香榧初结果母枝性状变化规律与结实能力的关系
2022-03-01叶淑媛曾燕如胡渊渊汪舍平喻卫武
叶淑媛,曾燕如,胡渊渊,龙 伟,汪舍平,喻卫武
(1. 浙江农林大学 省部共建亚热带森林培育国家重点实验室,浙江 杭州 311300;2. 中国林业科学研究院 亚热带林业研究所,浙江 杭州 311400;3. 常山县林业水利局,浙江 常山 324200)
香榧Torreya grandis‘Merrillii’属裸子植物红豆杉科Taxaceae榧树属Torreya常绿乔木,为第三纪孑遗植物[1],是榧树Torreya grandis中唯一优良栽培类型,也是中国特有的珍贵经济干果树种。由于香榧具有极高的经济价值,产值超18亿元,近年香榧产业向榧树分布的省市推广扩展,发展速度位居山地经济林之首[2]。结果母枝是着生结果枝的基枝,是香榧生长发育所需要营养的主要输送通道,对种实表现出来的性状产生很大影响。减少香榧树体的营养消耗,培育重点结果母枝,对增加结果枝的结实能力、坐果率及提高种实的品质有重要的意义。叶淑媛等[3]研究发现:香榧砧苗分枝数越少,表皮越光滑,越有利于嫁接口的愈合;刘萌萌等[4−5]研究了香榧叶片中矿质元素年周期季节性变化规律及矿质元素动态变化的关系。邬玉芬等[6]研究发现:香榧枝条黄化与钙、镁、铁元素水平低以及氮、锰元素水平过高有关;詹利云等[7]研究发现:香榧叶片长度在居群间差异显著(P<0.05)。但是,目前有关香榧结果母枝性状方面的研究还未见报道。鉴于此,本研究对香榧初结果林的结果母枝分布规律与结实关系及枝叶矿质元素间的关联性进行分析,旨在为香榧科学控梢、培养结果母枝、提高种实产量提供理论依据。
1 研究地区与研究方法
1.1 研究区概况
研究区位于浙江省杭州市临安区锦源村 (29°56′~30°23′N,118°51′~119°52′E)。该区属中亚热带季风气候区南缘季风型气候,温暖湿润,日照充足,雨量充沛,四季分明。6—7月为集中降雨期,年均降水量为1 613.9 mm,降水日158 d,无霜期237 d。研究区内有原生榧树分布,为适宜栽培区。
1.2 材料
供试材料为2007年12月种植的3年生香榧苗,株行距4 m×5 m,树势生长健壮,无病害的初结果香榧林。林分单株地径为9.55~14.97 cm,种植后从未进行过人为树体干预,单株均已开花结果。研究区 0~20 cm 土壤中全氮质量分数为 1 846.19 mg·kg−1,全磷质量分数为 802.85 mg·kg−1,全钾质量分数为9 816.70 mg·kg−1。每年施用复合肥 2~3次,平均 750~1 000 g·株−1·a−1,有机肥隔年施 1次。
1.3 方法
1.3.1 样株选择和测定 用卷尺、游标卡尺分别测定9株香榧株高、冠幅(东西向和南北向的树冠直径计算冠幅)、地径、主干茎周等表观指标。株高小于2.8 m,地径小于11.0 cm为弱势树;株高2.8~3.5 m、地径11.1~13.0 cm为中庸树;株高大于3.5 m、地径大于13.0 cm为强壮树。确定3种类型香榧树作为研究对象,每种类型重复3个。
中庸树的标准:叶色绿,主枝、延长枝生长旺盛,顶芽粗壮,树体中部结果枝数量多,结果枝比例近40%。
1.3.2 结果枝和营养枝数量测定 于2019年5月上旬,待混合芽、营养芽全部开展后,分单株统计3种类型树体的全部混合芽(发展成结果枝)、营养芽(发展成营养枝)数量,计算混合芽比例。
1.3.3 结果母枝粗度和长度测定 将香榧树体高度3等分,划定树体的上、中、下3个层面。于2019年3月(落果前)和6月(落果后),对3种树势树体上、中、下3个层面的东南西北方向各取5个3年生枝条,共540个枝条。图1为2017年生枝条。同时各树势植株上、中、下3个层面的东南西北方向各取1个3年生枝条,3个重复,共108个枝条,用游标卡尺测其结果母枝的粗度和长度,重复3次;根据测定结果,计算整株树结果母枝粗度和长度。

图1 香榧 3年生枝叶测量部位示意图Figure 1 Schematic diagram of measuring parts of the 3-year-old branches and leaves of T. grandis ‘Merrillii’
1.3.4 坐果率和结果量测定 2019年 3月统计单株膨大前的2代果幼果数量,并于7月初(生理落果后),统计枝条着生的膨大果,计算全株2代果坐果率=(7月种实数量/3月种实数量)×100%。
1.3.5 矿质元素测定 将采集的样品分别用自来水、纯净水冲洗干净,再用滤纸干燥,待测。将香榧1年生枝叶去顶端和末梢,取中段枝叶进行测定。将枝条置于烘箱中,105 ℃杀青30 min,然后60 ℃左右烘干至恒量,计算枝叶样品的含水率。用植物粉碎机研磨烘干的枝条,过60目筛均匀混合。用硫酸-过氧化氢(H2SO4-H2O2)联合消煮液消煮待测样品。分别采用凯氏定氮法、钼锑抗比色法、原子吸收分光光度法[8]测定氮、磷、钾、钙、镁、锰、锌等质量分数。
1.4 数据分析
利用SPSS 22.0开展单因素方差分析和相关分析,采用最小显著差数法(LSD法)进行平均数间多重比较,计算变异系数和回归系数,并用Excel 2007作图。
2 结果与分析
2.1 香榧树体生长结果情况
由表1可知:香榧单株树体营养枝数量为5 134条,结果枝数量为2 909条,结果母枝粗度2.34 mm、长度8.57 cm,膨大果数量538个,坐果率3.78%。其中冠幅、结果母枝粗度和膨大果数量均在中庸树最大,而结果母枝数量则在中庸树最少,树势越强坐果率越低,其余参数均随树势增加而增大。

表1 9棵香榧树体基本情况与调查结果Table 1 Basic information and investigation results of 9 T. grandis ‘Merrillii’ trees
2.2 结果母枝与结实性状特征分析
如表2所示:单株间的结果母枝粗度、长度、混合芽比例、膨大果数量和坐果率的性状均存在较大变幅,其中坐果率变幅最大,变异系数为53.44%;结果母枝粗度、长度变幅分别为9.85%、8.04%。表明香榧树单株间的结果母枝粗度和长度差异较小,受树势影响较小;混合芽比例、膨大果数量及坐果率在树体单株间变化较大。

表2 结果母枝及结实性状Table 2 Seed-bearing mother shoots and seed-bearing traits
2.3 树势间、层面间结果母枝与结实指标的表现
2.3.1 树势间结果母枝性状及结实指标多重比较 如表3所示:3种树势的结果母枝数量均无显著差异(P>0.05);中庸树的结果母枝粗度最粗,与其余2种树型呈显著差异(P<0.05),强壮树与弱势树的结果母枝粗度无显著差异(P>0.05);中庸树和强壮树的结果母枝长度无显著差异(P>0.05),但两者均显著大于弱势树(P<0.05);弱势树的混合芽比例显著高于其他2种树型(P<0.05),坐果率在3种树势间均存在显著差异(P<0.05),两者从大到小依次均为弱势树、中庸树、强壮树。
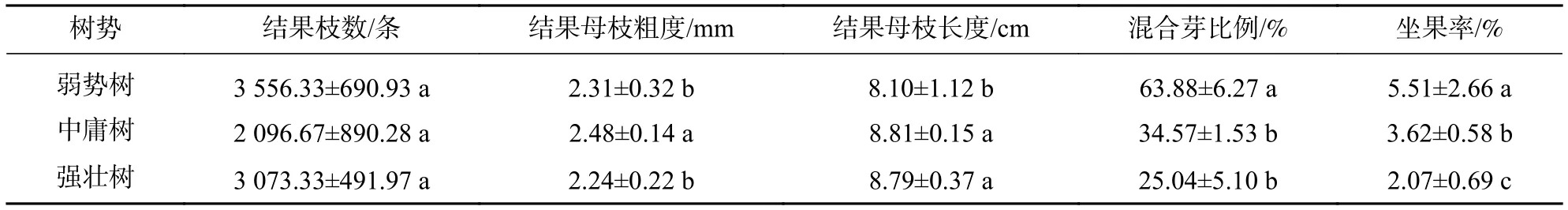
表3 树势间结果母枝及结实性状的多重比较Table 3 A multiple comparison in seed-bearing mother shoots and seed-bearing traits among trees of various vigor
2.3.2 层面间结果母枝性状及结实指标多重比较 如表4所示:3种树势的结果枝数量均无显著差异(P>0.05);结果母枝粗度在树势间均为上层最粗,下层最细(P<0.05),其中中庸树的上层、中层无显著差异(P>0.05);弱势树和强壮树的结果母枝长度均是上层最长,其中弱势树中层、下层无显著差异(P>0.05),强壮树则是下层最短,中层居中(P<0.05),中庸树层面间无显著差异(P>0.05);中庸树与弱势树均是下层的坐果率最高,上层的最低(P<0.05),其中中庸树的上层、中层之间的坐果率无显著差异 (P>0.05)。

表4 层面间结果母枝及结实性状的多重比较Table 4 A multiple comparison in seed-bearing mother shoots and seed-bearing traits among layers of a crown
2.4 结果母枝粗度和长度分析
2.4.1 结果母枝数量分布 图 2所示:粗度在 1.91~2.50 mm 的结果母枝数量最多,占44.49%;粗度在2.51~3.10 mm的结果母枝数量次之,占26.40%;粗度≤1.90 mm和≥3.11 mm的结果母枝数量最少,分别占21.83%和7.28%。长度在5.51~8.70 cm的结果母枝数量最多,占57.56%;长度在8.71~11.90的结果母枝数量次之,占38.92%;长度≤5.50和≥11.90的结果母枝数量均较少,共占3.52%。

图2 结果母枝粗度和长度分布比例Figure 2 Distribution map of thickness and length of fruiting mother branch
2.4.2 粗度、长度区间结果母枝及结实性状多重比较 如表5所示:结果母枝数量、2代果总数和膨大果数量在粗度1.91~2.50 mm最多,2.51~3.10 mm次之,≤1.90 mm和≥3.11 mm最少(P<0.05);此3项指标在长度为5.51~8.70 cm最多,8.70~11.90 cm次之,≤5.50 cm和≥11.90 cm最少(P<0.05);坐果率在不同粗度和长度均无显著差异(P>0.05)。
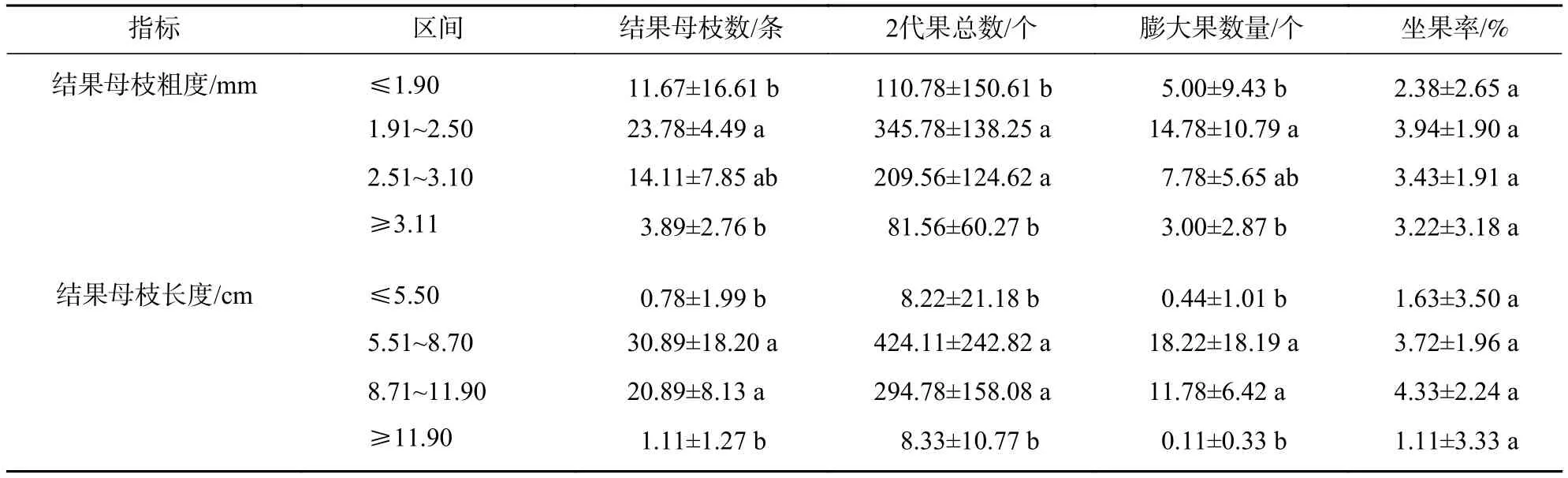
表5 粗度、长度区间结果母枝及结实性状的多重比较Table 5 A multiple comparison in seed-bearing mother shoots and seed-bearing traits in terms of thickness and length
2.4.3 结果母枝粗度、长度、混合芽比例、坐果率的相关分析 如表6所示:结果母枝粗度、长度与混合芽比例均呈负相关,相关系数分别为−0.043、−0.552,相关性均不显著;同样,结果母枝粗度及长度与营养芽比例相关性也不显著。结果母枝粗度及长度与2代果坐果率均呈负相关,其中结果母枝长度与2代果坐果率的相关系数为−0.825,呈极显著负相关(P<0.01)。结果母枝粗度与2代果坐果率相关系数为−0.461,无显著相关;混合芽比例与坐果率的相关系数为0.680,呈显著正相关(P<0.05)。
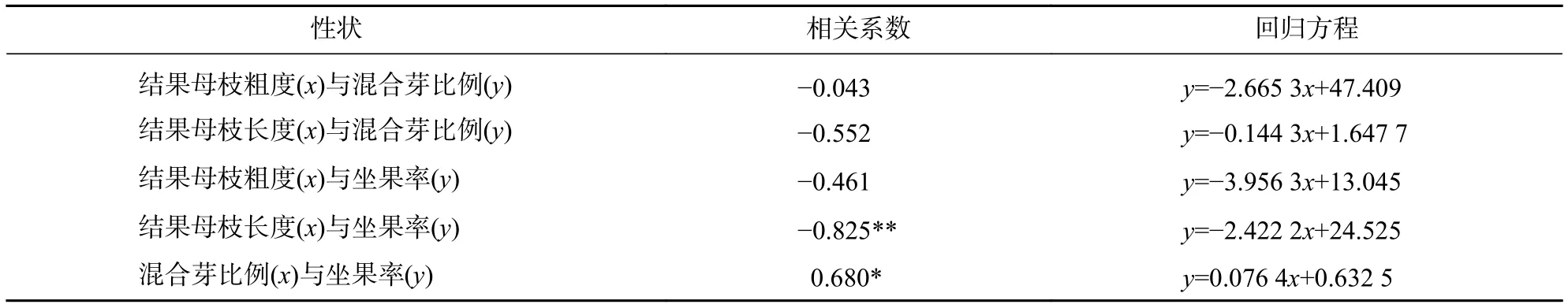
表6 结果母枝与结实性状的相关性分析Table 6 Correlation between seed-bearing mother shoots and seed-bearing traits
2.5 落果前后香榧枝叶矿质元素相关分析
2.5.1 枝叶氮、磷、钾、钙元素多重比较 图3所示:3月香榧枝叶的氮质量分数从大到小依次为强壮树、中庸树、弱势树(P<0.05);而6月中庸树、弱势树氮质量分数极显著高于强壮树(P<0.05),弱势树与中庸树间无显著差异(P>0.05);3和6月的磷、钾质量分数在树势间均无显著差异(P>0.05),但磷、钾质量分数均为6月大于3月;3月的钙质量分数无显著差异(P>0.05),6月强壮树钙质量分数显著低于另外2种树势(P<0.05),弱势树与中庸树间没有显著差异(P>0.05)。其中3月的强壮树、中庸树、弱势树间的磷、钾质量分数接近,6月强壮树磷最低,中庸树钾最低;3种树势的钙质量分数均为3月大于6月,3月3种树势间钙质量分数接近,6月强壮树最低。
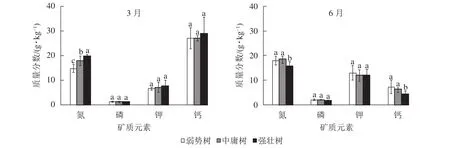
图3 3和 6月枝叶矿质元素之间的多重比较Figure 3 Multiple comparison of mineral elements in branches and leaves of T. grandis ‘Merrillii’ in March and June
2.5.2 枝叶矿质元素与结果母枝形态特性及结实特性之间的相关性分析 如表7所示:3月结果母枝粗度与钙呈显著负相关(P<0.05);混合芽比例与氮呈极显著负相关(P<0.01),与锌、锰呈显著负相关(P<0.05);其余各指标之间无显著相关。6月结果母枝粗度与氮呈极显著正相关(P<0.01),与磷呈显著正相关(P<0.05);结果母枝长度与钙呈显著负相关(P<0.05),与铁呈显著正相关(P<0.05);混合芽比例与锰呈显著的负相关(P<0.05);坐果率与钙、铁分别呈极显著正、负相关(P<0.01)。

表7 3和6月结果母枝、结实性状与枝叶矿质元素相关性分析Table 7 Correlation between seed-bearing mother shoots and seed-bearing traits and mineral elements in March and June
2.6 营养生长与生殖生长的相关性分析
如表8所示:营养芽与混合芽比例呈极显著负相关(P<0.01),与膨大果数量及坐果率呈显著负相关(P<0.05),相关系数分别为−0.830、−0.673和−0.692。
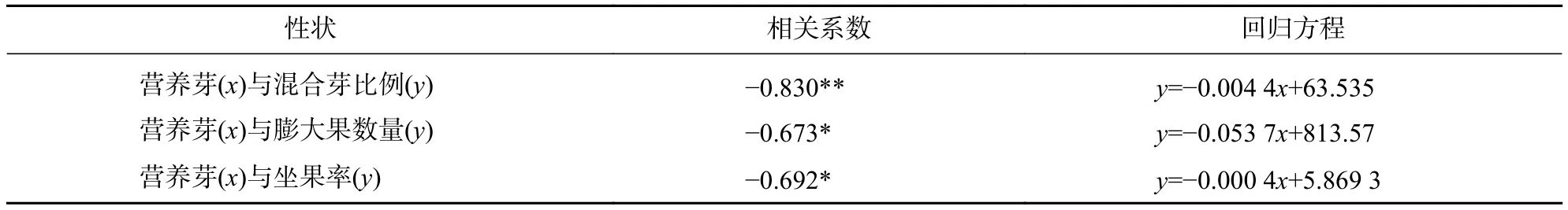
表8 香榧营养生长与生殖生长相关性分析Table 8 Analysis of correlation between vegetative growth and reproductive growth
3 讨论
香榧结果母枝是着生结果枝的基枝,是香榧种实生长发育所需营养的主要供应源,因此结果母枝的生长状况对香榧种实性状及产量有重要影响[9−10]。变异系数的大小反映了性状离散程度的高低,可衡量性状间的变异程度[10]。本研究发现:未在人为干预情况下,结果母枝粗度与长度在单株间较为稳定,而结果枝比例、膨大果数量及坐果率的性状特征可能受树势影响较大,导致其变异程度较大。
本研究结果显示:结果母枝数量与树势、层面均无密切关系;而结果母枝粗度及长度、混合芽比例及坐果率受树势的影响较为明显。中庸树结果母枝粗度、长度均最大,混合芽比例及坐果率在中庸树中表现良好,表明初结果林分香榧树的树势越强,营养生长越强,反之,则生殖生长强于营养生长。中庸树的生殖生长和营养生长较为平衡,可根据生产实际需求,以中庸树为培育目标。同时,结果母枝粗度自下而上均越来越粗;长度在强壮树、弱势树中也是自下而上越来越长;混合芽比例弱势树为下层最高,强壮树为上层最高;坐果率在弱势树和中庸树中均是下层最高,表明树势的强弱、层面的分布均能影响结果母枝的粗度和长度。随着层面的增高,营养生长逐渐增强,这与光照强度由下向上逐渐增加的规律一致[11]。层面越低生殖生长越明显,可能与香榧树下垂枝有关。
结果母枝粗度、长度分布情况与结果母枝数量、2代果总数和膨大果数量(结果量)呈一致性,均在粗度为1.91~2.50 mm最多,长度为5.51~8.70 cm最多,坐果率不受结果母枝粗度和长度的影响。由此可见,初结果林分香榧树的结果性状在不同粗度、长度的分布状况呈现一定的规律,枝粗和枝长是影响结果性状指标的重要因素之一,是能否迅速提高早期产量的关键,也是衡量结果母枝坐果能力的重要指标,这与陈霞等[12]的研究相一致。本研究相关分析显示:与产量相关的坐果率和混合芽比例等参数受结果母枝粗度、长度的制约,结果母枝粗度越粗,长度越大,香榧树的坐果率就越低,特别是枝越长坐果率越低。结果母枝粗度、长度对混合芽、营养芽的生长也有一定的影响,但影响不明显;混合芽比例越高坐果率越高。同时,香榧初结果树的营养生长制约着生殖生长,营养生长越旺盛,越不利于香榧果实的膨大和产量的提高。因此,培养中庸树型,保持营养生长与生殖生长平衡,对提高香榧种实产量有重要意义。
氮、磷、钾是香榧种子生长发育所必需的矿质元素,对香榧苗木生长的影响作用从大到小依次为氮、钾、磷[13]。本研究表明:3和6月的氮在树势间均呈极显著差异,氮质量分数在3月从大到小依次为强壮树、中庸树、弱势树,但在6月强壮树氮质量分数最低,且3月平均值高于6月,可能与香榧种子开始生长膨大,叶片中的氮元素优先供应种子的生长发育有关。本研究结果与刘萌萌等[5]的结论相一致。而6月强壮树氮质量分数下降的幅度高于另外2种树势类型,可能与叶片中的氮元素在供应种子生长和营养枝旺盛生长的同时,消耗了大量氮元素有关,这与其他物种果实发育的研究结论一致[14]。钾能促进果实的增大和质量的增加,尤以在氮营养水平较高时,钾多则效果更为明显,因为钾可提高原生质活性,促进糖的转运,增加果实干质量[15]。磷、钾质量分数不受香榧树势影响,但6月的磷、钾质量分数均高于3月,这可能是由于6月叶片中的磷质量分数相对稳定,钾质量分数有所增加,香榧速生期和种子内部充实期,是对氮、钾需求量最大的时期[5]。香榧花芽分化受到氮、锌、锰的影响,摄入过多的氮、锌、锰会制约混合芽数量增加,这种现象在杏Armeniaca vulgaris树中也存在[16]。3、6月镁与混合芽比例无显著相关,可能是由于该时期枝叶中的镁并不是香榧花芽中镁的供应源[17],表明树体矿质元素随时间季节存在动态变化[5],因此,施肥时要考虑元素间的相互作用。
钙是果胶质中重要物质——果胶酸钙的主要组成物质,也是细胞壁构成成分,果实中钙的高低,影响果实硬度和耐储性[18]。本研究显示:6月铁质量分数的提高促进了结果母枝长度的生长,但抑制了香榧种子的膨大;而钙与枝叶长度呈显著负相关,可能是由于随着香榧种实膨大、细胞分裂迅速,大量的钙元素流向种子,钙元素促进了种子的发育。但当种子由细胞分裂转入增大时,正值新梢旺盛生长期,从而加剧了对钙的竞争。由此可见,种实膨大期适时适度增施钙肥,可以提高香榧坐果率。钙与硼对悬浮细胞的生长有显著的交互作用,增加钙可以部分减缓缺硼对细胞生长的影响[19],植物在缺硼状态下,会触发钙离子的信号转导途径,影响植物体某些蛋白的合成,从而导致植物体的代谢变化[20]。硼、钙与腐植酸的配合施用能够显著提高番茄的株高、果数、单果质量及产量[21]。因此,后续还需开展香榧初结果林中不同钙肥水平对硼等其他元素的响应研究。
4 结论
不同树势和冠层部位的结果母枝类型、结实性状及矿质营养元素特征差异显著。生产实践中,平衡氮肥和钙肥的施用,培养中庸树型,控制香榧结果母枝长度和树体的高度,将树体修剪成开心型,控制结果母枝粗度在1.91~3.10 mm、长度5.51~11.90 cm,有利于提高香榧坐果率和混合芽的比例,对实现香榧丰产、稳产具有重要意义。
