不同山生柳无性系低温胁迫的生理响应及抗寒性比较
2022-03-05王凯月李强峰李玲霞
王凯月,李强峰,李玲霞
(青海大学农牧学院,青海 西宁 810016)
山生柳(SalixoritrephaSchneid.)属杨柳科柳属直立矮小灌木[1],高10~120 cm,是我国特有物种[2]。以山生柳为建群种或优势种构成的灌木林,是青海等高寒地区灌丛的主要类型之一[3]。山生柳灌丛主要分布于我国四川、青海东南部、甘肃东南部、西藏东部[4]。山生柳具有生长迅速、无病虫害等优点,在涵养水源、保持水土、防沙治沙等方面起着重要作用[5]。青海位于中国西部,全省属高原大陆性气候[6],平均海拔3 500 m以上,年均气温-3.7~6.0 ℃[7],在青海地区,低温成为限制植物生长发育的重要环境因素之一。国内外对山生柳的研究较少,主要集中在山生柳的群落分布特征[8-10]、遗传多样性[11]、药理分析及成分分析[12-15]、扦插繁育[16]等方面的研究。Gao等[17]研究发现,山生柳能够广泛生存在高海拔干旱地区。Kang等[18]通过嫁接等无性系选育方法,对山生柳进行繁育,并将这种技术应用于退化草原的恢复。以上研究表明,山生柳在高寒植被群落中具有较高的生态系统功能地位、遗传多样性及药用价值,并在适宜的条件下可进行扦插繁殖。但是针对山生柳优良无性系抗寒性的相关研究相对较少。为了筛选出耐寒性更强的山生柳无性系,更好地发挥山生柳的生态效益,本文对青海不同地区初选出的11个山生柳无性系进行抗寒性差异研究,通过对山生柳无性系抗寒性比较分析得出抗寒性较强的山生柳无性系,为青海高海拔地区城镇植树提供科学依据。
1 材料与方法
1.1 试验材料
从2018年开始在野外进行优树选择确定50个山生柳无性系,通过生长指标的测定,以生长迅速为选育的主要目标,最终确定了11个优良无性系,分别标记为山生柳T10、T15、G25、T11、Q43、T3、G23、T20、T17、G21、G24,所选试验材料的采集分布情况见表1。其中同一林分的不同山生柳无性系是依据优势木对比法,通过测定株高、冠幅、地径、叶长、叶宽、叶面积、一年生新枝生长量、分枝角度等指标,分析各个选优指标变异系数来确定的。
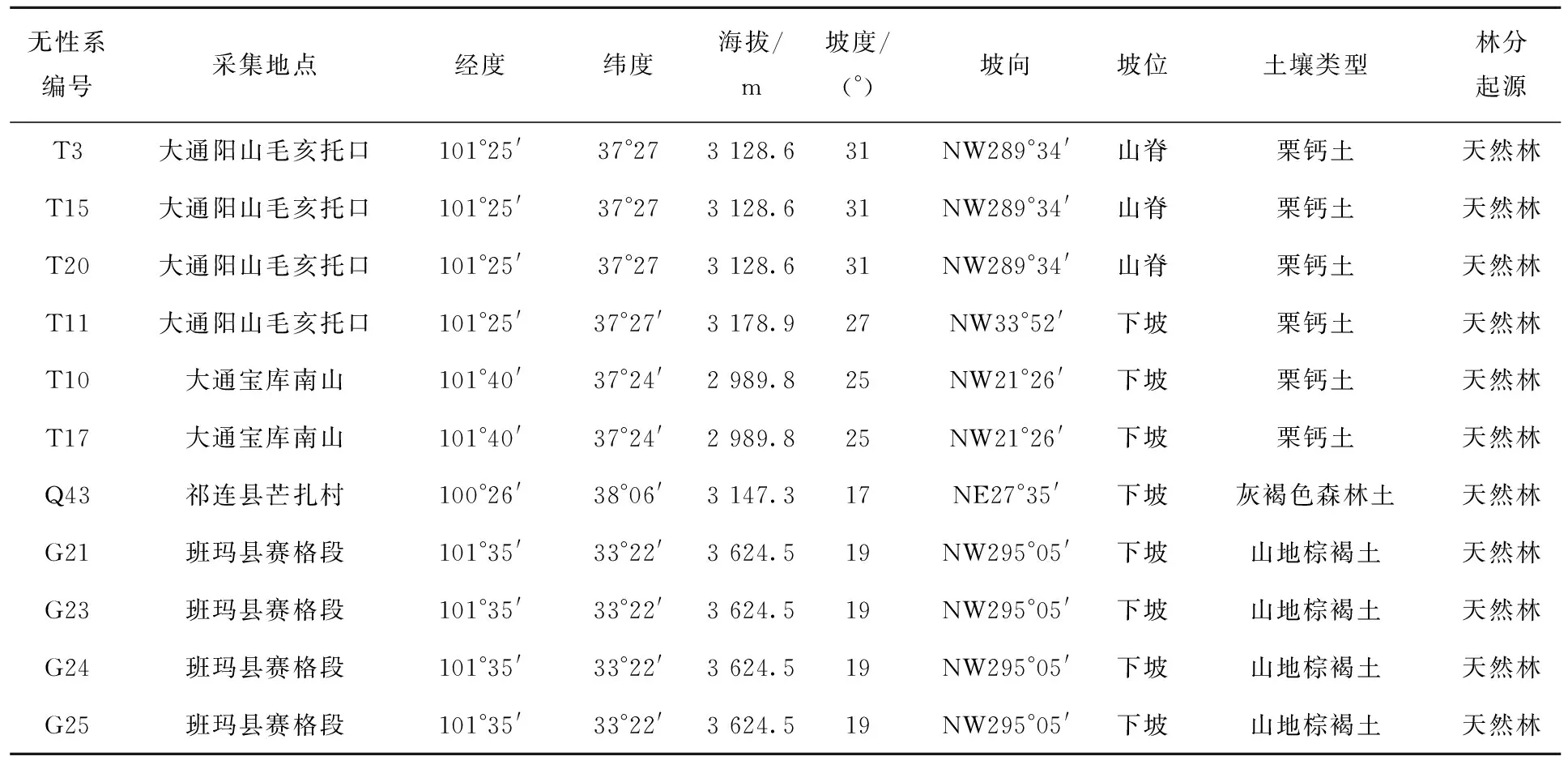
表1 11个山生柳优良无性系林分分布表Tab.1 Distribution of 11 superior Salix oritrepha Schneid.clones
1.2 试验设计
取11个山生柳无性系一年生扦插苗截取插穗,于2020年4月上旬扦插于直径为30 cm、高为30 cm的塑料花盆内,每盆1株,盆中装入8 kg耕作土(田间持水量为30%)。盆栽苗木放置于青海大通植物园温室大棚中,保证11个山生柳无性系苗木正常生长。2020年8月5日进行低温胁迫试验,每个山生柳无性系随机选取3株,通过人工气候箱模拟连续降温过程。设置20(CK)、15、10、4 ℃四个温度梯度,每个温度恒温处理3天后依次降到下一温度梯度,分别测定不同温度处理下植株叶片超氧化物歧化酶(SOD)活性、可溶性蛋白(SP)含量、丙二醛(MDA)含量生理指标,试验重复3次。采用隶属函数法对11个山生柳无性系抗寒适应性进行综合评价。
1.3 测定指标
SOD活性测定采用氮蓝四唑光化还原法[19];可溶性蛋白含量测定采用考马斯亮蓝法[20];MDA含量测定采用硫代巴比妥酸显色法[21]。
1.4 数学模型及数据分析方法
11个山生柳无性系的叶片生理指标数据采用模糊隶属函数法[22]进行综合评判,用隶属度公式对试验测定指标数据进行定量转换,再将各指标隶属函数值取平均进行无性系间相互比较。
隶属函数公式为:
Xij=(xij-xjmin)/(xjmax-xjmin)
(1)
Xij=(xjmax-xij)/(xjmax-xjmin)
(2)
式中:Xij为i树种j指标的隶属函数值,xij为i树种的j性状值,xjmin为各树种j性状的最小值,xjmax为各树种j性状的最大值。当j性状与植物的耐寒性成正相关时用(1)式,成负相关时用(2)式。
通过Excel 2010对所测数据进行整理,运用SPSS23.0软件分析各指标和耐寒性之间的相关性。
2 结果与分析
2.1 11个山生柳无性系叶片SOD活性变化
从图1可以看出,随着胁迫温度的降低,11个山生柳无性系的SOD活性除T10、T20、T3外均表现为先上升后降低再上升的变化趋势。由于试验材料抗寒能力的不同,其SOD活性的变化幅度存在差异。除Q43、G24、T3外,其他山生柳无性系的SOD活性都在4 ℃出现最大值,与20 ℃(CK)相比,增加幅度最大的是山生柳T11,增幅为19.37%。山生柳G25和T3的SOD活性在整个低温胁迫过程中均处于较高水平,说明其抗寒性较强。
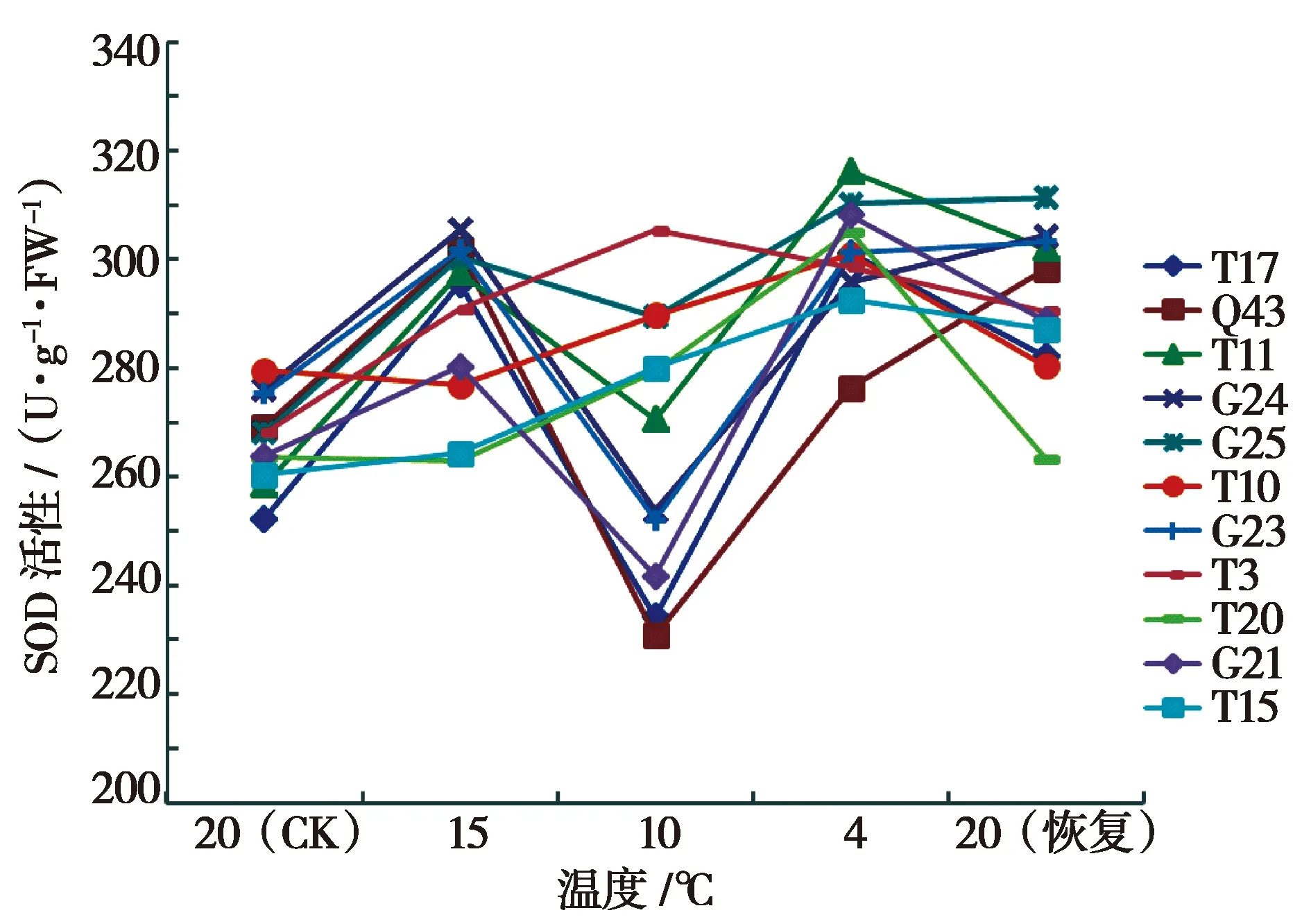
图1 低温处理下11个山生柳无性系SOD活性变化折线图Fig.1 Line diagram of SOD activity changes in 11 clones of Salix oritrepha Schneid treated at low temperature
对11个山生柳无性系在不同温度处理下SOD活性的均值作显著性分析(表2)。低温处理下不同无性系间SOD活性无显著差异。根据SOD活性平均值得出11个山生柳无性系抗寒性强弱:G25>T3>T10>T11>G24>G23>T20>T15>G21>T17>Q43。

表2 低温处理下11个山生柳无性系SOD活性显著性分析表Tab.2 Significance analysis of SOD activity of different clones at low temperature
2.2 11个山生柳无性系叶片可溶性蛋白含量变化
如图2所示,在整个低温胁迫过程中山生柳Q43、G21、G24的可溶性蛋白含量表现为先降低后升高再降低的变化趋势;其他山生柳无性系的可溶性蛋白含量则表现为在20(CK)~10 ℃升高,在10~4 ℃降低的趋势。Q43、G21、G24可溶性蛋白含量在低温处理初期降低的原因可能是开始的低温没有激起植物体内的抗寒性生理调节机制,但又降低了植物体内酶的活性,从而阻碍了蛋白质的合成。山生柳无性系的可溶性蛋白含量在20(CK)~ 15 ℃处理中升高或降低的幅度都比较缓慢,而在15~10 ℃的低温胁迫过程中,所有试验材料的可溶性蛋白含量都出现不同程度的升高,其中T10、T17、Q43增幅最为明显,其他山生柳无性系的可溶性蛋白含量的增幅相对较低。可见,在低温胁迫过程中可溶性蛋白含量的增加对植物抗寒性起到了关键作用。

图2 低温处理下11个山生柳无性系可溶性蛋白含量变化折线图Fig.2 Line chart of soluble protein content variations of 11 Salix oritrepha Schneid.clones under low temperature
对低温处理下11个无性系叶片可溶性蛋白含量的均值进行显著性分析(表3)。山生柳T17与T3、T15、G25的可溶性蛋白含量达到了差异显著水平(P<0.05)。根据可溶性蛋白含量平均值分析得出11个山生柳无性系抗寒性强弱:T17>T10>Q43>T20>G23>T11>G21>G24>T3>T15>G25。

表3 低温处理下11个山生柳无性系可溶性蛋白含量显著性分析表Tab.3 Significance analysis of soluble protein content of different clones under low temperature
2.3 11个山生柳无性系叶片MDA含量变化
如图3所示,随着处理温度的逐渐降低,11个无性系的MDA含量除T10、Q43、G21和G25外,整体变化趋势相似,均表现为先降低后升高的趋势。其中T15、T20、Q43在10 ℃低温下MDA含量达到最小值,T11、T17、T3、G23、G24在15 ℃时MDA含量最低。山生柳T10、Q43和G21的MDA含量在20(CK)~15 ℃的温度处理过程中逐渐升高,在15~10 ℃的温度处理过程中逐渐下降,在10~4 ℃下T10和G21的MDA含量变化趋于平缓。山生柳G25在整个降温处理过程中表现为一直降低的趋势,说明植物体内的抗氧化酶一直处于较高水平,阻止了MDA含量的升高,证明该无性系抗寒能力较强。另外,通过图3可以看出,T15在整个低温胁迫处理过程中MDA含量一直处于较低水平,说明其抗寒能力很强。

图3 低温处理下11个山生柳无性系丙二醛含量变化折线图Fig.3 Line chart of malondialdehyde content variations in 11 Salix oritrepha Schneid.clones under low temperature
由表4得出,山生柳G24、T17与T15叶片中的MDA含量达到了差异极显著水平(P<0.01)。山生柳G23、T3、T20、T10、T11、G25、Q43之间MDA含量差异不显著。由于植物体内的MDA含量与抗寒性呈负相关,MDA含量越高,表明其抗寒性越差;MDA含量越低则抗寒性越强。根据供试材料的MDA含量分析比较可以得出11个山生柳无性系抗寒性强弱:T15>Q43>G25>T11>T10> T20>T3>G23>G21>T17>G2。
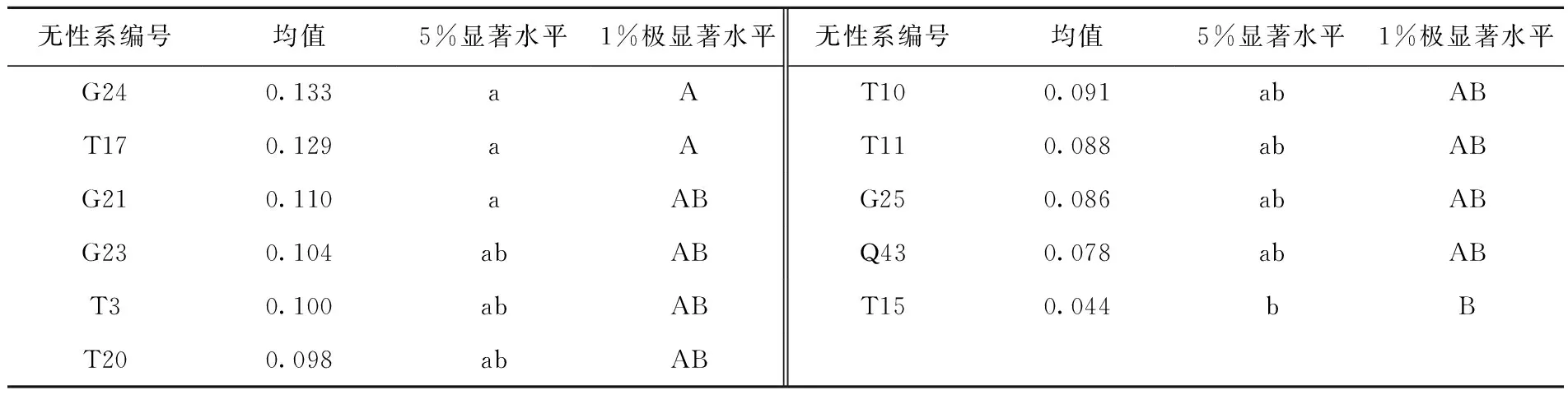
表4 低温处理下11个山生柳无性系MDA显著性分析表Tab.4 Significance analysis of MDA of different clones under low temperature
2.4 11个山生柳无性系抗寒性综合评价
植物在低温胁迫过程中,是多个因子共同作用下引发的一系列结构及生理变化,所以单一指标具有一定的局限性和片面性[23]。因此,采用隶属函数法得出各指标的隶属函数值及各材料的平均隶属度,各无性系耐寒性指标的隶属平均值越大则表明该种无性系抗寒性越强。由表5可知11个山生柳无性系抗寒性排序:T10>T15>G25>T11>Q43>T3>G23>T20>T17>G21>G24。
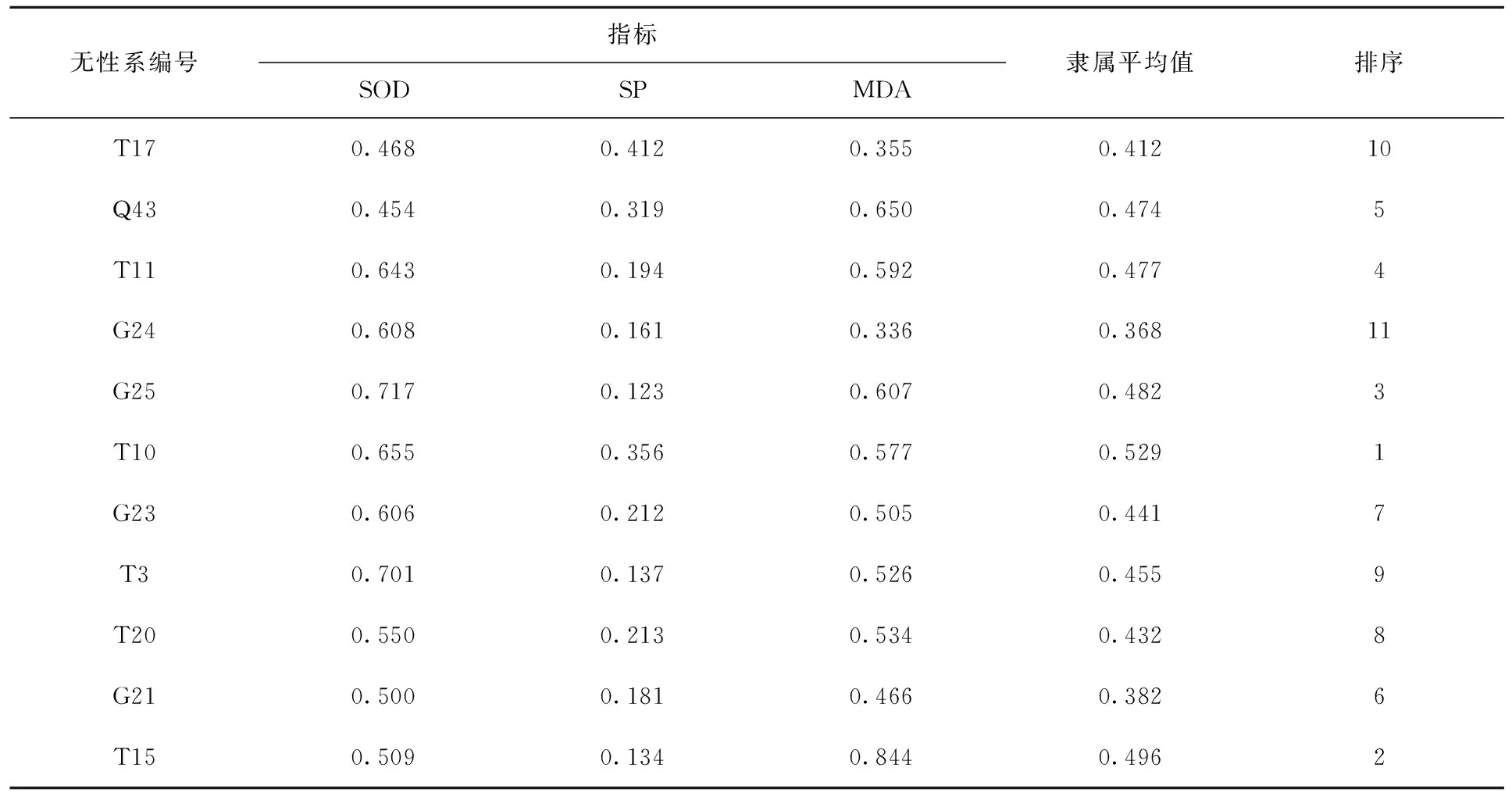
表5 温度处理下11个山生柳无性系抗寒性综合评价Tab.5 Comprehensive evaluation of cold resistance of 11 Salix oritrepha Schneid.clones treated with temperature
2.5 相关性分析
运用SPSS23.0数据分析软件对SOD活性、可溶性蛋白(SP)含量、MDA含量进行相关性分析,见表6。

表6 11个山生柳无性系抗寒性指标相关系数Tab.6 Correlation coefficient of cold resistance index of 11 Salix oritrepha Schneid.clones
由表6可知,MDA含量与其他两个指标呈负相关,11个山生柳无性系抗寒适应力的3个指标间未达到显著水平,说明各个指标之间相对独立。
3 讨论与结论
植物在低温胁迫下会发生一系列生理生化反应,例如代谢异常、生理脱水、光合作用减弱、细胞硬化等[24]。SOD是植物体内重要的保护酶之一,SOD的活性可以间接证明植物抗寒性的强弱,SOD活性的上升证明了低温胁迫下其清除自由基的能力增强[25]。本研究中,在低温胁迫初期,所有山生柳无性系除T10、T20外均表现出升高的趋势,其中G25和T3叶片SOD的活性较高,证明其耐寒性较强。在植物遭受低温胁迫时,可溶性蛋白含量的变化存在两种观点:一种观点认为低温胁迫会导致植物细胞中可溶性蛋白含量增加;另一种观点则认为植物细胞中可溶性蛋白含量的增加与植物抗寒性的增强没有因果关系[26]。在本试验低温处理过程中,11个无性系叶片可溶性蛋白含量的变化分为两种:一种是先升高后降低,另一种表现为先降低后升高再降低的变化趋势。其中T10可溶性蛋白含量与20 ℃对照相比,增幅较明显且耐寒性较强,这与上述第一种观点相符合;此外,11个山生柳无性系在低温胁迫处理后叶片的MDA含量变化规律为除T10、Q43、G21和G25外,其他整体变化趋势相似,均表现为先降低后升高的趋势。该研究结果与王斌[27]对其他植物的研究结果基本一致。在低温胁迫下三个生理指标所得出的山生柳无性系耐寒性具有一定的差异,说明植物的耐寒性是一种复合性状,单用一个抗寒指标很难说明其耐寒性。通过隶属函数法以及生理指标的相关性分析得出,11个山生柳优良无性系的综合抗寒性从高到低依次为山生柳T10、T15、G25、T11、Q43、T3、G23、T20、T17、G21、G24。
植物往往因原产地的不同从而产生内在种质的差异,进而形成植物种质资源抗寒性的不同[28]。林木的地理变异是因为长期在不同的环境条件下,基因交流受到限制,所以在适应生态环境过程中林木种内性状发生遗传分化[29]。本研究11个山生柳无性系中来自高海拔地区的G25、T11、Q43耐寒性比低海拔地区的T3、T20、T17较强,这基本符合Neilsen等[30]发现高海拔种源比低海拔种源及沿海种源更耐寒这一规律,且同一区域的无性系耐寒性指标的隶属函数平均值相差不大。通过无性系叶片生理指标隶属函数综合评价分析得出,大通宝库南山的T10、大通阳山毛亥托口的T15以及班玛县赛格段G25的耐寒性较强,可作为青海省高海拔地区城镇绿化的优选种质材料。
本研究只是对一年生山生柳苗木进行了低温胁迫下部分生理生化指标的测定,并不能完全反映11个山生柳无性系的抗寒性。但本研究所有试验材料均在相同处理、同一条件下测定,结果具有可对比性,可以体现出11个山生柳无性系抗寒性的相对强弱。
