玉米转录因子bZIP G亚家族基因的表达模式
2022-03-03贾利强赵秋芳
贾利强,赵秋芳,陈 曙,丁 波,*
(1.贵州师范学院 生物科学学院,贵州 贵阳 550018;2.中国热带农业科学院 南亚热带作物研究所,广东 湛江 524091)
碱性亮氨酸拉链(bZIP)蛋白家族是众多转录因子家族中最大的家族之一,广泛分布于真核生物中。bZIP转录因子家族都含有典型的bZIP结构域,由60~80个氨基酸残基组成,一般包含N端和C端2部分。N端为含有N-x7-R/K蛋白基序高度保守的碱性区域,功能为识别和结合下游基因启动子区域的DNA序列。C端为2个或2个以上的a-螺旋组成的亮氨酸拉链区域,该区域一般有7个氨基酸残基组成,第7位为亮氨酸(或者被异亮氨酸、缬氨酸或苯丙氨酸替代),可形成二聚体结构调控bZIP蛋白与下游基因DNA的结合。随着植物基因组计划的实施,越来越多的植物基因组序列得以报道,植物众多转录因子家族的研究也开始加速。自2002年Jakoby报道第1个拟南芥bZIP转录因子家族以来,相继在水稻、大豆、玉米、蓖麻、黄瓜、葡萄、苹果、番茄、荠麦、辣椒等植物中发现了bZIP转录因子家族。根据系统进化分析结果,植物的bZIP蛋白可以进一步划分为多个亚家族。拟南芥bZIP转录因子可以划分为10个亚家族,水稻和玉米bZIP家族分别有11个亚家族。众多研究表明,bZIP转录因子家族在植物生长发育与逆境胁迫进程中发挥重要的作用。植物bZIP转录因子家族影响植物生长发育进程,如细胞伸长、组织器官发育、胚形态建成和花器官发育、种子成熟。bZIP转录因子家族作为植物应答逆境胁迫信号途径的中枢位置已在许多研究中体现。它们通过调控下游基因网络途径,影响植物各种生理生化代谢过程,抵御不利的环境因子,如干旱和盐胁迫、高渗胁迫和低温胁迫。bZIP转录因子家族还参与调控激素和糖信号通路、碳氮代谢平衡、病原菌响应和光信号通路。各类植物bZIP家族基因的组织与应答逆境胁迫表达模式研究报道日益增多,这些也加快了植物bZIP家族基因功能研究进程。
玉米作为世界和我国重要的粮经饲三位一体的大宗作物,在世界粮食和饲料安全方面发挥着重要作用。玉米生长中经常遭受各类逆境胁迫因子的影响,造成玉米产量减产,导致种植者经济收入遭受损失。研究表明,转录因子在调控植物生长发育与抗逆性等方面发挥多种作用,其中bZIP家族基因报道的相对较多。玉米自交系郑58是我国重要的骨干自交系,其bZIP家族基因应答逆境胁迫的表达模式还未见报道。本研究选取bZIP家族G类亚家族的20个基因成员,通过人为模拟高盐、干旱、低温和氮缺乏等非生物逆境胁迫条件,系统地研究了bZIP G亚家族成员的表达模式,可为进一步深入研究这些基因的生物学功能提供科学依据。
1 材料与方法
1.1 材料
以玉米自交系郑58为试验材料。在授粉扬花期上午10:00左右,采集成熟根系、茎、第12片叶、雄花和授粉15 d的嫩穗,快速液氮处理,-80 ℃保存备用。
1.2 方法
1.2.1 生物信息学分析
75个拟南芥和20个玉米基因及其蛋白序列来自于Phytozome v9.1数据库(http://www.phytozome.net/)、MaizeGDB(Maize Genetics and Genomics Database)数据库(http://www.maizedb.org/)和TAIR(https://www.arabidopsis.org/)。依据基因的物理距离,利用Mapinspect软件进行染色体定位分析。依据的CDS和基因组序列,利用在线软件GSDS(http://gsds.cbi.pku.edu.cn)进行序列比对,确定的基因结构。75个AtbZIP和20个ZmbZIP蛋白序列组成的共计95个蛋白数据矩阵,利用在线软件MAFFT的L-ISN-i方法进行序列比对分析,去除非保守蛋白区域,保守的bZIP蛋白序列导入MEGA X软件,利用NJ法构建系统进化树,参数设置为-distance,bootstrap值设为1 000,其他为默认值。
1.2.2 逆境胁迫处理
玉米自交系郑58种子经5%次氯酸钠溶液表面消毒后,28 ℃黑暗催芽,挑选出芽一致的健康种子放于网纱上,用Holland营养液培养。营养液配方为:2 mmol·LNHNO,0.17 mmol·LKHPO·12HO,0.27 mmol·LKSO,0.47 mmol·LCaCl·2HO,0.37 mmol·LMgSO·7HO,45 μmol·LFe·EDTA,0.16 μmol·LCuSO·5HO,0.15 μmol·LZnSO·7HO,0.10 μmol·LNaMoO·2HO,15 μmol·LHBO,4.6 μmol·LMnSO·5HO,1 μmol·LNiSO,用HCl调节pH值为5.7~5.8。生长条件设置为长日照(16 h光照,28 ℃;8 h黑暗,25 ℃)。待玉米幼苗生长至三叶期时进行胁迫实验。
干旱或盐胁迫处理:取生长一致的玉米幼苗转移至含有20% PEG6000或200 mmol·LNaCl的营养液中培养;低温处理:将生长一致的健康玉米幼苗置于4 ℃培养。这3种胁迫处理分别在0、1、6、24 h取叶片。铵态氮或硝态氮胁迫处理:取生长一致的玉米幼苗转移至硝态氮或氨态氮缺乏的Holland营养液中培养,分别在0、1、6、24 h取叶片和根系。样品迅速液氮速冻处理,于-80 ℃冰箱保存备用。
1.2.3 RNA提取和实时荧光定量PCR(qRT-PCR)分析
取0.1 g玉米叶片或根系液氮快速研磨,利用RNA提取试剂盒提取总RNA,检测其浓度和纯度后,按照反转录试剂盒说明书完成cDNA的合成和纯化。依据目的基因保守性,利用Oligo 7设计特异性引物,引物信息见表1。以不同胁迫处理的cDNA为模板,以玉米基因为内参,利用Roche Light Cycler 480 System(Roche,Basel,Switzerland)进行定量分析。荧光染料为SYBR Premix EX Taqkit。扩增程序为95 ℃ 3 min;95 ℃ 10 s,58 ℃ 10 s,72 ℃ 25 s,45个循环。采用2法计算基因的相对表达量,每个处理设置3个重复。
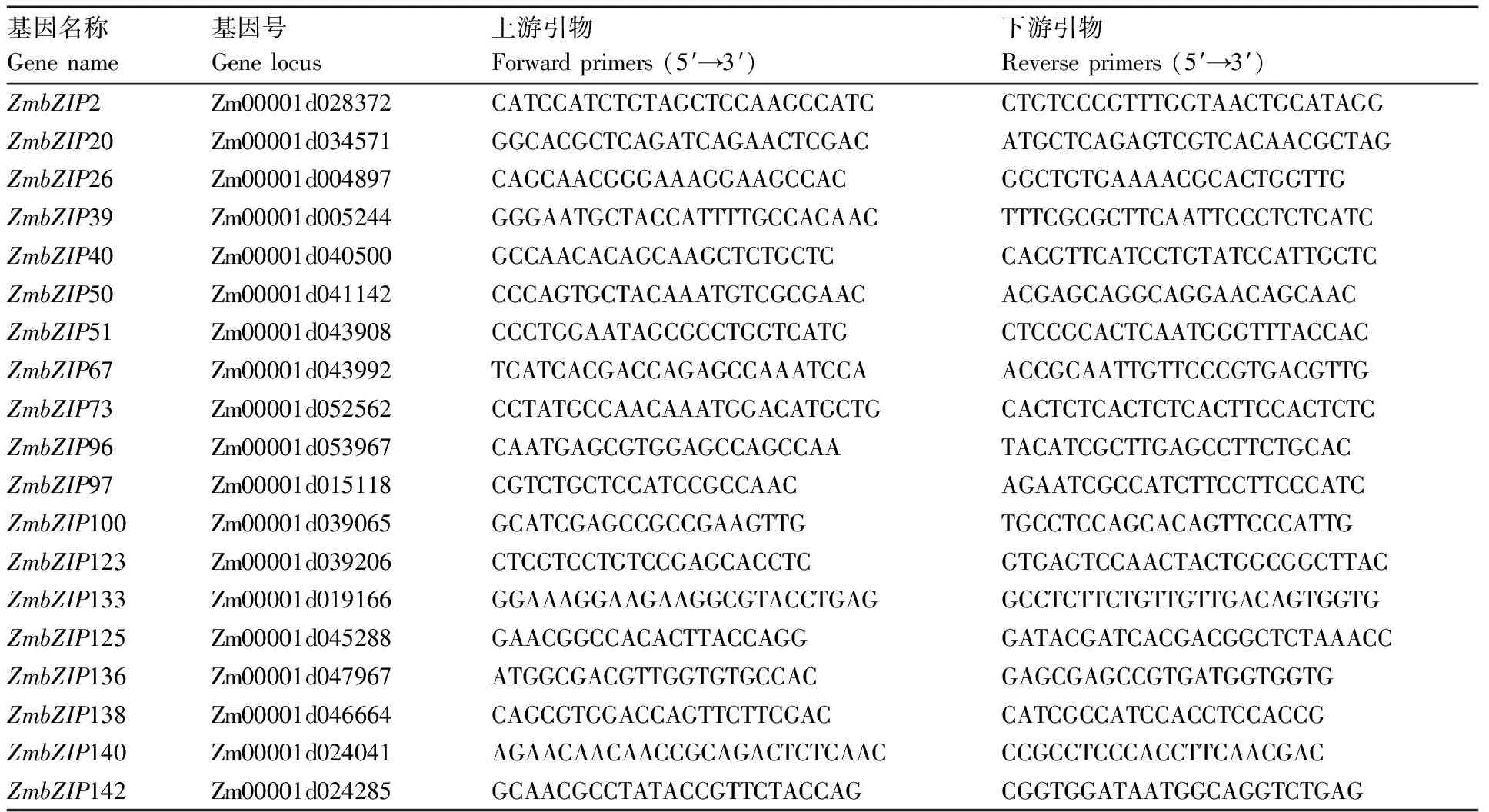
表1 ZmbZIP基因的实时荧光定量引物
2 结果与分析
2.1 bZIP亚家族生物信息学分析
为了探测玉米bZIP家族基因的基本表达信息,根据前人的研究结果,选择玉米bZIP家族中G亚家族基因共计20个作为研究对象,20个基因的基本信息见表2。20个基因分布于8个染色体上,其中,Chr3和Chr10上各有4个基因,Chr1、Chr2、Chr4、Chr6和Chr9上各有2个基因,Chr5和Chr7上各有1个基因(图1-A)。20个基因内含子数目变异较大,67和97没有内含子,50有14个内含子(图1-B)。20个ZmbZIP蛋白N端含有典型的bZIP结构域,包括保守的碱性蛋白序列和亮氨酸拉链蛋白序列(图2-A)。进化树分析结果显示,20个ZmbZIP蛋白和6个拟南芥AtbZIP蛋白聚合在一起,形成一个家族,推测这些蛋白起源于一个共同的祖先;同时,玉米的基因发生了明显的基因复制,基因数量比拟南芥中的同源基因多(图2-B)。
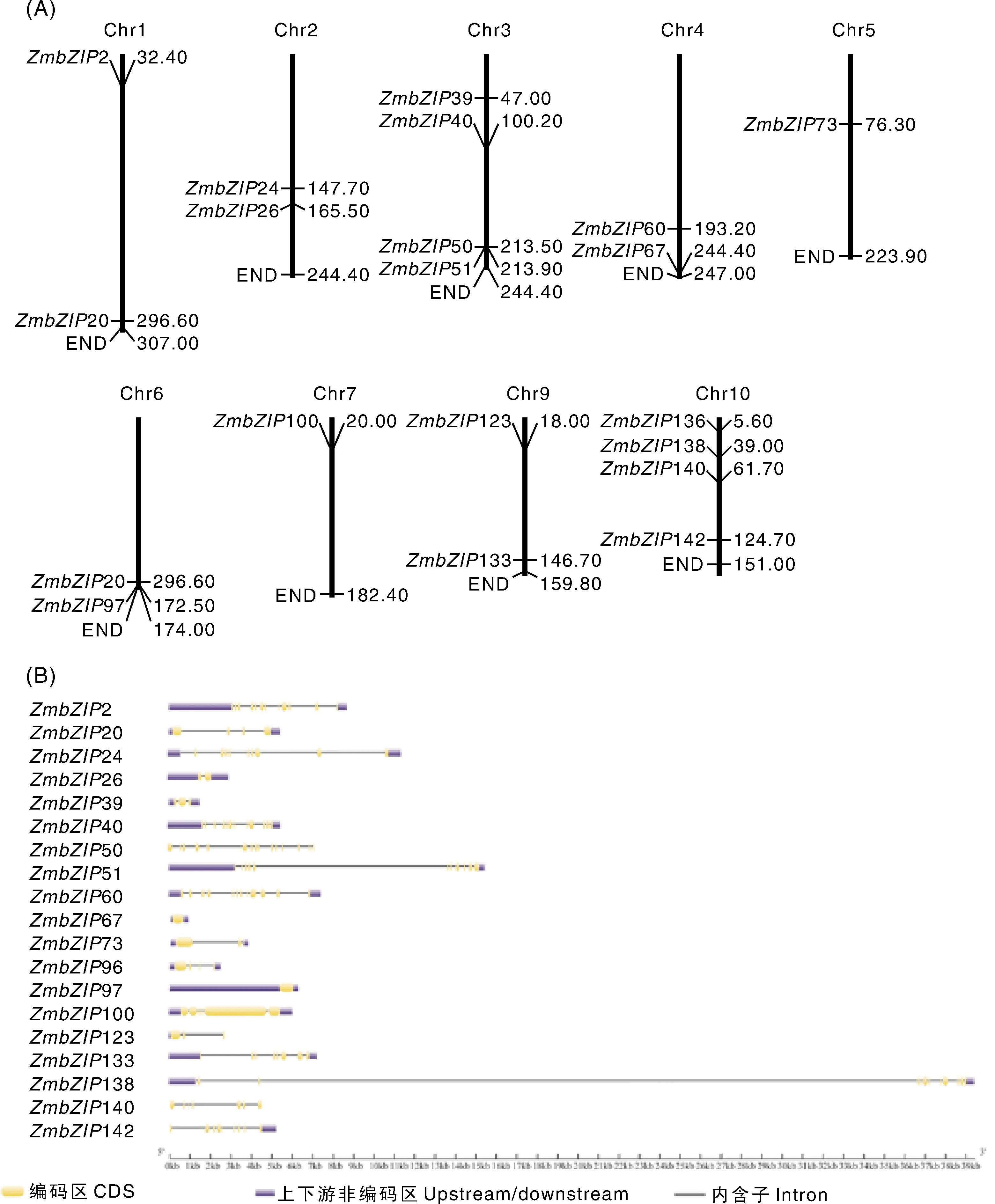
A,ZmbZIP基因的染色体定位;B,ZmbZIP的基因结构。

A,ZmbZIP蛋白保守结构域多重序列比对;B,ZmbZIP蛋白的进化树。

表2 玉米bZIP家族基因基本信息
2.2 ZmbZIP在玉米组织中的表达模式
为了研究基因在玉米不同组织部位的表达模式,利用qRT-PCR技术,分析了这些基因在玉米根系、茎、叶、雄花和嫩穗中的表达水平。结果显示,15个基因在5个组织中的表达模式不同(图3)。5个基因(26、40、73、100和140)主要在根系中表达,其表达量是其他组织器官的2倍以上。50和142主要在嫩穗中表达,分别比其他部位高70倍和3倍以上,表明这2个基因可能在玉米穗的形成过程中发挥重要作用。2主要在雄花中表达,是其他部位的2倍以上;20呈现组成型表达模式,在探测的5个组织器官中都有较高的表达水平。6个基因(39、51、67、96、97和138)主要在2个或2个以上的组织器官中表达,预示这些基因功能具有多样性。
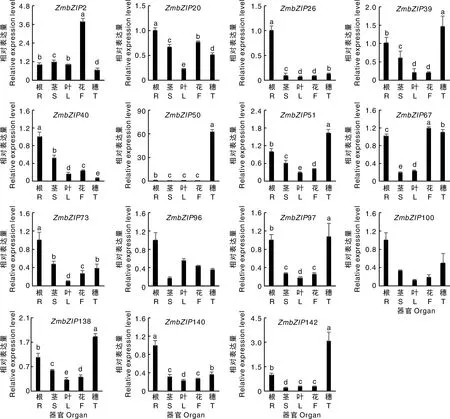
R、S、L、T、E分别代表根、茎、叶、雄花和幼穗。柱上无相同小写字母代表差异显著(P<0.05)。下同。
2.3 ZmbZIPs应答盐、干旱和低温胁迫的表达模式
由图4可知,玉米叶片中不同基因在3种逆境胁迫下呈现不同的表达模式。51在NaCl、PEG6000和低温胁迫处理6 h表达量达到了最高值,分别比处理前上升了4.8倍、10.3倍和5.0倍,推测该基因参与这3种逆境胁迫响应途径。20、96和142只受NaCl胁迫和PEG胁迫的诱导,并且在胁迫处理24 h时,达到最高值,低温胁迫却抑制这3个基因的表达。NaCl和PEG6000胁迫24 h,20表达量分别是0 h的4.3倍和4.9倍;低温胁迫24 h,该基因表达量下调了68%,这些数据表明这3个基因在应对NaCl、PEG6000和低温胁迫时具有不同的反应机制。NaCl胁迫诱导39表达,低温胁迫抑制其表达,分别是0 h的2.4倍和51%。7个(2、26、40、67、73、100、138)在应对3种逆境胁迫时表达量变化不超过2倍,这些基因在玉米抵御逆境胁迫时的作用还有待深入研究。
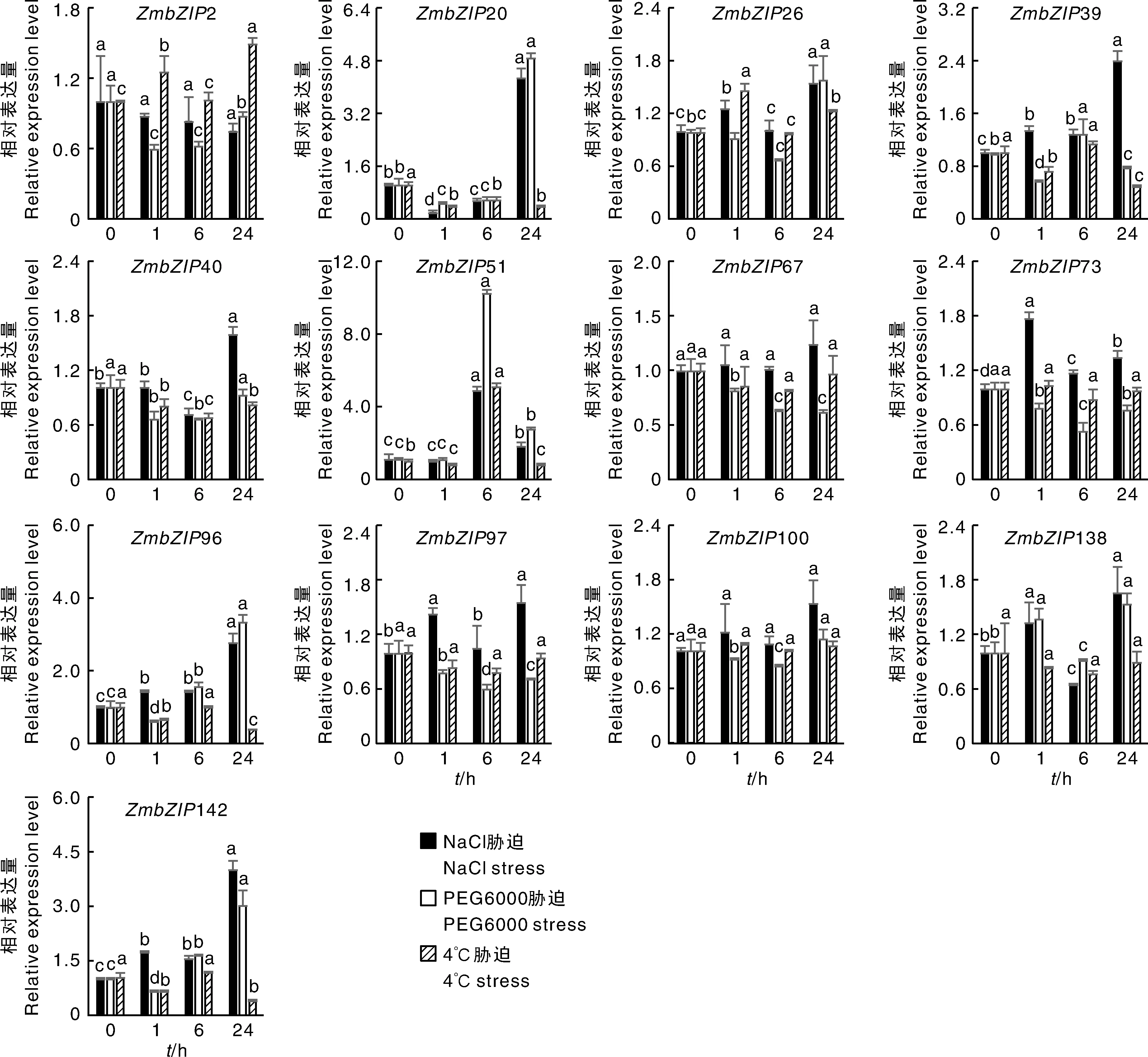
图4 不同逆境胁迫下叶片中ZmbZIP基因的相对表达量
2.4 ZmbZIP应答氨态氮和硝态氮胁迫的表达模式
由图5可知,在不同氮形态缺乏胁迫下,玉米叶片中基因的表达模式不同。胁迫处理24 h,20、26、73、97和100的表达同时受2种不同氮缺乏胁迫的抑制,表明这5个基因在调控不同氮形态缺乏胁迫中发挥着类似的作用。硝态氮和铵态氮缺乏胁迫处理24 h,20表达量比处理前分别下降了82%和48%。39、40和138在2种胁迫处理下,表达量先上升再下降。40在铵态氮缺乏胁迫1 h时,其表达量是0 h的48倍,之后出现回落,胁迫24 h时,其表达量相比处理前下降了54%,表明这些基因是应答氮胁迫的早期响应基因,在氮胁迫的早期发挥作用。50、51、67、96和142的表达在应答硝态氮和铵态氮缺乏胁迫时的表达模式不同。如96受铵态氮缺乏胁迫的持续强烈诱导,并在胁迫处理24 h达到最高值;在硝态氮缺乏胁迫下,该基因先受到强烈诱导,其表达量比处理前上升了230倍,之后出现明显回落,表明该基因是硝态氮缺乏胁迫的早期响应基因,在硝态氮和铵态氮缺乏胁迫中都发挥着独特的作用。

图5 氮缺乏胁迫下玉米叶片中ZmbZIPs的相对表达量
由图6可知,玉米根中14个基因的表达受不同氮形态缺乏胁迫的调控。在胁迫处理24 h,2、26、40、51、73、97、100、138和142的表达同时受硝态氮或铵态氮缺乏胁迫的抑制,表明这些基因在这2种胁迫响应途径中可能具有类似的功能。比如151在硝态氮或铵态氮缺乏胁迫24 h时,其表达量相比处理前分别下降了90%和99%。50则同时受2种胁迫处理的诱导,实验结束时,其表达量分别是处理前的12.8倍和4.1倍。67在胁迫早期同时受2种胁迫处理的诱导,之后出现回落,表明这2个基因在2种胁迫响应途径中也具有类似的作用。在硝态氮或铵态氮缺乏胁迫下,96的表达模式不同,硝态氮缺乏胁迫下,该基因受到持续的诱导,在实验结束时,其表达量是处理前的28倍;而铵态氮缺乏胁迫则先诱导该基因的表达,在胁迫处理1 h时,其表达量上升了10倍以上,之后回落到处理前的水平,表明该基因是铵态氮缺乏胁迫早期响应基因,该基因在2种不同的氮形态缺乏胁迫响应途径中具有不同的功能。
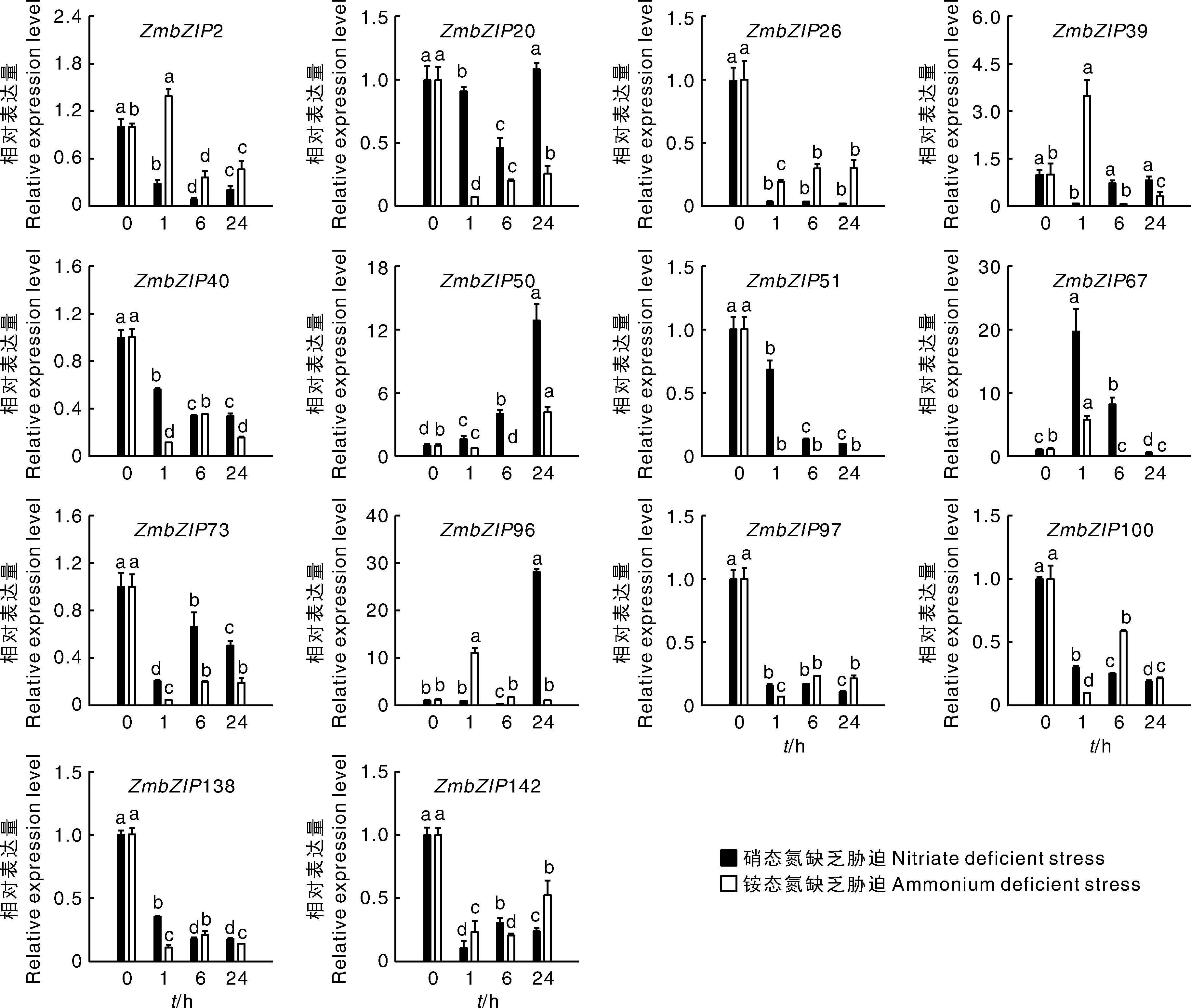
图6 氮缺乏胁迫下玉米根系中ZmbZIPs的相对表达量
3 结论与讨论
植物bZIP成员众多,生物学功能多样,一般情况下,根据植物bZIP转录因子的结构特点和功能划分为A、B、C、D、E、F、G、H、I和S共10个亚家族。玉米bZIP家族G亚家族可以进一步分化为3个亚组,这与前人的研究结果基本一致,表明了这20个基因在玉米进化过程中,基因序列发生了明显的分化,这也体现在这些基因在组织表达模式或逆境响应表达模式上有明显差异。例如39、51、142、50和96的序列进化关系比较近,组成一个亚组,但这5个基因的组织表达模式发生明显分化,其中,39、51和142的组织表达模式类似,在幼穗中的表达量最高,其次是根系,体现了基因进化进程中的保守性,50则呈现为幼穗特异性表达模式,96在根系中表达量最高,出现了基因表达的分化;这5个基因都显著受高盐胁迫诱导,而在PEG6000和低温胁迫下,表达模式发生了变化,142、96和51都受干旱胁迫的诱导,39的表达变化不明显;39、96和142都受低温胁迫的抑制,而51在低温胁迫下则明显受到诱导。其他亚组的基因应答不同逆境胁迫时也表现出这一特点,这种表达模式体现了基因进化过程中的保守性和分化性。
bZIP家族成员在不同植物中的组织表达模式差异非常大,显示了其进化过程中的功能分化。7个马铃薯基因呈现组成型表达模式,其中,同属G亚家族的16和55在根系中表达量比较高,而辣椒G亚家族基因在根系中表达量普遍不高。本研究中,大部分G亚家族成员在根系中表达量比较高,可能和G亚家族基因参与脱落酸(abscisic acid,ABA)响应途径有关,41、54和55显著受干旱胁迫诱导,而受盐胁迫诱导不显著;本研究中盐胁迫对基因表达的影响大于干旱胁迫,反映了不同植物功能的进化差异。玉米51基因参与调控ABA信号途径,在胚胎发育进程中表达量比较高;本研究中,该基因在幼穗中的表达量比较高,并且受干旱、高盐和低温胁迫的强烈诱导,与前人的研究结果基本一致,显示了该基因功能在不同种质中的保守性。玉米自交系Yu882中,20、39和96明显受到高盐胁迫的诱导,本研究结果与此一致,体现了基因功能的保守性。本研究中,26、67、73和100在逆境胁迫下表达量变化不显著,说明这些基因在不同玉米种质中功能的分化,这在其他研究中也有报道。
植物根系主要依靠硝态氮转运体或铵态氮转移体从土壤中吸收养分,这一过程受一系列精密的分子生理调控,其中,转录因子发挥着多样而又重要的作用。本研究中,玉米根系或叶片的bZIP家族基因广泛受到硝态氮或铵态氮缺乏胁迫的调控,同时还存在组织器官表达差异。在硝态氮或铵态氮缺乏胁迫下,根系或叶片中97同时受2种胁迫处理的抑制,而其他基因的表达模式在叶片或根系中存在明显差异,预示在调控氮代谢平衡体系中不同器官的基因存在生物学功能差异。
