马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性的变化特征
2022-02-25袁亚玲宋思梦
张 艳, 袁亚玲, 李 勋, 宋思梦, 周 扬, 张 健,①
(1. 四川民族学院 横断山区生态修复与特色产业培育研究中心, 四川 康定 626001;2. 四川农业大学 生态林业工程重点实验室, 四川 成都 611130)
马尾松(PinusmassonianaLamb.)是中国第5大树种,为中国南方分布较为广泛的造林树种之一[1-3]。然而,随着马尾松人工纯林林龄增大,林地的生态系统稳定性降低[3,4]。造成这种现象的主要原因是单一树种在吸收土壤养分时具有长期选择性,且单一凋落物不利于养分归还[5]。
凋落物是林地有机质的主要来源,是生态系统养分循环的基础[6]。凋落物分解是一个由土壤和凋落物中酶系统共同调控的复杂生物化学过程[7]。凋落物分解过程中的酶活性可反映凋落物中碳、氮、磷的释放水平[8],以及微生物对相应养分的需求和获取状况[9,10]。酶活性提高可促进凋落物的分解和养分释放,研究凋落物分解过程中酶活性的变化有助于了解凋落物分解在森林生态系统物质循环和能量流动中的作用,对森林土壤肥力提高有重要意义[11]。
研究发现,凋落物的酶活性会随着凋落物多样性及分解过程中基质的变化而发生相应的变化[12-15]。与单一凋落物相比,土壤脲酶、转化酶和脱氢酶活性在混合凋落物分解过程中显著提高[16]。针叶树与阔叶树混合凋落物的分解速率和养分释放明显不同于单一针叶树凋落物[17-20],阔叶树凋落物中的养分可通过淋溶作用或“菌丝桥”转移,进而影响酶活性,促进凋落物的分解和养分释放[21]。但也有研究发现针叶树与阔叶树的凋落物混合没有明显促进凋落物分解[22]。进一步研究发现,不同树种组合和不同比例混合凋落物的分解效应存在明显差异[23-26]。因此,研究针叶树与不同阔叶树种的混合凋落物及不同混合比例对凋落物分解过程中酶活性的混合效应,对于深入研究凋落物分解机制具有重要意义。
明确混交林中各树种混合凋落物分解对混交林物质循环的影响极其重要[27,28]。一方面,混交林树种的种类和混栽比例可通过改变混交林凋落物的种类、数量和比例影响凋落物的分解速率[29];另一方面,凋落物分解又通过碳循环改变林下土壤的理化性质,进而影响森林生态系统的生产力和生物量[6]。在凋落物分解过程中,碳循环相关酶不仅调控分解速率,而且影响养分释放[30,31]。因此,探究针叶树与阔叶树混合凋落物分解过程中碳循环相关酶活性对于深入了解混合凋落物的分解机制至关重要。
在作者所在研究团队前期工作的基础上[32],本研究按照不同树种组成和混合比例,将马尾松与3种乡土阔叶树檫木〔Sassafrastzumu(Hemsl.) Hemsl.〕、香樟〔Cinnamomumcamphora(Linn.) Presl〕和香椿〔Toonasinensis(A. Juss.) Roem.〕的凋落叶进行混合,分析了分解关键期(分解75、249、437和618 d)碳循环相关酶(包括纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶)活性的变化,并对混合凋落叶分解过程中上述酶活性进行混合效应分析,对混合凋落叶初始质量[33]及pH值和含水量(另文发表)与不同分解时间上述酶活性进行多元线性逐步回归分析,以期为将马尾松纯林改造成针阔混交林时伴生树种的筛选及合适混交比例的选择提供理论依据。
1 研究区概况和研究方法
1.1 研究区概况
实验林分在四川省都江堰市四川农业大学实验基地(东经103°34′~103°36′、北纬31°01′~31°02′,海拔720~850 m)内。该基地气候类型以亚热带季风湿润气候为主,年均温15.2 ℃,极端最高温38 ℃,极端最低温-10 ℃,年均降水量1 243 mm,年均空气相对湿度81%,无霜期280 d。降水量和月均温均在每年的7月份和8月份较高,在12月份至次年2月份较低。土壤为黄壤,厚度80~110 cm,土壤酸碱度为pH 4.5至pH 5.5。区内植被类型为亚热带常绿阔叶林,植物种类较为丰富,林龄约39 a,郁闭度约0.7。乔木层主要有马尾松、杉木〔Cunninghamialanceolata(Lamb.) Hook.〕和香樟等种类;灌木层盖度约10%,主要有十大功劳〔Mahoniafortunei(Lindl.) Fedde〕、光叶海桐(PittosporumglabratumLindl.)、针刺悬钩子(RubuspungensCamb.)、野花椒(ZanthoxylumsimulansHance)、胡枝子(LespedezabicolorTurcz.)和欧洲蕨〔Pteridiumaguilinum(Linn.) Kuhn〕等种类;草本层盖度约60%,主要有麦冬〔Ophiopogonjaponicus(Linn. f.) Ker-Gawl.〕、芒萁〔Dicranopterispedata(Houtt.) Nakaike〕、山羽藓〔Abietinellaabietina(Hedw.) Fleisch.〕、香附子 (CyperusrotundusLinn.)和蝴蝶花(IrisjaponicaThunb.)等种类。
1.2 研究方法
1.2.1 实验设计 根据前人研究结果[34]及中国混交林主要树种与伴生树种的常用混交比例[35]设置马尾松与3种阔叶树凋落叶的混合比例,共设置35种凋落叶组合,包括4种单一凋落叶(质量比10∶0)和31种混合凋落叶,混合凋落叶又分成一针一阔、一针两阔和一针三阔3种混合模式,其中,一针一阔混合模式设置3个质量比,分别为8∶2、7∶3和6∶4;一针两阔混合模式设置6个质量比,分别为8∶1∶1、7∶2∶1、7∶1∶2、6∶3∶1、6∶2∶2和6∶1∶3;一针三阔混合模式设置4个质量比,分别为7∶1∶1∶1、6∶2∶1∶1、6∶1∶2∶1和6∶1∶1∶2。
于2016年6月上旬,在宜宾市高县的典型低山丘陵区收集新鲜马尾松(P)、檫木(S)、香樟(C)和香椿(T)的凋落叶,自然晾干,按照上述混合比例将不同树种的凋落叶进行混合,装入上、下层孔径分别为3.00和0.04 mm的尼龙网袋(20 cm×25 cm)中,每组混合凋落叶的总质量均为15 g。同年8月28日,在实验基地内选取3个面积30 m×30 m的典型样地(海拔、地形地貌、坡度、坡位、土壤类型和林分密度等条件基本相同),去除地表的枯枝落叶,将装有混合凋落叶的尼龙网袋平铺在地表(袋间距2~5 cm),进行野外分解实验。凋落叶分解袋共420袋。
1.2.2 样品采集与保存 在凋落叶分解关键期,即分解75、249、437和618 d,每个处理组随机选取3个尼龙网袋,置于冷藏箱中带回实验室进行分析。
1.2.3 酶活性测定 参考袁亚玲[32]和Allison等[36]的方法测定凋落叶中纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶的活性,每个指标重复测定3次。
1.2.4 混合效应分析 根据混合凋落叶分解过程中上述酶活性的相对混合效应(RME)评估混合效应类型。参照Wardle等[37]的方法计算RME值。RME值显著(P<0.05)大于0或显著小于0为非加和效应,RME值不显著大于0或不显著小于0为加和效应。并且,RME值显著大于0为是协同效应;RME值显著小于0为拮抗效应。
1.3 数据处理及统计分析
采用EXCEL 2021和SPSS 20.0软件进行数据整理、计算和分析,采用单因素方差分析法和邓肯氏新复极差法对不同分解时间酶活性的差异进行分析,并采用重复测量方差分析(repeated measures analysis of variance)对混合凋落叶酶活性在不同树种组成和质量比间的差异进行分析。基于各混合凋落叶酶活性的RME值,采用配对t检验法进行混合效应分析。根据混合凋落叶的初始质量[33]及pH值和含水量,以碳含量为x1、氮含量为x2、磷含量为x3、木质素含量为x4、纤维素含量为x5、总酚含量为x6、缩合单宁含量为x7、碳含量与氮含量的比值为x8、木质素含量与氮含量的比值为x9、微生物量碳为x10、微生物量氮为x11、pH值为x12、含水量为x13,以混合凋落叶的4种酶活性为y,进行多元线性逐步回归分析。
2 结果和分析
2.1 不同凋落叶分解过程中碳循环相关酶活性差异
2.1.1 不同分解时间的差异 马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性的比较结果见表1。由表1可见:随着分解时间延长,单一马尾松凋落叶的纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶活性均先降低后升高,且均在分解75 d最高、在分解249 d最低。3种单一阔叶树凋落叶的4种酶活性变化各异。纤维二糖水解酶活性在单一檫木凋落叶中先降低后升高,在分解437 d最低、在分解618 d最高;在单一香樟凋落叶中持续升高;在单一香椿凋落叶中先升高后降低再升高,在分解437 d最低、在分解618 d最高。β-葡萄糖苷酶活性在3种单一阔叶树凋落叶中均先降低后升高,且均在分解437 d最低、在分解618 d最高。多酚氧化酶活性在3种单一阔叶树凋落叶中也先降低后升高,且均在分解75 d最高、在分解249 d最低。过氧化物酶活性在3种单一阔叶树凋落叶中总体上表现为先降低后升高并趋于稳定,且在单一檫木凋落叶和单一香椿凋落叶分解75 d最高、分解249 d最低,而在单一香樟凋落叶分解437 d最高、分解249 d最低。除单一香椿凋落叶的过氧化物酶活性外,3种单一阔叶树凋落叶的纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶活性的最高值与最低值的差异达到显著水平。

表1 马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性的比较
总体来看,随着分解时间延长,马尾松和香樟混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性持续升高,各混合凋落叶的过氧化物酶活性先降低后升高再降低,而各混合凋落叶的其余酶活性却先降低后升高。马尾松和香椿混合凋落叶(质量比6∶4),马尾松和檫木混合凋落叶(质量比7∶3和6∶4),马尾松、檫木和香椿混合凋落叶(质量比8∶1∶1、7∶2∶1、7∶1∶2、6∶3∶1、6∶2∶2和6∶1∶3),马尾松、檫木和香樟混合凋落叶(质量比7∶2∶1、6∶3∶1和6∶2∶2),马尾松、香樟和香椿混合凋落叶(质量比6∶3∶1和6∶1∶3)以及马尾松、檫木、香樟和香椿混合凋落叶(质量比7∶1∶1∶1、6∶2∶1∶1、6∶1∶2∶1和6∶1∶1∶2)的纤维二糖水解酶活性在分解75 d最高;其余混合凋落叶的纤维二糖水解酶活性基本在分解437或618 d最高;多数混合凋落叶的纤维二糖水解酶活性在分解249 d最低。马尾松和香椿混合凋落叶,马尾松、檫木和香椿混合凋落叶,马尾松、檫木和香樟混合凋落叶,马尾松、香樟和香椿混合凋落叶以及马尾松、檫木、香樟和香椿混合凋落叶的β-葡萄糖苷酶活性基本在分解75 d最高;其余混合凋落叶的β-葡萄糖苷酶活性在分解437或618 d最高;多数混合凋落叶的β-葡萄糖苷酶活性在分解249或437 d最低。马尾松和香椿混合凋落叶(质量比8∶2和6∶4),马尾松、檫木和香椿混合凋落叶(质量比8∶1∶1、6∶3∶1、6∶2∶2和6∶1∶3),马尾松、檫木和香樟混合凋落叶(质量比8∶1∶1和7∶2∶1),马尾松、香樟和香椿混合凋落叶(质量比7∶2∶1、6∶3∶1和6∶2∶2)以及马尾松、檫木、香樟和香椿混合凋落叶(质量比7∶1∶1∶1、6∶1∶2∶1和6∶1∶1∶2)的多酚氧化酶活性在分解75 d最高;其余混合凋落叶的多酚氧化酶活性基本在分解437或618 d最高;多数混合凋落叶的多酚氧化酶活性在分解249 d最低。除马尾松、檫木、香樟和香椿混合凋落叶(质量比6∶1∶2∶1)外,其余混合凋落叶的过氧化物酶均在分解437或618 d最高;多数混合凋落叶的过氧化物酶活性在分解249 d最低。并且,各混合凋落叶的纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶活性的最高值与最低值的差异基本上达到显著水平。
2.1.2 不同树种组成间的差异 基于表1的结果,对混合凋落叶分解过程中碳循环相关酶活性在不同树种组成间的差异进行统计,结果见表2。由表2可见:纤维二糖水解酶活性在马尾松、檫木、香樟和香椿混合凋落叶中最高,β-葡萄糖苷酶活性在马尾松和香椿混合凋落叶中最高,多酚氧化酶活性在马尾松、檫木和香樟混合凋落叶中最高,而过氧化物酶活性在马尾松和檫木混合凋落叶中最高。4种酶活性在单一马尾松凋落叶中较高,但在3种单一阔叶树凋落叶中较低。在2个树种的混合凋落叶中,马尾松和香椿混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性最高,多酚氧化酶和过氧化物酶活性最低。在3个树种的混合凋落叶中,马尾松、檫木和香樟混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性最低,但多酚氧化酶和过氧化物酶活性最高。马尾松、檫木和香椿混合凋落叶,马尾松、香樟和香椿混合凋落叶以及马尾松、檫木、香樟和香椿混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性高于单一马尾松凋落叶;马尾松和香椿混合凋落叶的纤维二糖水解酶活性低于单一马尾松凋落叶,但β-葡萄糖苷酶活性却高于单一马尾松凋落叶;其余混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性低于单一马尾松凋落叶。除马尾松、檫木和香樟混合凋落叶的多酚氧化酶活性以及马尾松和檫木混合凋落叶的过氧化物酶活性外,其余树种组成的混合凋落叶的多酚氧化酶和过氧化物酶活性低于单一马尾松凋落叶。
表2 马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性在不同树种组成间的差异
2.1.3 不同质量比间的差异 基于表1的结果,对混合凋落叶分解过程中碳循环相关酶活性在不同质量比间的差异进行统计,结果见表3。由表3可见:纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶活性分别在质量比6∶1∶1∶2、6∶1∶2∶1、6∶3∶1和8∶2的马尾松和阔叶树混合凋落叶中最高。除质量比6∶1∶2∶1的马尾松和阔叶树混合凋落叶的多酚氧化酶活性低于单一凋落叶外,其余马尾松和阔叶树凋落叶质量比混合凋落叶的4种酶活性均高于单一凋落叶。
表3 马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性在不同质量比间的差异
2.2 混合凋落叶分解过程中碳循环相关酶活性的混合效应
从马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性的相对混合效应(RME)统计结果(表4)可见:实验期间,马尾松和3种阔叶树混合凋落叶的纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶活性以加和效应为主,尤其是在分解437和618 d,仅少部分混合凋落叶的酶活性表现为非加和效应。在分解75、249、437和618 d,纤维二糖水解酶活性表现为非加和效应的混合凋落叶分别有11、8、1和4种,其中,表现为拮抗效应的混合凋落叶分别有10、1、1和4种,表现为协同效应的混合凋落叶分别有1、7、0和0种;β-葡萄糖苷酶活性表现为非加和效应的混合凋落叶分别有10、10、6和2种,其中,表现为拮抗效应的混合凋落叶分别有6、2、1和2种,表现为协同效应的混合凋落叶分别有4、8、5和0种;多酚氧化酶活性表现为非加和效应的混合凋落叶分别有22、10、1和3种,其中,表现为拮抗效应的混合凋落叶分别有22、1、1和1种,表现为协同效应的混合凋落叶分别有0、9、0和2种;过氧化物酶活性表现为非加和效应的混合凋落叶分别有19、7、3和4种,其中,表现为拮抗效应的混合凋落叶分别有19、1、0和3种,表现为协同效应的混合凋落叶分别有0、6、3和1种。
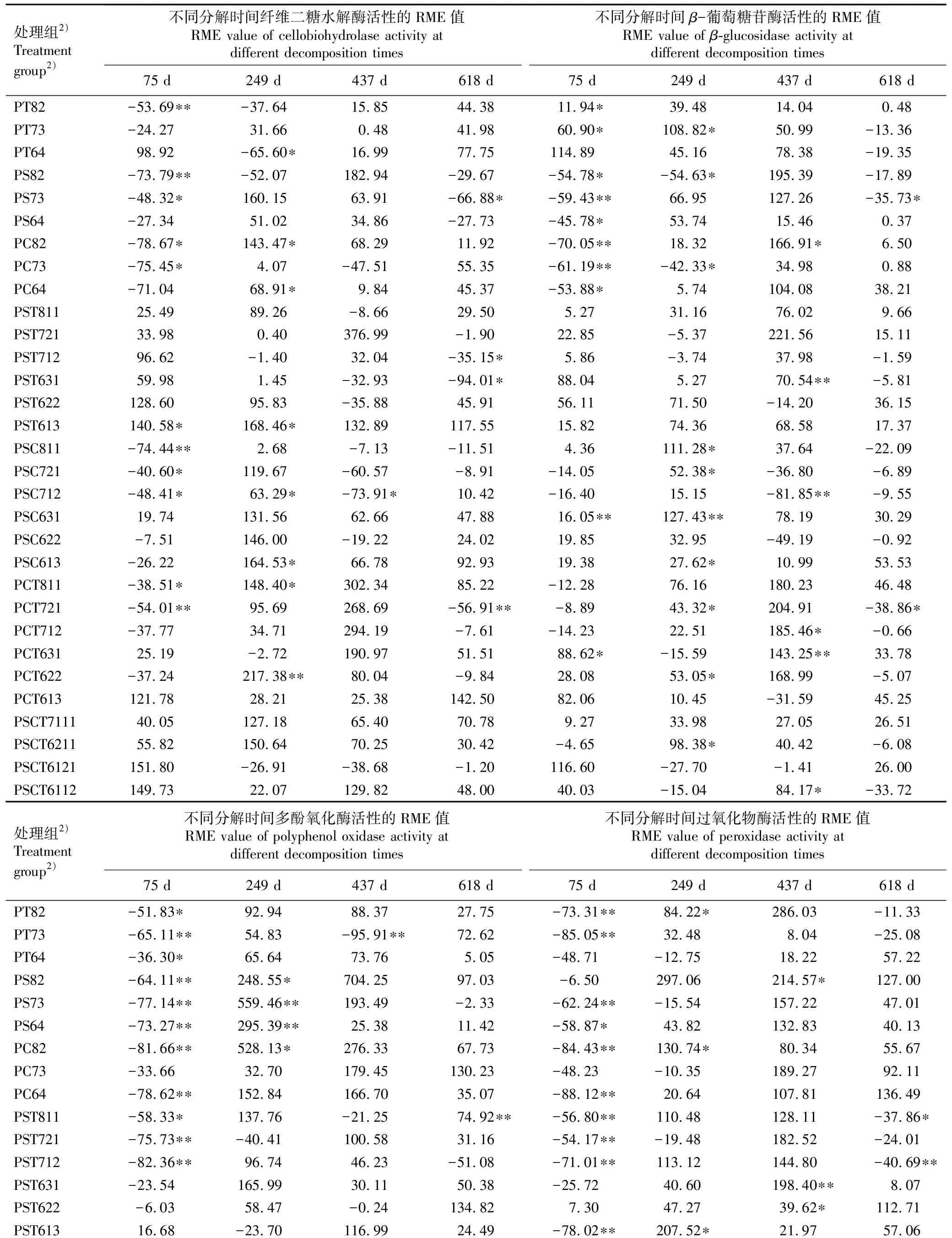
表4 马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性的相对混合效应(RME)分析1)
2.3 混合凋落叶分解过程中酶活性的多元线性逐步回归分析
多元线性逐步回归分析结果(表5)表明:马尾松与3种阔叶树混合凋落叶的初始质量及pH值和含水量对不同分解时间纤维二糖水解酶、β-葡萄糖苷酶、多酚氧化酶和过氧化物酶活性的影响存在明显差异,部分因子在不同分解时间对同一酶活性的影响完全相反。混合凋落叶分解过程中,碳含量、纤维素含量、总酚含量、缩合单宁含量和pH值抑制纤维二糖水解酶活性,而磷含量和含水量则促进纤维二糖水解酶活性;碳含量、纤维素含量和缩合单宁含量抑制β-葡萄糖苷酶活性,而木质素含量、微生物量碳、微生物量氮和含水量则促进β-葡萄糖苷酶活性;磷含量、缩合单宁含量和pH值抑制多酚氧化酶活性,而碳含量、木质素含量、纤维素含量、缩合单宁含量、碳含量与氮含量的比值、微生物量碳和含水量则促进多酚氧化酶活性;磷含量、pH值和含水量抑制过氧化物酶活性,而磷含量、木质素含量、纤维素含量、总酚含量、缩合单宁含量、微生物量碳和含水量则促进过氧化物酶活性。整个分解过程中,混合凋落叶的纤维二糖水解酶活性受到碳含量的抑制作用以及磷含量和微生物量碳的促进作用,β-葡萄糖苷酶活性受到缩合单宁含量和微生物量碳的抑制作用以及磷含量的促进作用,多酚氧化酶活性受到磷含量的抑制作用和微生物量碳的促进作用,过氧化物酶活性仅受到缩合单宁含量的促进作用。

表5 马尾松与3种阔叶树混合凋落叶分解过程中碳循环相关酶活性的多元线性逐步回归分析1)
3 讨论和结论
3.1 混合凋落叶分解过程中碳循环相关酶活性变化
马尾松针叶的木质素含量较高且比叶面积较小[38],不利于微生物的生长和繁殖,林下凋落物的分解能力相对较差,而阔叶树的叶片较大且质地柔软,容易被生活在土壤中的动物咀嚼和粉碎,有助于微生物的生长和繁殖。此外,针叶树通常具有一些化感物质,将针叶树凋落物与阔叶树凋落物混合不但会降低单位混合凋落物的化感物质浓度,而且能够为微生物提供营养成分[39],提高相关酶的活性,有利于凋落物的分解。本研究结果表明:总体来看,随着分解时间延长,混合凋落叶的过氧化物酶活性表现为先降低后升高再降低的变化趋势,混合凋落叶及单一凋落叶的纤维二糖水解酶、β-葡萄糖苷酶和多酚氧化酶活性则呈先降低后升高的变化趋势。过氧化物酶活性基本在混合凋落叶分解437或618 d达到最高值,其余3种酶活性则基本在混合凋落叶分解75 d达到最高值。在分解早期,为了获得更多的养分,凋落叶中的微生物大量分泌纤维二糖水解酶和β-葡萄糖苷酶,使凋落叶中的易降解成分大量释放,致使2种酶活性较高;随着分解进程的持续,凋落叶中的难降解成分(木质素等)占比逐渐升高,纤维二糖水解酶和β-葡萄糖苷酶活性随之逐渐降低;然而,随着分解进程的继续,凋落叶的组织结构发生很大变化,部分难降解成分逐步转化为易降解成分[6],同时因马尾松凋落叶在分解后期出现纤维素分解菌群诱集过程[40],最终导致纤维二糖水解酶和β-葡萄糖苷酶活性再次升高。多酚氧化酶和过氧化物酶主要参与凋落物中难降解碳的分解过程[19,41,42],因此,在马尾松与3种阔叶树混合凋落叶中难降解物质较多时(即分解后期),过氧化物酶和多酚氧化酶活性显著升高。然而,分解后期凋落叶基质质量降低,导致参与凋落叶分解的土壤动物数量下降,同时,分解后期较低的养分限制微生物分泌过氧化物酶,从而导致其活性降低。
值得注意的是,在分解249~618 d,马尾松和香樟混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性持续升高,推测造成此结果的原因是适量的香樟凋落叶为微生物的生长繁殖提供了稳定的环境[43],并且,随着分解进程的持续,香樟叶片中养分的可利用性通过淋溶作用和土壤生物破碎作用被逐渐提高,这些均有利于混合凋落叶中纤维二糖水解酶和β-葡萄糖苷酶活性升高。
3.2 树种组成对混合凋落叶分解过程中碳循环相关酶活性的影响
凋落物分解可调控森林生态系统的物质循环和能量流动,酶在凋落物分解过程中发挥着重要的调控作用[23]。与单一马尾松凋落叶相比,3种阔叶树的单一凋落叶碳循环相关酶活性显著降低,马尾松与3种阔叶树的混合凋落叶碳循环相关酶活性未显著升高。由于单一马尾松凋落叶的初始碳含量最高,且凋落物中不稳定和易被利用的含碳化合物可提高分解难降解化合物的酶活性[10],导致单一马尾松凋落叶的碳循环相关酶活性高于单一阔叶树凋落叶。马尾松凋落叶与阔叶树凋落叶混合后,由于阔叶树凋落叶中含有更多的可溶性碳,导致混合凋落叶中碳的可利用性发生改变,进而提高相关酶的活性。本研究结果表明:不同树种组成对混合凋落叶的碳循环相关酶活性有较大影响,例如:马尾松和香椿混合凋落叶的纤维二糖水解酶和β-葡萄糖苷酶活性高于马尾松和檫木混合凋落叶以及马尾松和香樟混合凋落叶,而马尾松、檫木和香椿混合凋落叶以及马尾松、香樟和香椿混合凋落叶的这2种酶活性则高于马尾松、檫木和香樟混合凋落叶;多酚氧化酶和过氧化物酶活性则恰好相反。纤维二糖水解酶活性在马尾松、檫木、香樟和香椿混合凋落叶中最高,β-葡萄糖苷酶活性在马尾松和香椿混合凋落叶中最高,多酚氧化酶活性在马尾松、檫木和香樟混合凋落叶中最高,而过氧化物酶活性在马尾松和檫木混合凋落叶中最高。可见,马尾松与3种阔叶树混合凋落叶的碳循环相关酶活性虽然没有较单一马尾松凋落叶显著提高,但混合香椿凋落叶有利于纤维二糖水解酶和β-葡萄糖苷酶活性提高,而混合檫木和香樟凋落叶有利于多酚氧化酶和过氧化物酶活性提高。这主要归因于混合凋落叶的自身特性:香椿凋落叶的养分含量最高,碳含量与氮含量的比值和难降解成分含量最低[38],可为微生物提供更多可利用的养分,有利于碳循环相关酶活性的提高[44,45];香樟凋落叶厚且革质,不利于土壤动物破碎及微生物的侵入和定植[46],而檫木凋落叶的木质素含量较高,可抑制降解易分解成分的酶活性,而且难降解成分可与蛋白质结合形成络合物,致使碳循环相关酶活性降低[40]。
多元线性逐步回归分析结果表明:凋落叶中的难分解物质(碳含量、纤维素含量和缩合单宁含量等)可抑制纤维二糖水解酶和β-葡萄糖苷酶活性,并促进多酚氧化酶和过氧化物酶活性。混合效应分析结果表明:分解过程中,混合凋落叶的4种酶活性主要表现为加和效应,少数混合凋落叶的酶活性表现为非加和效应,并且,仅少部分混合凋落叶的酶活性在分解75 d表现为拮抗效应,其中,多数混合凋落叶的多酚氧化酶和过氧化物酶活性在分解75 d表现为拮抗效应;分解249 d,酶活性表现为协同效应的混合凋落叶增多,并以混有香椿和香樟的混合凋落叶为主。研究发现:马尾松含有较多的树脂和木质素等较难分解的成分[38],且在分解后会产生酸性成分,抑制微生物的捕食和繁殖等活动[47],这可能是部分混合凋落叶的酶活性在分解75 d表现为拮抗效应的主要原因。随着分解时间的延续,阔叶树凋落叶中的养分不断释放,有利于微生物生长,特别是混有香椿凋落叶的混合凋落叶最为明显,而香樟凋落叶的养分可利用性在淋溶作用及土壤动物破碎作用下逐渐提高,以致混有香椿和香樟的部分混合凋落叶的酶活性表现为协同效应。可见,凋落叶自身的化学和物理性质相互作用,共同影响混合凋落叶分解过程中的酶活性[48,49]。
3.3 凋落叶质量比对混合凋落叶分解过程中碳循环相关酶活性的影响
凋落叶的混合比例对混合凋落物的酶活性有一定的影响,只有凋落叶混合比例合适才能显著促进混合凋落叶的分解[49]。相关研究结果表明:碳循环相关酶主要参与易分解有机质的分解过程,这些酶是凋落物分解初期的重要指标[31]。总体来看,除过氧化物酶活性在质量比8∶2的马尾松和阔叶树混合凋落叶中较高外,其余3种酶活性均在质量比6∶4的马尾松和阔叶树混合凋落叶中较高。这是因为阔叶树凋落叶占比较大的混合凋落叶可为微生物生长和繁殖提供更多可利用的养分,酶活性相应升高。廖利平等[22]的研究结果也表明:阔叶树凋落叶占比较大(30%以上)的混合凋落叶的分解速率明显加快,而针叶树凋落叶占比较大(针叶树凋落叶与阔叶树凋落叶质量比9∶1和8∶2)的混合凋落叶的分解速率加快不明显。林开敏等[18]在研究杉木与楠木(PhoebezhennanS. Lee et F. N. Wei)混合凋落叶分解时发现楠木凋落叶占比越大,混合凋落叶中N和K的释放越快。Zhang等[50]研究马尾松与阔叶树凋落叶混合分解过程中真菌多样性时发现,香椿凋落叶占比达30%~40%的混合凋落叶的真菌多样性提高。由此可见,针叶树和阔叶树凋落叶的混合比例对混合凋落叶的分解具有重要的调控作用,阔叶树凋落叶占比增加更有利于混合凋落叶的分解和酶活性的提高。
3.4 结论
综上所述,虽然马尾松与3种阔叶树凋落叶混合后没有显著促进碳循环相关酶活性,但酶活性在混合凋落叶分解过程中以及不同树种组成和质量比的混合凋落叶中均存在一定差异。总体来看,随着分解时间延长,混合凋落叶的纤维二糖水解酶、β-葡萄糖苷酶和多酚氧化酶活性先降低后升高,而过氧化物酶活性则先降低后升高再降低。混合香椿凋落叶有利于混合凋落叶中纤维二糖水解酶和β-葡萄糖苷酶活性的提高,而混合檫木和香樟凋落叶有利于多酚氧化酶和过氧化物酶活性的提高。马尾松与阔叶树凋落叶质量比6∶4有利于混合凋落叶中碳循环相关酶活性的提高。
