中国肉盘衣属地衣新记录种
2022-02-25朱孟丽赵欣王伟成贾泽峰
朱孟丽, 赵欣, 王伟成, 贾泽峰*
中国肉盘衣属地衣新记录种
朱孟丽1*, 赵欣1*, 王伟成2,3, 贾泽峰1**
(1.聊城大学生命科学学院,山东 聊城 252059;2.中国科学院微生物研究所,北京 100101;3.中国科学院大学,北京 100049)
在对海南省叶生地衣资源的调查研究中发现了中国肉盘衣属地衣新记录种:乳头肉盘衣[(Vain.) Lücking & Sérus.],描述了其形态学、解剖学及化学特征,提供了显微结构照片。主要识别特征为:地衣体壳状,布满灰绿色乳头状颗粒,子囊盘棕色,囊盘被外部由疏丝组织构成,子囊孢子(3~)7隔,26~36m×2~3m,呈末端变窄的细棍棒状。基于线粒体小亚基(mtSSU)序列构建的系统发育树表明该种与本属模式种(Müll.Arg.) Vězda具有较近的亲缘关系。编制了中国该属地衣的分种检索表。
乳头肉盘衣;茶渍纲;地衣型真菌;mtSSU;分类学;新记录
The genus, belonging to family Pilocarpaceae, in order, class Lecanoro- mycetes was established in 1986 with its type(Müll.Arg.) Vězda[1-2], including more than 90 species worldwide[3-4].is the second largest genus among foliicolous lichens next toAcharius[5]and quite diverse in morphological fea- tures[6].Thedistinctive traits of this genus are the comparatively small apothecia with a thin margin; paraplectenchymatous excipulum; usually indistinct, branched, and sparsely to densely anastomosing para- physes;-type asci; ellipsoid to cylindrical, transversely septate to muriform ascospores; and pycnidial conidiomata[6].are mostly folii- colous, common in pan-tropical ecologies.
China had previously reported ninespecies, which are distributed in Yunnan, Hainan, Hongkong and Taiwan[7-13].Based on specimens collected from Hainan Province,(Vain.) Lücking & Sérus.is found new to China.
1 Materials and methods
1.1 Specimens and morphology
The specimens of new record are deposited in the Fungarium of College of Life Sciences, Liaocheng University (LCUF) and Herbarium Mycologicum Aca- demiae Sinicae-Lichenes (HMAS-L).A dissecting microscope (Olympus SZX16) and a light microscope (Olympus BX53) were used for the morphological and anatomical studies.Measurements were taken from mature vertical sections of fruit bodies mounted in water.
1.2 Chemistry
Amyloidity of the ascospores was tested using Lugol’s solution.Spot tests with K (10% aqueous solution of potassium hydroxide), C (saturated solution of aqueous sodium hypochlorite), and P (saturated solution of-phenylenediamine in 95% ethyl alcohol) were performed on the thallus surface.The lichen substances were detected and identified by thin-layer chromatography, using solvent C[14-16].
1.3 DNA extraction, amplification, and sequencing
Genomic DNA was extractedfrom ascomata of the specimens using the Hi-DNAsecure Plant Kit (Tiangen, Beijing, China) according tothe manufac- turer’s protocol.PCR amplification was performed using the mtSSU1 and mtSSU3R primer pair for mtSSU[17].The 25L PCR reaction system containing 1L each primer solution (10mol/L), 0.5Lgenomic DNA, 10L ddH2O, and 12.5L2×PCR Master- Mix (Tiangen, Beijing, China).Thermocyclingcondi- tions comprised initial denaturation at 95℃(5 min); 35 denaturation cycles at 94 ℃(45 s), annealing at 50 ℃(1 min), extension at 72 ℃(1.5 min) and a final extension at 72 ℃(10 min).The target product of PCR was affirmed by electrophoresis on 1% agarose gels and sequenced by Biosune Inc.(Shanghai).Nine newly generated sequences were submitted to GenBank.The sequences obtained were evaluated using BLASTn and combined with selected sequences of Pilocarpaceae from GenBank (Fig.1),andwere used as the outgroup based on Ekman et al.[18]
1.4 Phylogenetic analysis
Contigs were assembled and edited using the program Geneious v.6.1.2 (Biomatters Ltd., Auckland, NZ).A total of 39 sequences were aligned using MAFFT v.7[19].The program Gblocks v.0.91b was used to delimit ambiguous regions, implementing all the options for a less stringent selection (http://molevol.cmima.csic.es/castresana/Gblocks_server.html)[20], which yielded final alignment of 670 bp.Maximum likely- hood (ML) and Bayesian inference (BI) were per- formed using the CIPRES Scientific gateway portal (http://www.phylo.org/portal2/)[21].Maximum likelihood bootstrapping analysis was performed with RAxML- HPC v.8[22], using the default parameters as imple- mented on the CIPRES, NSF XSEDE resource with bootstrap statistics calculated from 1 000 bootstrap replicates.For the Bayesian analysis, the best substi- tution model was estimated using jModelTest 2.1.6[23].Based on the results, we used GTR+I+G model.Baye- sian analysis was performed using MrBayes v.3.2.2 on CIPRES with 2 independent runs, searching for 10 000 000 generations with four independent chains and sampling every 1000th tree[24].After discarding the burn-in, the remaining 7 500 trees of each run werepooled to calculate a 50% majority rule consensus tree.Generated phylogenetic tree was visualized under Figtree v.1.4.2[25].
2 Results
2.1 Phylogenetic analysis
The final alignment consisted of 9 newly generated mtSSU sequences and 30 sequences downloaded from NCBI (Fig.1).The phylogenetic trees obtained from maximum likelihood (ML) and Bayesian inference analysis (BI) exhibited the same topology; we there- fore present only the ML tree.The molecular phylo- geny based on the mitochondrial small subunit marker (mtSSU) of Pilocarpaceae exhibits a well-supported monophyletic lineage containing the genera,,,,,and.The tree showsis polyphyletic in its current delimitation.is revealed as a sister clade to the type speciesThese two species together with,andform a monophyletic lineage without good support (BS=59%, PP=0.85).,,,andcluster together and get a high support (BS=96%, PP=1.0).While anotherspecies,shows a close relationship with.
2.2 Taxonomy
(Vain.) Lücking & Sérus, in Lichenologist 33(3): 192 (2001) Fig.2

Fig.1 Maximum likelihood tree of Fellhanera mastothallina and related species within Pilocarpaceae based on the mitochondrial small subunit marker (mtSSU).ML bootstrap values and MCMC posterior probabilities (second value) are displayed above each branch.Branches recovered with ML-BS support ≥70% and BI-PP support ≥0.95 were regarded as strongly supported.GenBank accessions are attached to the sequences.Newly generated sequences are shown in bold.
≡Vain., in Ann.Acad.Sci.Fenn., ser.A, 15: 64 (1921)
≡(Vain.) Vězda, in Vězda et al., Ann.Naturh.Mus.Wien 99B: 738 (1997)
Type: Philippines, Robinson & Ramos 11900 (TUR-holotype).
Description:Thallus foliicolous, crustose, conti- nuous, verrucose, 10-40 mm across and 10-15m thick, greyish-green, rough, often irregular in outline, with light green to orange-red verrucosa of 0.01-0.03 mm diam.Apothecia when mature sessile, rounded, 0.2-0.5 mm diam.and 150-200m high; disc plane to slightly convex, brown to dark brown; margin distinct and thin, about 0.1 mm wide, usually accom- panied by a white pruina.Excipulum light grey to light brown, 7-15m thick, internal parts appear paraplectenchymatuous and external parts prosoplec- tenchymatous.Hymenium 40-66m high, colourless.Hypothecium 26-74m high, brown to dark brown.Asci clavate, 36-48m×4-9m.Ascospores 8 per ascus, narrowly clavate, and tapering towards pro- ximal end, (3-)7-septate, 26-36m×2-3m, about 10-13 times as long as broad; I+ violet.Pycnidia not observed.
Chemistry: Trace amounts of substances were detected by TLC.Spot tests on thallus: K+ dark brown, C-, P-, KC+ dark brown.
Ecology and distribution: The species is a typi- cally foliicolous lichen, grows mainly in tropical Asia and also reported in Papua New Guinea, Australia and New Caledonia[26].New to China.
Specimen examined: CHINA.Hainan: WuzhishanCity, Wuzhishan Nature Reserve, on leaves, 18°54′27″ N, 109°40′48″ E, elev.730 m, 12 Dec.2019,Y.H.JuHN19446-a(GB accession No.: MW045559),HN19458(GB accession No.: MW045560),HN19459 (LCUF).Changjiang County, Bawangling Nature Reserve, Bai- shitan Scenic Area, on leaves, 19°7′17″ N, 109°4′53″E, elev.700 m, 4 Sep, 2017,W.C.Wang HN20170025 (HMAS-L 139457, GB accession No.: MW553282); Yajia Scenic Area, on leaves, 19º7′17″N, 109º4′53″, elev.550 m, 5 Sep.2017.W.C.WangHN20170124(HMAS-L 139601, GB accession No.: MW553281).
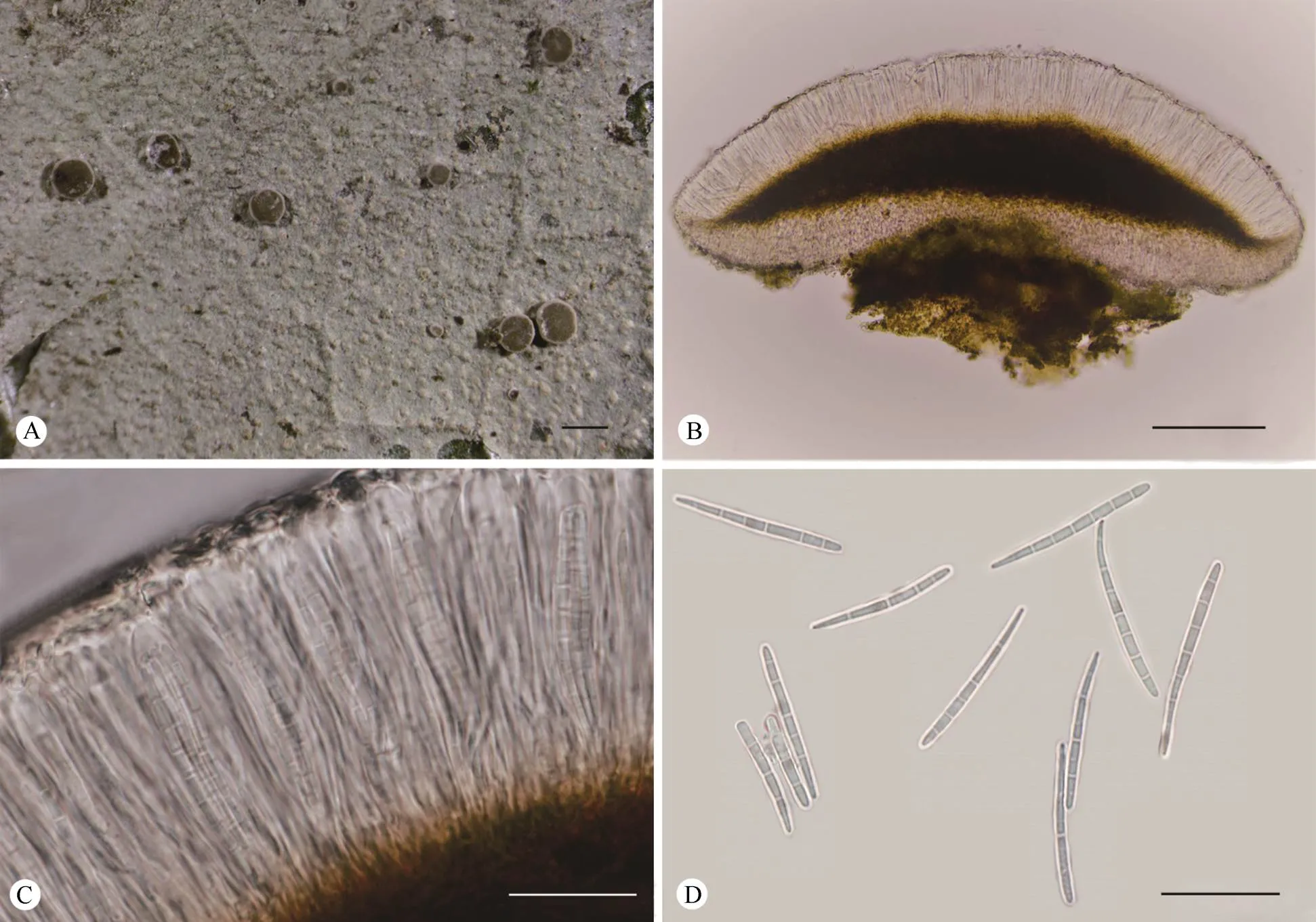
Fig.2 Fellhanera mastothallina (Y.H.Ju HN19458).A: Thallus with apothecia; B: Vertical section of an apothecium; C: Part of a vertical section of an apothecium; D: Ascospores.Bars: A=0.5 mm, B=100 µm, C-D=20 µm
3 Discussion
Phylogeneticallyis close related to the type speciesthis result confirms the hypothesis thatbelonged togroup by Lücking based on evident from thallus and apothecial characters[6].can be distinguished by reddish brown apothecia, paraplectenchymatous exciple and shorter ascospores (18-24m×3-4.5m).The mor- phological characteristics of our specimens collected from Hainan are almost identical with the type speci- men from Phillippines and materials from Papua New Guinea and New Caledonia except for the longer and narrower ascospores (the latter are 22-32m×3- 4m)[26-27].Our phylogenetic result thatbeing heterogeneous in its current circumscription is coincident with the preliminary studies of the family Pilocarpaceae[13,28-29].Some taxa withincan be placed in other genera once sufficient evidence obtained in the future molecular studies with larger taxon sampling.Comparisons of the characteristics of the known Chinese species ofare shown in Table 1.

Table 1 Comparisons of the characteristics of the known Chinese species of Fellhanera
TLocalities newly recorded are shown in boldface.
Key to the known Chinese species of
1a.Lichen compounds present, mainly usnic, isousnic and zeorin acids………………………………2
1b.Lichen compounds absent………………………………………………………………3
2a.Ascospores 1-septate, 10-17m×3-6m, 2.5-3.5 times as long as broad………………………………
2b.Ascospores 3(-4)-septate, 10-16m×3-4.5m, 3-4 times as long as broad………………………………
3a.Ascospores l-septate or non-septate………………………………………………………………4
3b.Ascospores more than 1-septate………………………………………………………………5
4a.Ascospores oblong-ovoid, 1-septate, with constriction at septa, 10-16m×4-5m, 2.5-3.5 times as long as broad………………………………
4b.Ascospores elongate ellipsoid, (0-)l-septate, without constriction at septa, (12-)14-17m×3-5.5m, 3-4 times as long as broad………………………………………………………………
5a.Ascospores only 3-septate………………………………………………………………6
5b.Ascospores 5 or 7-septate, rarely 3-septate………………………………………………………………7
6a.Disc orange; ascospores 11-16m×2.5-4.5m, 3.5-4.5 times as long as broad………………………………
6b.Disc reddish brown to dark greyish brown; ascospores 12-18m×3-4m, 4-5 times as long as broad………………………………
7a.Ascospores 5-septate, rarely 3-septate………………………………………………………………8
7b.Ascospores 7-septate, rarely 3 or 5-septate………………………………………………………………9
8a.Ascospores ellipsoid, usually slightly curved and attenuated at one end, 14-21m×3.5-5, 3.5-4.5 times as long as broad
………………………………………………………………
8b.Ascospores oblong, with slight constrictions at septa, 14-26m×3-5, 4-6 times as long as broad………………………………
9a.Ascospores oblong, 7-septate, with constrictions at septa, 18- 24m×3-4.5, 5-6 times as long as broad………………………………
9b.Ascospores clavate, (3-)7-septate, rarely 3 or 5-septate, without constriction at septa, 26-36m×2-3, 10-13 times
as long as broad………………………………
[1] Vězda A.Neue gattungen der familie Lecideaceae s.lat.(Lichenes) [J].Folia Geobot Phytotax, Praha, 1986, 21(2): 199-219.
[2] LÜCKING R, HODLKINSON B P, LEAVITT S D.The 2016 classi- fication of lichenized fungi in the Ascomycota and Basidiomycota- approaching one thousand genera [J].Bryologist, 2017,119(4): 361- 416.doi: 10.1639/0007-2745-119.4.361.
[3] KONDRATYUK S Y, LŐKÖS L, TSCHABANENKO S, et al.New and noteworthy lichen-forming and lichenicolous fungi [J].Acta Bot Hung, 2013, 55(3/4): 275-349.doi: https://doi.org/10.1556/abot.55.2013.3-4.9.
[4] WIJAYAWARDENE N N, HYDE K D, KUNHIRAMAN C R, et al.Notes for genera: Ascomycota [J].Fung Diver, 2017, 86(1): 1-594.doi: 10.1007/s13225-017-0386-0.
[5] ACHARIUS E.Förteckning pa de i Sverige växande arter af Lafvarnes famille 4 [J].Kongl Vetensk Acad Nya Handl Ser.2, 1809, 30(3): 145- 169.
[6] LÜCKING R.Foliicolous lichenized fungi [J].Flora Neotrop, 2008, 103: 1-866.
[7] THROWER S L.Hong Kong Lichens [M].Hong Kong: The Urban Council, 1988: 1-193.
[8] WEI J C, JIANG Y M.Some foliicolous lichens in Xishuangbanna, China [C]Galloway D.Systematics, Conservation and Ecology of Tropical Lichens.Systematics Association Special Volume 42.Oxford: Clarendon Press, 1991: 201-216.
[9] APTROOT A, SEAWARD M R D.Annotated checklist of Hongkong lichens [J].Trop Bryol, 1999, 17(1): 57-101.doi: 10.11646/bde.17.1.12.
[10] APTROOT A, SIPMAN H J M.New Hong Kong lichens, ascomycetes and lichenicolous fungi [J].J Hatt Bot Lab, 2001, 91: 317-343.
[11] APTROOT A, SPARRIUS L B.New microlichens from Taiwan [J].Fung Diver, 2003, 14: 1-50.
[12] APTROOT A, FERRARO L I, SIMPMAN H J M., et al.Foliicolous lichens and their lichenicolous ascomycetes from Yunnan and Taiwan [J].Mycotaxon, 2003,88: 41-47.
[13] WANG W C, SANGVICHIEN E, WEI T Z, et al.A molecular phylo- geny of Pilocarpaceae Zahlbr., including a new species ofMüll.Arg.and new records of foliicolous lichenized fungi from Thai- land [J].Lichenologist, 2020, 52(5): 377-385.doi: 10.1017/S002428 2920000328.
[14] CULBERSON C F, KRISTINSSON H.A standardized method for the identification of lichen products [J].J Chromatogr, 1970, 46: 85-93.
[15] CULBERSON C F.Improved conditions and new data for the identi- fication of lichen products by a standardized thin-layer chromate- graphic method [J].J Chromatogr, 197272(1): 113-125.
[16] JIA Z F, WEI J C.Flora Lichenum Sinicorum, Vol.13, Ostropales (I), Graphidaceae 1 [M].Beijing: Science Press, 2016: 1-210.(in Chinese)
[17] ZOLLER S, SCHEIDEGGER C, SPERISEN C.PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen- forming ascomycetes [J].Lichenologist, 1999, 31(5): 511-516.
[18] EKMAN S, ANDERSEN H L, WEDIN M.The Limitations of ancestral state reconstruction and the evolution of the ascus in the Lecanorales (Lichenized Ascomycota) [J].Syst Biol, 2008, 57(1): 141- 156.doi: 10.1080/10635150801910451.
[19] KATOH K, STANDLEY D M.MAFFT multiple sequence alignment software version 7: Improvements in performance and usability [J].Mol Biol Evol, 2013, 30(4): 772-780.doi: 10.1093/molbev/mst010.
[20] CASTRESANA J.Selection of conserved blocks from multiple align- ments for their use in phylogenetic analysis [J].Mol Biol Evol, 2000, 17(4): 540-552.doi: 10.1093/oxfordjournals.molbev.a026334.
[21] MILLER M A, PFEIFFER W, SCHWARTZ T.Creating the CIPRES Science Gateway for inference of large phylogenetic trees [C]// Proceedings of the Gateway Computing Environments Workshop (Gce).LA: New Orleans, 2010: 1-8.
[22] STAMATAKIS A.RAxML Version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies [J].Bioinformatics, 2014, 30(9): 1312-1313.doi: 10.1093/bioinformatics/btu033.
[23] DARRIBA D, TABOADA G L, DOALLO R, et al.jModelTest 2: More models, new heuristics and parallel computing [J].Nat Methods, 2012, 9: 772.doi: 10.1038/nmeth.2109.
[24] RONQUIST F, HUELSENBECK J P.MrBayes 3: Bayesian phylogenetic inference under mixed models [J].Bioinformatics, 2003, 19: 1572- 1574.doi: 10.1093/bioinformatics/btg180.
[25] RAMBAUT A.FigTree 1.2.2.[OL].2009.(2018-01-03) http://tree.bio.ed.ac.uk/software/figtree/.
[26] LÜCKING R., CÁCERES M E S, KALB K, et al.Studies in(lichenized Ascomycetes: Lecanorales): II.Six new combi- nations inVězda [J].Lichenologist, 2001, 33(3): 189-194.doi: 10.1006/lich.2000.0318.
[27] SANTESSON R.Foliicolous lichens: I.A revision of the taxonomy of the obligately foliicolous, lichenized fungi [J].Symb Bot Upsal, 1952, 12(1): 1-590.
[28] ANDERSEN H L, EKMMAN S.Disintegration of the Micareaceae (lichenized Ascomycota): A molecular phylogeny based on mitochon- drial rDNA sequences [J].Mycol Res, 2005, 109 (1): 21-30.doi: 10.1017/S0953756204001625.
[29] APRTOOT A, SPARRIUS L B, ALVARADO P., a new genus to accommodate a group of skiophilous temperatespecies that belong in the Pilocarpaceae (lichenized ascomycetes) [J].Gorteria, 2018, 40: 11-14.
[30] SPIRE L, APTROOT A, HERK K V.Asemone, an additional secon- dary substance inin Europe [J].Lichenologist, 2002, 34(5): 447-449.doi: 10.1006/lich.2002.0411.
A Newly Recorded Species offrom China
ZHU Mengli1*, ZHAO Xin1*, WANG Weicheng2,3, JIA Zefeng1**
(1.College of Life Sciences, LiaochengUniversity,Liaocheng 252059, Shandong, China; 2.Institute of Microbiology, Chinese Academy of Sciences,Beijing 100101, China; 3.University of Chinese Academy of Sciences,Beijing 100049, China)
(Vain.) Lücking & Sérus.wasreported as a newly recorded species from China based on the foliicolous lichen specimens collected from Hainan Province.The characteristics of morphology, anatomy and chemistry were provided with microstructure photos.It was characterized by the crustose thallus with light green to orange-red verrucosa, brown apothecia, external exciple prosoplecten- chymatous and narrowly clavate ascospores tapering towards proximal end, (3-)7-septate, 26-36m×2-3m.The phylogenetic tree based on the mtSSU region showed that it had close relationship with the type species(Müll.Arg.) Vězda.A key to known Chinesespecies was also provided.
; Lecanoromycetes; Lichenized fungi; mtSSU; Taxonomy; New record
2021-03-01
2021-04-19
10.11926/jtsb.4398
This work was supported by the National Natural Science Foundation of China (Grant No.31700018; 31800010; 31750001).
ZHU Mengli (Born in 1994), Undergraduate, interesting in taxonomy of lichenous fungi.E-mail: 982340973@qq.com
*Co-first author
Corresponding author.E-mail: zfjia2008@163.com
