盐胁迫对培忠杉苗木离子分布的影响
2022-03-02史锋厚张于卉尹中明沈永宝
史锋厚,宋 珂,郭 锐,张于卉,尹中明,沈永宝
(1.南京林业大学 a.林学院;b.南方现代林业协同创新中心,江苏 南京 210037;2.上海市林业总站,上海 200072)
土壤盐渍化是一个世界性生态问题,全球约7%的陆地表面受到盐渍化的威胁[1]。中国是土壤盐渍化较为严重的国家之一,盐渍土总面积约为3.6×107hm2,占全国可利用土地总面积的5%,严重制约着农林牧业生产,对生态环境也造成了一定的影响[2]。长期以来,国内外在盐渍土改良利用方面已经进行了大量深入研究。与物理、化学及农艺改良措施相比,通过种植耐盐植物改良盐渍土具有投资少、见效快、收益大等优点,既能绿化环境,又能产生一定的经济效益。因此,筛选和培育耐盐植物对于盐渍地的改良具有重要的现实意义[3]。
培忠杉×Taxodiomera peizhongii为我国林木育种专家叶培忠教授以墨西哥落羽杉Taxodium mucronatum和柳杉Cryptomera fortunei进行杂交获得的远缘杂交种,该树种具有生长速度快、落叶期短、抗盐碱、适应性强和绿化效果好等特性,在沿海、沿江滩地防护林建设中具有较高的推广价值[4]。前人围绕培忠杉的耐盐性进行了一系列的研究。陈端钦等[5]研究发现,培忠杉具有一定的抗滨海沙地土壤贫瘠能力,但土壤盐分含量对培忠杉生长影响较大,在盐含量为0.789 g·kg-1的条件下可以生长正常,而在盐含量为8.928 g·kg-1下已致死,即培忠杉适合在没有盐渍化或轻度盐渍化的滨海沙地上造林;而孔强等[6]、张晓磊等[7]均集中开展对盐胁迫下培忠杉苗木生长及生理生化变化方面的研究;其中,张晓磊等[7]研究认为,培忠杉苗木的耐盐临界值在0.4%左右。但以上研究均未从盐离子在培忠杉苗木不同部位间累积与分配的角度探讨其耐盐机制。因此,本研究将采用NaCl 溶液浇灌的方法对3年生培忠杉苗木进行盐胁迫试验,测定培忠杉苗木根、茎、叶各部位Na+、K+、Ca2+等离子含量的变化,深入了解培忠杉苗木在盐胁迫下的适应机制,为培忠杉苗木在盐渍化区域栽培与应用提供科学依据。
1 材料与方法
1.1 试验材料
培忠杉苗木由上海市林业总站提供,为2年生扦插苗。3月下旬,在南京林业大学白马教学科研基地(南京市溧水区白马镇),将苗木移栽至高25 cm、口径20 cm 的塑料花盆中,每个塑料花盆种植1 株;种植基质为镇江兴农有机栽培基质和黄土混合物,混合体积比例为1∶2;露天摆放整齐,正常水分管理。5月中旬将盆栽苗移入塑料大棚温室中,正常水分管理,缓苗1年。于翌年6月选取长势基本一致的120株培忠杉苗木进行盐胁迫试验。
1.2 试验方法
盐胁迫试验采取随机区组设计,4 个处理,每个处理30 株苗木。试验前先采用烘干残渣法[8]测定盆栽土壤积盐层中的总含盐量,计算需要浇灌的NaCl 质量(按盆土干质量计,扣除脱盐土中0.1%的含盐量),在6月12日进行浇盐处理,将称量好的NaCl 用水溶解至250 mL,一次性加入盆中,待盆土吸收后,加自来水至土壤水分饱和,使土壤中可溶性盐的最终含量分别达到0.3%、0.4%和0.5%(干物质含量),对照组浇水至土壤水分饱和。若有盐水下漏到托盘中,可在适当时间返浇入盆,加盐后根据土壤干湿度,每隔2~5 d 浇自来水一次。盐胁迫第92 d 时进行采样,用以测定苗木不同部位的离子含量。
1.3 测定方法
盐胁迫第92 天时,每个处理组随机选取10株苗木,挖出整个植株,于水中清洗干净,按根、茎、叶部位分开,放置在80~90℃的鼓风烘箱中杀青30 min,然后在65℃下烘12~14 h,直至质量不再减轻为止。粉碎,过2 mm 筛孔。
离子测定参照LY/T1270—1999 和LY/T1271—1999 操作。采用硫酸—高氯酸消煮,用电子天平称取0.8 g 烘干样品放置于凯氏瓶中,加10 mL混合酸(硫酸与高氯酸体积比为9∶1),混匀,放置过夜,同时做两个空白试验。次日在调温电炉上消煮样品,把温度控制在瓶内样品与酸作用后产生的泡沫不到达瓶颈处,并只有少量的烟从瓶口冒出为度。直至无色透明后再消煮20 min,整个消煮过程经常摇动凯氏瓶,以防局部烧干。消煮完后使用去离子水将消煮液稀释,定容至100 mL作为测定钾、钠和钙的待测液。使用原子吸收光谱测定相应的离子值,再根据稀释倍数、样品干重和分取液体积计算钾、钠和钙含量。
1.4 数据分析与处理
采用SPSS 24.0 软件进行数据统计分析,单因素方差分析法进行差异显著性分析,LSD 方法进行多重比较,并采用Origin 2018 软件绘制相关图表。
2 结果与分析
2.1 盐胁迫对培忠杉苗木不同器官离子含量的影响
由图1 可知,培忠杉苗木体内的Na+总量随着土壤含盐量的增大而不断增大。不同盐含量处理对培忠杉苗木不同部位的Na+含量具有显著性影响(P<0.05)。对照组苗木体内的Na+主要分布在叶部,根部和茎部的Na+含量仅为叶部的31.14%和32.93%。而3 种盐含量处理组苗木体内的Na+则主要分布在根部和叶部,均显著大于茎部(P<0.05);其中,土壤含盐量为0.3%和0.4%的处理组苗木根部和叶部的Na+含量差异不显著(P>0.05),而土壤含盐量为0.5%的处理组苗木叶部Na+含量则显著高于根部(P<0.05),为根部的1.55 倍。此外,与对照组相比,土壤含盐量为0.3%、0.4%和0.5%的处理组苗木根部和叶部的Na+含量分别增加了146%、312%、267%,7%、53%、76%。由此可知,培忠杉苗木根部Na+积累的速率远大于在叶部积累的速率。
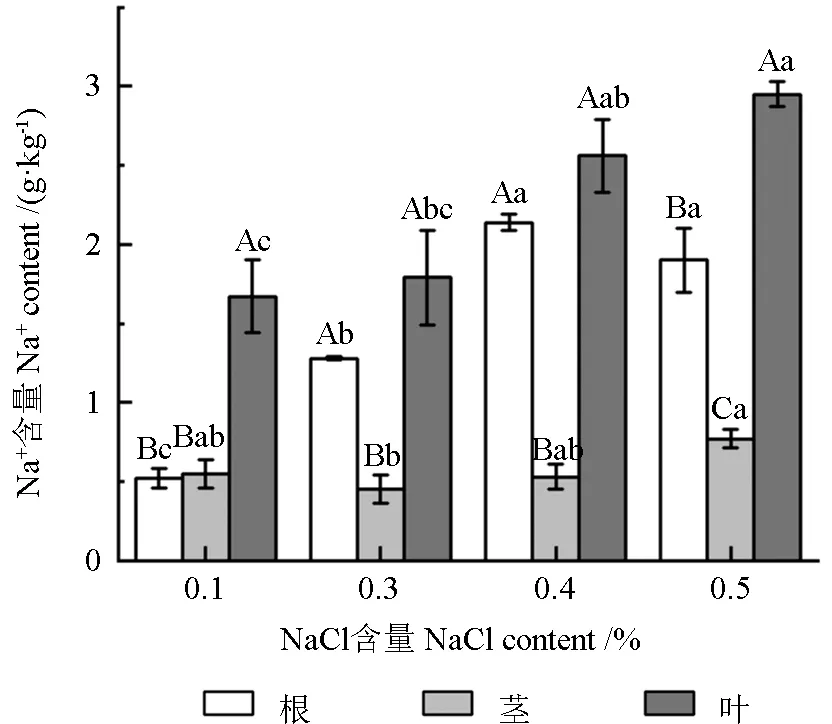
图1 不同盐含量处理下培忠杉苗木不同部位的Na+含量Fig.1 Na+ contents in different parts of ×Taxodiomera peizhongii seedlings under different salinity conditions
由图2 可知,与对照组相比,3 种盐含量处理组培忠杉苗木体内的K+总量变化幅度较小。不同盐含量处理对培忠杉苗木不同部位的K+含量具有显著的影响(P<0.05)。同一盐含量处理条件下,培忠杉苗木不同部位的K+含量整体表现为:叶部>茎部>根部。其中,苗木叶部的K+含量随着土壤含盐量的增大而增大,同比增长16.80%、18.39%、30.32%,均显著大于对照组(P<0.05)。苗木茎部的K+含量均呈负增长,土壤含盐量为0.3%的处理组苗木茎部的K+含量略有下降,但差异不显著(P>0.05);土壤含盐量为0.4%和0.5%的处理组苗木茎部的K+含量显著下降16.86%和12.46%(P<0.05)。而不同盐含量处理条件下,培忠杉苗木根部的K+含量与对照相比均未发生显著变化(P>0.05)。
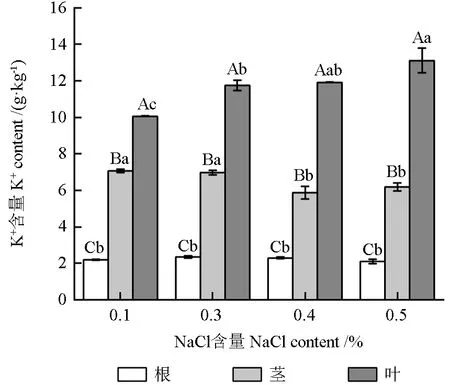
图2 不同盐含量处理下培忠杉苗木不同部位的K+含量Fig.2 K+ contents in different parts of ×Taxodiomera peizhongii seedlings under different salinity conditions
由图3 可知,与对照组相比,3 种盐含量处理组培忠杉苗木体内的Ca2+总量均明显增大。不同盐含量处理对培忠杉苗木不同部位的Ca2+含量具有显著性影响(P<0.05)。同一盐含量处理条件下,培忠杉苗木不同部位的Ca2+含量整体表现为:茎部>叶部>根部。其中,3 种盐含量处理组苗木根部的Ca2+含量与对照组相比差异不显著(P>0.05),而苗木茎部和叶部的Ca2+含量与对照组相比均显著增大(P<0.05),但不同盐含量处理组之间的Ca2+含量无显著差异(P>0.05)。

图3 不同盐含量处理下培忠杉苗木不同部位的Ca2+含量Fig.3 Ca2+ contents in different parts of ×Taxodiomera peizhongii seedlings under different salinity conditions
2.2 盐胁迫对培忠杉苗木不同器官离子含量比值的影响
由图4 可知,同一处理组培忠杉苗木不同部位的K+/Na+比值整体表现为:茎部>叶部>根部。与对照组相比,3 种盐含量处理组苗木根部的K+/Na+比值分别显著下降58.03%、75.23%和74.08%(P<0.05)。3 种盐含量处理组苗木茎部的K+/Na+比值与对照组相比差异不显著(P<0.05),但3 种盐含量处理组之间苗木茎部的K+/Na+比值存在差异,土壤盐含量为0.5%的处理组苗木茎部的K+/Na+比值与土壤含盐量为0.3%的处理组相比显著下降(P<0.05)。不同处理组之间苗木叶部的K+/Na+比值变化幅度较小,差异不显著(P>0.05)。

图4 不同盐含量处理下培忠杉苗木不同部位的K+/Na+比值Fig.4 K+/Na+ ratios in different parts of ×Taxodiomera peizhongii seedlings under different salinity conditions
由图5 可知,同一盐含量处理条件下,培忠杉苗木茎部的Ca2+/Na+比值显著高于根部与叶部(P<0.05),根部与茎部差异不显著(P>0.05)。与对照组相比,土壤含盐量为0.3%、0.4%和0.5%的处理组苗木根部的Ca2+/Na+比值分别显著下降133.58%、77.53% 和73.53%(P<0.05),但处理间差异不显著(P<0.05)。苗木茎部和叶部的Ca2+/Na+比值变化幅度较小,差异不显著(P>0.05)。

图5 不同盐含量处理下培忠杉苗木不同部位Ca2+/Na+比值Fig.5 Ca2+/Na+ ratios in different parts of ×Taxodiomera peizhongii seedlings under different salinity conditions
3 讨论与结论
3.1 讨论
在盐胁迫条件下,植物对Na+、Cl-等盐离子的大量吸收是引起渗透胁迫、离子毒害及营养失衡的主要原因[9]。盐分对植物的作用可能涉及一系列形态、生理、生化和分子变化,对种子萌发、植物生长和生产力产生不利影响,但在对盐胁迫长期适应的过程中,不同植物已经进化形成了特定的应对策略[10-11]。在本试验中,培忠杉苗木体内的Na+总量随着土壤含盐量的增大而增大,且各处理组培忠杉苗木在盐胁迫10 d 时已经表现出盐害症状(叶片尖端开始枯黄),随着盐胁迫时间的延长和土壤含盐量的增大,苗木盐害症状逐渐加重,即主要表现为小叶枯黄,伴有二级分枝的脱落,并且只要叶基部出现淡黄色,叶片就一触即掉;但直至试验结束时也未出现苗木死亡的现象,这证明了培忠杉苗木具有一定的耐盐性,且在应对盐胁迫的过程中形成了自身的适应策略。
Na+的吸收、转运和分布特性是植物在盐胁迫条件下能否正常生长的关键。通过对培忠杉苗木不同部位的Na+含量的测定分析可知,培忠杉苗木的耐盐策略是首先将Na+运输至叶片,对照组苗木亦是如此,其次将Na+积累在根部,茎中的Na+含量最少。一方面,培忠杉苗木的叶片较小而且数量多,将Na+积累在成熟叶片、次级分枝和叶子密集的地方,可以通过形成离层的方式使叶片和小枝脱落来减少苗木体内的盐分积累量,这与四季竹Oligostachyum lubricum的耐盐策略相似[12]。另一方面,苗木可以通过提高叶片中Na+含量来降低渗透势,以促进水分向地上部分的运输,缓解盐胁迫造成的生理干旱[13]。此外,培忠杉苗木根部Na+积累的速率远远大于在叶部积累的速率,推测可能是由于培忠杉苗木根部存在盐离子的区隔化作用[14],能够主动将Na+控制在根部,从而减少过多的Na+运输至地上部分,降低Na+对苗木地上部分的毒害作用。
在盐胁迫条件下,植物常遭受Na+毒害和K+、Ca2+亏缺的双重危害。植物体内Na+含量过高会抑制植物对K+、Ca2+等营养元素的吸收[15]。钾(K)作为植物体内保证植物正常代谢和生长发育的大量营养元素之一,具有调节离子平衡、渗透压、细胞膨压和光合作用等生理功能[16-17]。由于Na+和K+的物理化学性质相似(即离子半径和离子水合能),Na+会竞争细胞质关键代谢过程中K+的主要结合位点,从而导致K+的流失[18]。在本试验中,3 种盐含量处理组苗木体内的K+总量变化幅度较小,并未出现下降的现象,在对唐古特白刺[19]Nitraria tangutorum、欧洲落叶松[20]Larix deciduaMill.的研究中也有类似结果,表明盐胁迫对培忠杉苗木K+的吸收抑制作用影响较小。此外,3 种盐含量处理组苗木根部的K+含量与对照相比差异不显著,而叶部的K+含量则均显著增大,茎部的K+含量则均呈负增长,说明苗木在盐胁迫条件下,其体内的K+被重新分配,苗木茎部的K+更多地被运输至叶片中,这与对流苏[21]Chionanthus retusus、 鹅耳枥[22]Carpinus turczaninowiiHance 的研究结果具有相似之处。植物叶片内K+含量的增大有利于叶部维持正常生理代谢,是培忠杉苗木适应盐胁迫的一种正常表现。
钙(Ca)是植物细胞膜的主要成分,起着维持细胞膜结构和功能的作用[23]。高含量的Na+可置换质膜和细胞内膜系统所结合的Ca2+,但由于Na+和Ca2+的电荷密度不同,这种置换破坏了膜结构,增加了膜的通透性,致使细胞内K+、P 和有机溶质外渗[24]。在本试验中,3 种含盐量处理组苗木体内的Ca2+总量与对照组相比均明显增大。且茎部和叶部的Ca2+含量均显著上升,根部的Ca2+含量则与对照相比差异不显著,表明培忠杉苗木在盐胁迫条件下将更多的Ca2+运输至地上部分。黄璐瑶等[25]对金银花Lonicera japonicaThunb的研究中发现,在盐胁迫条件下,施用外源Ca2+将导致金银花幼苗体内的K+含量增大,而Na+含量明显降低。因此,培忠杉苗木地上部分Ca2+含量的增大可能是维持其地上部分Na+和K+平衡,从而适应盐胁迫的一种重要机制。
植物正常生长发育需要维持细胞内离子的相对平衡状态。在盐胁迫条件下,各种元素与Na+比值的变化更能反映植物叶片受伤害的程度,特别是反映植物整体的营养平衡状况,其中,K+/Na+、Ca2+/Na+值是评定树种受盐害程度和耐盐性的重要指标[26-27]。在本试验中,培忠杉苗木根部的K+/Na+、Ca2+/Na+比值均显著下降,结合上述有关K+和Ca2+含量的变化分析,产生这种变化的主要原因是苗木根部的Na+含量的净升高,即3 种盐含量处理组苗木根部的离子平衡被打破。相反,3 种盐含量处理组苗木叶部的K+/Na+、Ca2+/Na+比值则与对照组差异不显著,说明培忠杉苗木地上部分仍能维持离子平衡,保证了苗木叶片在盐胁迫条件下仍能进行正常的生理代谢。同时,这也证明了K+/Na+比值和Ca2+/Na+比值比单一的K+和Ca2+含量更能反应树种的耐盐特性。
3.2 结论
综上所述,从离子吸收及分布的角度分析可知,培忠杉苗木的耐盐策略主要是:首先将吸收的Na+积累在成熟叶片,随着成熟叶片的脱落来降低树体的盐分总量;其次是将Na+固定在根系中以减少对地上部分的伤害。在一定盐胁迫范围内,维持苗木地上部分K+、Ca2+和Na+的相对平衡状态,减轻盐胁迫条件下的离子毒害和渗透胁迫作用。在本试验中设置的3 种盐含量处理还不能准确地确定培忠杉苗木的耐盐临界点,并且对于培忠杉苗木耐盐反应中的盐分的感应和信号转导,以及相关的基因表达调控等方面的研究较少,未来仍需开展更加深入的研究。
