下植被及林木凋落物、根系分泌物增多,土壤各层的有机质含量均增加,尽管土壤微生物活性增强,但土壤有机质的补充超过分解,土壤有机碳含量增加[43],导致土壤全磷含量增加(表4)。过熟林阶段,深层土壤呈弱碱性[42],淋溶到深层的速效磷一部分转化为钙磷沉淀下来,一部分保留在土壤中,增加土壤中速效磷含量,因而在60—80 cm层,土壤有机碳与土壤速效磷含量均增加。不仅有机质的分解增加了土壤中的氮含量,大气的氮沉降也能促进土壤氮的积累[44—45],土壤氮的积累促进了植物的生长,但加快了磷素的周转[45],使得土壤中的磷素降低,因此,研究中的土壤速效磷与土壤全氮含量在0—10、40—80 cm层负相关(表5)。长此以往,即使地处温带,樟子松固沙林生态系统的维持与发展也将会受到土壤磷的限制。
土壤pH值既显著影响土壤碳、氮、磷含量[14],也影响养分空间分布[46]。研究表明,在樟子松林生长过程中,表层土壤的pH值逐渐降低,深层逐渐升高[42]。表层pH值的降低,促进了土壤中无机磷酸盐的溶解,增加土壤中速效磷的含量。而深层pH值的升高使得表层淋溶下来的速效磷被钙离子固定,形成难溶解的钙磷,导致土壤中的速效磷含量的降低。
土壤容重的升高和降低不仅能反映土壤的持水性能好坏和蓄水能力的强弱,还能反映土壤结构优劣和有机质含量的高低[47]。本研究中土壤全磷在0—10和40—80 cm层与土壤容重负相关。造林初期,由于破坏了土壤结构,造成风蚀增大,0—10 cm层土壤容重增加,土壤碳、氮、磷含量均降低;另外,樟子松个体生长吸收的养分高于草本植物,使得土壤中的养分含量及土壤微生物量明显降低[48],土壤容重升高。随着樟子松人工林的生长,林地地表枯落物积累量逐渐增加。通过枯落物分解、细根脱落和根系分泌物等途径产生的养分归还土壤[45]。养分的增加使得土壤生物和微生物活动加剧,土壤生物化学反应增强,土壤结构得到改善,导致土壤容重降低[19]。因此,土壤中的全磷含量与土壤容重互相影响。
研究表明,土壤含水率能显著促进土壤磷素的迁移转化[49]、植物根系分布、微生物活动和凋落物的归还[50],从而间接影响磷素循环过程。本文的土壤全磷含量在0—10 cm层与土壤含水率正相关证实了这一观点。草地建立林分后,随着林木的生长,大气中细颗粒物质的沉积、林木凋落物及根系分泌物的分解转化,使得土壤细粒含量增加,土壤持水能力增强,土壤容重降低[19]。土壤含水率升高改善了土壤理化性质,增加了土壤养分含量,也增加了土壤中的磷素。与其他土壤性质相比,土壤含水率的影响更复杂,因为水分是动态变化的,土壤含水率与土壤磷素关系的研究今后应继续加强。
4 结论
沙质草地营造樟子松人工林后,林分的生长促进了土壤中全磷含量的变化,从幼林至成熟林,土壤中的全磷逐渐增加,但过熟林期间土壤全磷含量出现了明显降低。除林分生长的影响外,表层的土壤全磷变化还受到地表覆盖情况、土壤容重的影响,同时,土壤中的全磷含量对土层深度不敏感。樟子松的生长对土壤速效磷含量影响不显著,成熟林地土壤中速效磷含量略高于幼林和中龄林,过熟林最低,但与幼林、中龄林没有显著差异。除此以外,草地营造樟子松林后,随着林分的生长,土壤中的全磷含量还受到土壤容重和土壤全氮含量的影响,土壤中的速效磷含量还受到土壤有机碳含量和pH值变化的影响。
