鱼类对海洋升温与酸化的响应
2022-03-10王晓杰谢金玲袁一鑫
王晓杰,谢金玲,袁一鑫
1上海海洋大学 海洋生物系统与神经科学研究所,上海 201306 2上海海洋大学 国家海洋生物科学国际联合研究中心,上海 201306 3上海海洋大学 水产种质资源发掘与利用教育部重点实验室,上海 201306
自工业革命以来,由于人类燃烧化石燃料,向大气释放了大量CO2,导致地球表面平均气温升高大约 0.7℃,并且以此 CO2排放速率计算,至 21 世纪末地表温度将升高 2—4℃[1]。全球平均气温上升了约1℃已经改变了物种的地理分布,海洋变温动物通过向两极扩散来寻找温度适宜的水域[2]。由于海-气CO2交换,人为排放的CO2总量的四分之一进入海洋,引起海水化学改变,包括海水中溶解CO2的量增加,海水pH从工业革命前的8.2,至2100年将会降低到7.8,以及降低生物组织中碳酸钙的溶解度[3—5]。
海洋鱼类尽管具有较强的酸碱调节能力,但也可能受到酸化的不利影响[6—13]。并且,温度是鱼类生理和生态的主要非生物决定因子[14—15]。海洋升温也将对海洋鱼类产生广泛影响,包括已有报道的向两极扩散[16—17]和降低低纬度鱼类的生长率[18]。目前,升温和酸化双重环境胁迫因子对海洋鱼类影响的研究也开始引起关注[8, 19]。海水升温和酸化不仅能够直接影响鱼类的生理和行为,也可能通过改变栖息地环境以及改变食物网结构而间接影响鱼类[20]。而鱼类同时也是海洋食物网的中心环节,海洋环境对鱼类资源的影响必将影响全球渔业资源产量[21]。
为此,本文综述了海洋酸化、海洋升温以及两者共同作用对鱼类的影响,为预测鱼类响应全球海洋环境变化的趋势提供相关依据。
1 海洋升温对鱼类个体、种群及生态系统的影响
至21世纪末,全球海洋的平均温度将升高2.6—4.8℃[22],这将对不同区域、不同鱼类造成不同的影响。在赤道及两极地区,海水水温的日变化和季节变化较小[23—25],耐受温度变化范围较窄[26],温度变化一旦超出其适宜范围,海洋鱼类将消耗更多能量用于维持日常代谢,因此会降低其生存适宜度[27]。这些鱼类或者通过生理调节适应升温,或者迁移到其他温度适宜的纬度或水深度,也可能在局部地区彻底消失[16]。温度变化会影响鱼体内生物化学反应速度和代谢速率[28—29],进而影响生长、觅食和繁殖等生命过程中能量供给,这将间接影响鱼类种群分布、群落结构及生态系统功能。
1.1 海洋升温对鱼类生理代谢影响

图1 温度对鱼类有氧范围的影响Fig.1 Influence of temperature on aerobic range of fish
不同种类、不同地理区域的鱼类对温度耐受范围不同。经研究发现,有氧范围是决定鱼类温度耐受能力的重要因素。有氧范围即基础氧气摄取和最大氧气摄取之间的间距(图1)[30]。温度升高对有氧范围的影响是海洋升温影响鱼类存活最重要的生理机制。伴随温度升高,鱼体合成代谢需氧量增加,然而水中溶解氧却在下降,鱼体血液循环和呼吸系统的能力不能满足基础代谢氧气需求量的增加。氧气需求量和氧气供给量之间的不平衡将会限制有氧范围,而有氧范围的减少将会降低鱼类生存适宜度[27, 31]。
热带海洋鱼类生活在水温相对恒定的环境中,他们对于海洋升温更加敏感。当水温升高至比夏季平均水温高 2—4℃时,5种珊瑚礁鱼类基础代谢耗氧率升高,而最大氧气摄取量并不增加,从而造成有氧范围的下降[32]。对于绝大多数海洋鱼类,在发育过程中首先经历浮游生活阶段,然后到近岸适宜栖息地定居,其中外海漂浮生活的仔鱼最容易受到环境改变的影响,死亡率较高[33]。从1998年到2011年,从澳大利亚Lizard岛共采集10批次刚定居的雀鲷(Pomacentrusmoluccensis),通过耳石微结构分析,推算仔鱼漂浮阶段的时间(pelagic larval duration, PLD)和仔鱼的日生长率。研究发现随着温度升高,PLD逐渐下降而生长率不断增加,而当海水温度超过28℃后,温度再升高,PLD趋向增加而生长率却在下降[34]。实验表明,鱼类对水温的耐受能力与水温之间是呈倾斜的圆顶形的关系,在最适温度范围内,鱼类的生长率会随水温升高增加,但是当水温高于最适温度后,其生长又会快速下降[26]。通常变温动物在温度升高时,会降低食物特殊动力系数,减少对食物消化、吸收和同化作用的能量消耗,以此维持日能量收支平衡[35]。但是,对出膜6—9d小丑鱼(Amphiprionpercula)基础代谢、餐后代谢反应的研究,发现升温提高仔鱼基础代谢率,对餐后用于消化和吸收的能量消耗却没有影响,因此在海水温度升高时,仔鱼可能需要寻找更多食物才能维持正常生长速率[36]。而当食物不充足时,升温增加能量消耗,用于生长和发育的能量相应地减少,导致仔鱼生长速度下降,降低变态成功率[37]。并且,在野外长期的观察也发现海洋升温对鱼类早期发育的潜在威胁。在长达58年对加利福尼亚州南部海区43种鱼类幼鱼的物候学研究中发现,在近十几年,39%的种类(主要是外海漂浮性鱼类)季节丰度最高值提前,19%的种类(主要是近岸底栖性鱼类)延后。而浮游动物的物候学并没有随之提前,因此,那些物候学没有与浮游动物同步改变的鱼类将得不到充足的食物,最终减少渔业补充量[38]。
在温度较高的环境中,生物个体趋向变小,因为这样可以增加表面积与体积比,从而容易保持氧气吸收和消耗平衡[39]。在40年间,伴随着冬季水温升高1—2℃,北海8种主要的经济鱼类中的6种,其渐近体型的大小随之下降,从而使这些群体的单位补充渔获量平均降低23%[40]。从全球角度,由于海洋升温使得小个体鱼种可能代替大个体鱼种,据估计到2050年,鱼类集合平均最大体重将会减少14%—24%[41]。
某些海区海面平均温度的升高速度高于全球平均气温的升高速度,等温线在向两极移动[42]。作为物种表型可塑性的一种方式,行为温度调节可以降低环境温度改变对物种和种群的不利影响。鱼类是外温动物,通过迁移活动来维持体内适宜温度,而最适温度理论上反映了鱼类有氧运动能力的最适温度。然而,目前许多热带鱼类所在纬度的最高温度已经接近它们的最适温度[43]。因此,这些种群可能就会暂时性地迁移到更深水层或者高纬度等水温更低的水域,以此降低基础代谢消耗。最近的报道称有80种热带和亚热带的草食性鱼类的生活区域已经扩散到了温带珊瑚礁区[44]。
1.2 海洋升温对鱼类与其他物种间相互关系的影响
在以往研究全球气候变化影响时主要关注全球升温对单一物种直接的生理影响,然而最近通过对陆地和淡水系统的长期数据(>20年)的meta 分析发现,全球升温对鱼类分布区域改变,以及带来新的种间关系的影响,更有可能改变整个生态群落的结构和功能[45]。
当小生境水温不同时,鱼类会通过迁移寻找适宜温度的栖息地,尤其是当食物链顶端的捕食性鱼类对温度比较敏感,它迁移栖息地后,将会改变食物网结构和能量流动。例如,湖红点鲑(Salvelinusnamaycush)是冷温性鱼类(适宜温度10—12℃),对温度敏感,是美国北部湖泊中顶级捕食者,是食物网中关键物种。夏季平均气温从15—20℃变化过程中,因为湖泊中有温度分层,鲑鱼会离开水温较高的近岸,而游向水温较低的深水区,与此相伴,它对来自近岸的小型鱼类和无脊椎动物的捕食量将会下降,而更加依赖深水区的浮游植物的生产力,使得湖泊中食物网结构发生改变[46]。而这种基本食物网结构再造,会通过调节到捕食者的能量通量,改变生物量积累,在环境变化快速时期终将威胁到生态系统的可持续性。
在全球,随着海洋升温和海流的向极化增强,许多热带鱼类入侵至温带海域,由于物种入侵,海洋升温将会间接影响鱼类与其他物种间的关系,进而影响到群落结构和海洋生物多样性。入侵物种主要是珊瑚礁关键草食性鱼类,例如独角鱼(Nasounicornis)、鹦鹉鱼、兔子鱼(Siganusrivulatus)和刺尾鱼属(Acanthurussp.)的一些种类。其中,热带草食性兔子鱼,因为海水升温经苏伊士运河拓殖到地中海,并建立了大量的种群。他们破坏了近岸大型海藻林,并阻止新藻场的建立,深刻改变了近岸岩礁系统[44]。澳大利亚东部地处热带-温带过渡地带,有海藻森林这种关键栖息地。进行长期观测发现,10年间水温升高0.6℃,伴随着入侵的草食性鱼类种类增加以及高温下这些鱼类较高的啃食速率,海藻林逐渐减少,最后消失。同时,当地之前丰富的鱼类群落也逐渐消失,取而代之以热带草食性鱼类占优势的群落[47]。在日本南部部分海区观察到珊瑚已入侵至纬度较高的温带海区,珊瑚-海藻相互作用增强,这些区域正在以海藻为主慢慢转变以珊瑚为主。如果大型海藻场消失而不是被替代,那么物种多样性会急速下降,然而,若被珊瑚替代,那物种多样性可能不受影响,甚至会增加。据估计到2100年,大范围入侵将会导致多区域群落间的物种趋同化,从全球看局部物种丰富度净增加,在入侵物种多的海域将会形成非相似群落[48]。
受全球升温影响,海洋鱼类分布区移向高纬度和深水区。利用捕捞平均气温指数,分析1970—2006年间的年渔业捕捞量,得知海洋渔业捕捞量受气温变化的显著影响,在高纬度地区暖水性物种占捕捞优势种的趋势越来越明显,而在热带海区亚热带物种捕捞量越来越少。这种捕捞组成的改变对沿岸地区渔业经济发展有较大影响[41]。例如,金枪鱼具有重要经济价值,它们依赖于逆流热交换系统维持体温高于环境水温,因而水温是决定其分布的重要因素。热带金枪鱼主要分布于水温高于18℃的水域,然而1967—2011年间延绳钓捕捞量的数据显示,在大西洋、西太平洋和印度洋的亚热带水域中热带金枪鱼的捕获百分比呈上升趋势。由此表明,伴随全球变暖,热带金枪鱼种群在向两极移动[49]。
2 海洋酸化对鱼类早期发育和行为影响
2.1 海洋酸化对鱼类早期发育影响
研究表明海洋鱼类对海洋酸化具有较强耐受性,因为成鱼可以通过鳃和肾脏进行酸碱调节[9, 50],然而幼鱼由于呼吸和离子交换模式与成体不同[51],并且与成鱼相比具有较大的表面积和体积比[52—53],因此幼鱼对海洋酸化比较敏感。酸化会影响一些鱼类的胚胎发育[54]、仔稚鱼的生长[6],破坏仔鱼组织和器官结构[10],降低仔鱼存活率[6]等。然而,也有研究发现,海水酸化并不影响精子和卵子的受精率[55]、受精卵的孵化率[56]、仔鱼的生长和发育[7, 10—11, 56—57]以及游泳能力[58]等。尽管这些研究在海水中CO2分压为700—900 μatm 时(以目前CO2排放速度,到21世纪末海水中CO2分压),对鱼类存活等并未造成直接的影响[59],但是,酸化对鱼类造成的亚致死效应将可能是影响个体甚至种群最大的危险因素[60]。例如,酸化造成鱼类生长缓慢,个体变小等,在自然种群中,这些个体将会增加被敌害捕食的几率、降低获得食物的能力,进而会增加死亡率[61]。
2.2 海洋酸化对鱼类行为的影响及其机制
在以往的研究中高CO2的研究主要从 H+浓度升高带来的酸性效应研究对鱼类影响,大多研究发现鱼类具有高效的酸碱调节方法,认为鱼类可以耐受较高的CO2分压。
直至2009年在对大堡礁鱼类的研究中,发现酸化会影响珊瑚礁鱼类的感觉和行为。Philip L. Munday 首先报道了酸化海水会干扰小丑鱼幼鱼嗅觉对不同气味的辨别能力。在正常水体中(pH 8.15)小丑鱼幼鱼能够识别一系列不同的气味,正确识别这些气味可以帮助他们找到适合成体生活的珊瑚礁栖息地。而在酸化(pH 7.8)处理刚刚产下的受精卵并继续处理至孵化出膜11 d后,小丑鱼幼鱼会被正常情况下避让的气味所吸引,而在pH 7.6的酸化组,小丑鱼幼鱼对任何气味都没有反应[62]。随后,通过越来越多的室内模拟及野外酸化实验发现,酸化还能影响鱼类听觉敏感度[7]、视觉[63]、降低行为侧向化[64]以及影响幼鱼视觉识别捕食者的学习能力[65],表明酸化对鱼类的整个神经系统都可能产生影响。
海洋酸化对鱼类种群影响取决于该物种的适应潜能,而后者又取决于种群内个体间的遗传变异度。研究发现,酸化对鱼类种群内不同个体的行为影响程度不同[66—67],而且这种个体差异能够跨代传递。将亲代棘鲷(Acanthochromispolyacanthus)根据嗅觉行为分为CO2敏感个体和CO2耐受个体,然后再将它们的后代进行酸化处理。根据脑组织的基因组、转录组和蛋白质组学分析结果发现,高浓度CO2酸化处理后,耐受亲代和敏感亲代的后代的基因和蛋白表达均有差异,这种跨代分子特征表明个体间对CO2敏感度不同将为鱼类种群适应海洋酸化提供可能[68]。
3 海洋酸化和升温对鱼类的复合效应
大气中CO2浓度升高,给海洋带来了双重效应,即海洋升温和海洋酸化[73—75]。然而,目前,同时研究海洋酸化和升温两种胁迫因子对鱼类影响的报道很少,已有报道主要是研究它们对鱼类生理和行为的影响。例如,在2种珊瑚鱼类(Ostorhinchusdoederleini)和(O.cyanosoma)的研究中发现,升温能够降低这两种鱼的有氧范围,这与之前的研究结果一致,更重要的是,酸化会加剧升温的不良影响,使得有氧范围进一步下降[28]。在研究这两种胁迫因子对鳐鱼(Leucorajaerinacea)胚胎发育影响中发现,升温会影响胚胎发育、存活率和代谢率,但酸化会加剧升温的影响,通过增加活动代谢消耗、延长发育时间和降低初孵仔鱼体重面积比等影响胚胎发育[76]。在对冷水性、发育缓慢的南极龙鱼(Gymnodracoacuticeps)的研究中,龙鱼胚胎发育对升温更加敏感,而只有在升温和酸化同时处理时,酸化才会对胚胎发育产生不利影响。升温和酸化协同累加效应,如降低孵化率、影响发育和代谢等会改变生物气候学(如提早孵化),会对于季节分化非常明显的极地生态系统带来不利影响[77]。
海洋酸化和升温还会影响珊瑚礁鱼类的侧向行为和觅食行为等。例如,对照组雀鲷(Pomacentruswardi)有明显的向右转弯的侧向行为,酸化组表现出向左转弯的行为,而酸化升温组侧向性则显著降低[78]。捕食动态是一个关键的生态过程,在对海洋升温和酸化对珊瑚礁群落中鱼类的猎物-捕食者影响的研究中发现,升温和酸化对于总体捕食率的影响是协同累加的,但对于捕食者对猎物选择性的影响则是相互拮抗的[79]。虽然海洋酸化和海洋升温只影响部分海洋鱼类的生理及行为,但可能会通过物种间的竞争、捕食、繁殖、共生和寄生关系,从而对群落和生态系统产生深远影响[80—81]。
4 研究展望
4.1 开展多重环境胁迫因子对鱼类影响的研究
在许多近岸和外海生态系统中,海区低氧和酸化是紧密相关的。在低氧区CO2分压会比预计21世纪末CO2分压高出一个数量级[82]。伴随着全球升温,海洋中低氧且酸化的区域将会进一步扩大[83]。在某些海域,当海水升温、缺氧与酸化三种环境胁迫因子同时存在时,它们共同作用对海洋生物的影响可能比单一因素的影响更为复杂[8, 83]。同时,海水升温、酸化以及缺氧等海水理化性质的改变,也可能改变海洋中污染物的性质和毒性[84]。因此应更加关注多重环境胁迫因子对鱼类的综合影响。
4.2 研究环境变化对鱼类及其生物间相互关系的影响
生物与环境以及不同物种之间复杂的相互作用共同构成了生物群落。竞争、捕食、繁殖、共生、疾病和寄生是种群和群落的关键组织力量[85]。目前,大部分研究是从个体水平研究环境变化对鱼类直接生理影响,然而这些研究不可能提供全面了解鱼类对复杂生态环境的响应。一个种群对环境变化的响应依赖于不同尺度的生态过程,比如行为改变、扩散和种群动态。这些生态过程依赖于栖息地和必需资源的可得性,后者又会受到环境因素的时空变化。因此,应该从目前单一物种的研究方式,逐渐转向研究生物间的相互关系,从更高水平如群落、生态系统水平,了解环境变化对鱼类以及全球渔业的影响。
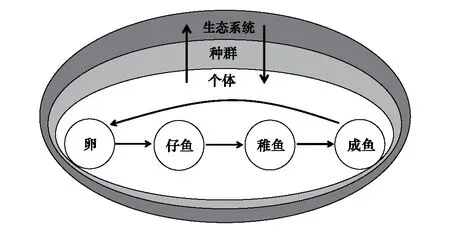
图2 环境变化从不同尺度上对鱼类种群的影响[86]Fig.2 Effects of environmental changes on fish populations at different scales[86]
4.3 鱼类对海洋环境变化的适应性进化
鱼类在从仔鱼、稚鱼到成鱼的不同的生活史阶段,由于其生理特性的不同,如对钙化生物需求量、自身酸碱等生理调节机能,对于海洋酸化和升温等环境变化的生理响应(如行为等)不同(图2)。目前绝大多数研究是通过短期、单世代实验,研究海洋升温、酸化对海洋生物的某一个生活史阶段的影响,而生物对环境变化的长期适应及适应性进化等相关研究则相对较少,因此需要利用数量遗传学和基因组学等方法广泛研究鱼类对环境变化的适应潜能。
