马尾松与乡土阔叶树种凋落叶木质素降解的混合效应
2022-02-25李勋张艳覃宇张健
李勋, 张艳, 覃宇, 张健
马尾松与乡土阔叶树种凋落叶木质素降解的混合效应
李勋1, 张艳1, 覃宇2, 张健3*
(1.四川民族学院,横断山区生态修复与特色产业培育研究中心,四川 康定 626001;2.阿坝师范学院,四川 汶川 623002;3.四川农业大学林学院,生态林业工程重点实验室,长江上游生态安全协同创新中心,成都 611130)
为了解森林凋落叶分解过程中木质素的释放规律,对马尾松(, P)、檫木(, S)、香樟(, C)和香椿(, T)凋落叶分解过程中的木质素降解率进行了研究。结果表明,大部分混合凋落叶的木质素在分解过程中出现富集现象,PT和PC组合的木质素含量在第1年较高,之后降低。而PS、PST、PSC、PCT和PSCT组合在0~6、0~9和15~18个月表现出富集现象,其余时期降低。在不同分解时期,部分混合凋落叶组合的木质素降解率表现出非加和效应,呈协同效应,以春季和夏季的协同效应较强,秋冬季较弱。此外,PSCT6121、PSC622、PS64和PC64的木质素降解率在大部分分解时期(≥6/8)表现出协同效应。因此,马尾松与乡土阔叶树种凋落叶混合后促进了木质素的降解,在马尾松人工林改造过程中,与乡土阔叶树种适当混种,可促进凋落叶中木质素的降解。
马尾松;乡土树种;凋落物;木质素
凋落物分解不仅能为植物生长提供不可缺少的养分,同时也是森林生态系统中土壤有机质形成、养分元素矿化和循环的一个重要环节。木质素是凋落物的主要组成部分,其主要由交叉链接的酚聚合物组成,直接关系到森林生态系统的碳循环[1],因此凋落物中木质素的降解在森林生态系统中起到至关重要的作用。作为凋落物中最耐微生物分解的复合物,木质素的分子结构为无定型的三维体形,具有复杂、稳定、多样等特点,因此其降解速率较慢,被认为是凋落物中的难降解物质[2],此外,木质素还能调节凋落叶中其他物质的分解速率。有研究表明,凋落物前期的分解速率受到养分含量、水溶性碳化合物(WSOC)和结构碳化合物(SC)含量的强烈影响,而后期则更多地受到木质素和木质素/N的支配[3]。
马尾松()广泛分布于四川、贵州等17个省,南起雷州半岛(21º41ʹ N),北至秦岭(33º56ʹ N),西至四川盆地中部[4]。马尾松在环境保护、水土保持和治理水土流失等方面发挥着重要的作用。但随着林龄的增长,大面积马尾松人工纯林出现了土壤肥力下降、生物多样性下降等生态安全问题[5]。彭少麟等[6]报道,马尾松人工林的自然更新代严重退化,不同生育期的生长量、平均生长量和总生长量仅为第1代的1/10左右。田大论等[7]也报道,马尾松纯林达到成熟期后,归还给林地的养分速率慢,且需要从土壤中吸收更多的养分以维持自身的生长,进一步加剧林地养分的消耗。檫木()和香樟()不仅凋落量大,叶片养分含量高, 而且对土壤具有较好的改良作用[6-7]。香椿()作为国家II级重点保护野生植物,不仅是珍贵的速生树种,而且对部分重金属元素(如Pb)具有较强的耐受性和吸收能力[8-9],是中国西南地区珍贵的乡土树种。因此,本研究以马尾松与乡土阔叶树种凋落叶为研究对象,通过设置不同的树种组合和混合比例,对马尾松与香椿、檫木和香樟凋落叶混合分解特征进行研究,探讨马尾松与乡土阔叶树种凋落叶混合分解对木质素降解的作用和混合处理促进木质素降解的协同效应,探索森林物质循环的变化规律,充分发挥生物的自肥能力,为马尾松人工林混交树种的选择和科学经营提供理论依据。
1 材料和方法
1.1 研究区概况
本研究在四川省都江堰市四川农业大学试验基地(31°1′~31°2′ N, 103°34′~103°36′ E)进行。该区属于青藏高原向四川盆地的一段过渡地带,典型的亚热带季风湿润气候,年均降水量1 243.8 mm,年均温15.2 ℃。地带性土壤为发育在沙质岩石上的黄壤,质地为重壤土,在中国土壤分类系统中被归为铁铝土[10-11], 海拔约为800 m,主要林地内零散分布马尾松、杉木()、香樟和喜树()。
2016年8月,选择地形地貌、海拔、母岩、土壤类型、坡度、坡位等相近,以及林地情况、林分密度相似的马尾松、喜树等树种混交林为试验样地,设置3个具有代表性的大小约为30 m×30 m的样地(表1),放置凋落物分解袋。
1.2 试验设计
于2016年6月从四川省宜宾市高县(28°11′ N、104°48′ E)分别采集马尾松、檫木、香樟和香椿凋落叶,移除新鲜叶片和已经开始分解的叶片,只保留新鲜的凋落叶,在室温下风干2周, 取凋落物(15.00± 0.05) g放入内部为20 cm×23 cm、上、下层面孔径分别为3.00和0.04 mm的尼龙网分解袋中。根据混交林中主要树种的比例不应低于60%[12-13]设置35个(4个单一处理+31个混合处理)凋落物组合(表2), 共计940袋=35个处理×8个采样时期×3个样地+100 (预防试验过程中凋落袋遗失)。于2016年8月15日将这些凋落袋小心转移到3个样地,将凋落叶分解袋随机水平放置于样地表面,样品袋间距为2~5 cm, 从而避免样品之间相互干扰。采样时随机收集每种凋落物组合3份凋落物带回实验室,测定样品的损失量。
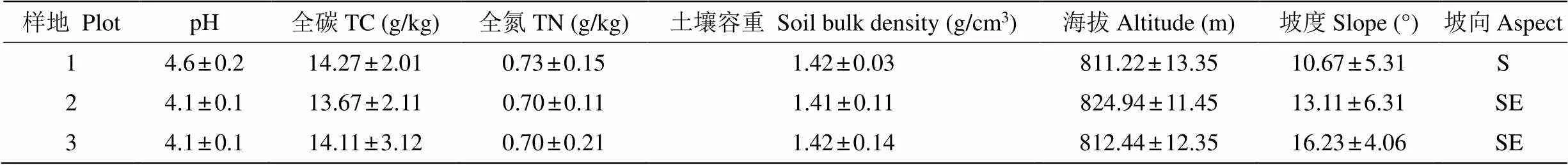
表1 样地基本信息
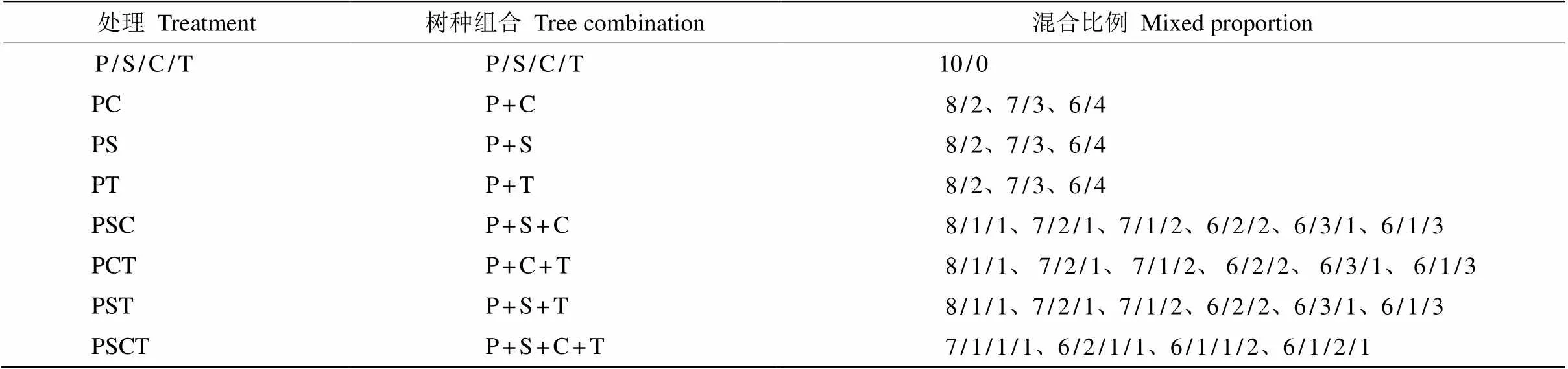
表2 试验设计
P: 马尾松; S: 檫木; C: 香樟; T: 香椿。
P:; S:; C:; T:.
1.3 样品采集
2016年8月开始试验,分别于2016年11月、2017年2、5、8和11月和2018年2、5和8月每隔3个月收集1次凋落物分解袋,相对应的分解时间分别为3、6、9、12、15、18、21和24个月。每次从3个样地随机收集每个凋落物组合分解袋3袋,用镊子将凋落叶分解后的大片碎片取出,尽可能仔细地去除土壤颗粒物、节肢动物和外来植物根系,测量凋落叶的含水量和干质量,剩余的凋落叶清理干净后备用。
1.4 样品分析
将凋落叶置于65 ℃烘箱干燥48 h后称量干质量,以评估分解后残留的凋落叶质量。取干燥样品在磨粉机中研磨,过0.25 mm的筛子以备用。木质素和纤维素含量测定采用改进的酸性洗涤法[14],全碳含量采用重铬酸钾加热法(GB 7657-8)测定;全氮含量采用凯氏定氮法(LY/T 1269-1999)测定;磷含量采用钼锑抗比色法(LY/T 1270-1999);总酚含量采用福林酚比色法[15]测定;缩合单宁含量采用香草醛盐酸法[16]。
1.5 数据的统计和分析
采用修正的Olson指数衰减模型模拟凋落叶分解过程,=-kt, 其中,为凋落叶的残留率(%),k为凋落叶分解常数,为分解时间。
凋落叶木质素降解率=(1-MC/00)×100%, 其中,为当次木质素降解率(%),M为当次凋落叶残留量(g),0为凋落叶初始质量(g),C为当次木质素含量(g/kg),0为木质素初始含量(g/kg)。
将不同物种凋落叶按照一定比例组合起来,得出混合凋落叶的预期木质素降解率,混合凋落叶木质素降解的非加性效应[17]T(%)=(R+R+…+R)/, 其中为物种数量,R为包含物种木质素降解率的测量值。
采用单因素方差分析(One-way ANOVA)和Turkey法比较不同组合间的差异显著性。用Levene’s法检验方差同质性,不满足该假设的数据先进行Log转化。根据=0.05的独立检验确定木质素降解率的观测值和预期值间的差异。根据统计结果,将混合效应分为加和效应(观测值和预期值之间没有显著差异)和非加和效应(观测值和预期值之间有显著差异)。其中,非加和效应又分为协同效应(观测值-预期值>0,且<0.05)和拮抗效应(观测值-预期值<0,且<0.05)。
采用偏最小二乘法(PLS)回归分析凋落叶初始质量对凋落叶木质素降解率的混合效应(观测值-预期值)的相对重要性。模型中单个预测因子的相对重要性由重要性变量(VIP)估计,VIP>1表示预测因子对因变量变化的显著贡献。所有统计分析均采用SPSS 25.0进行,图表用Excel和Origin软件制作。
2 结果和分析
2.1 凋落叶初始物质含量
由表3可知,马尾松凋落叶的初始C、木质素、纤维素、总酚和缩合单宁含量、木质素/N及木质素/P均显著高于另3种阔叶乡土树种凋落叶,C/N和C/P则显著高于香樟和香椿。香椿凋落叶的N和P含量最高,檫木和香樟凋落叶次之,马尾松凋落叶最低,但4种树种凋落叶的N/P无显著差异。
31个混合处理凋落叶的初始物质含量见表4。马尾松所占比例越大,混合凋落叶的C、木质素、纤维素、总酚、缩合单宁含量及木质素/N、木质素/P越高;香椿和香樟凋落叶所占比例越大,混合凋落叶的N和P含量越高。
同行数据后不同字母表示差异显著(<0.05)。
Data followed different letters in the same line indicate significant differences at 0.05 level.
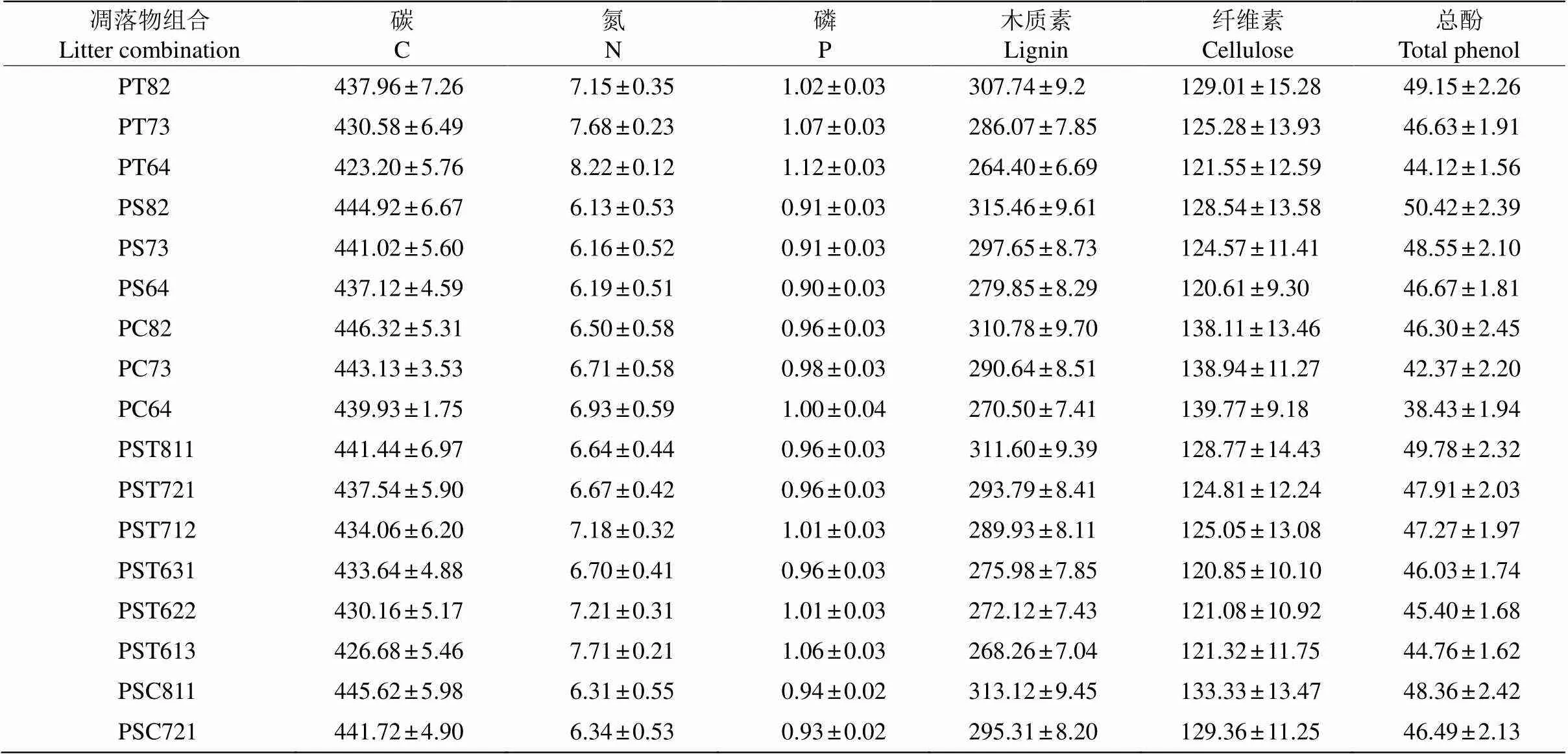
表4 混合树种凋落叶初始质量特征
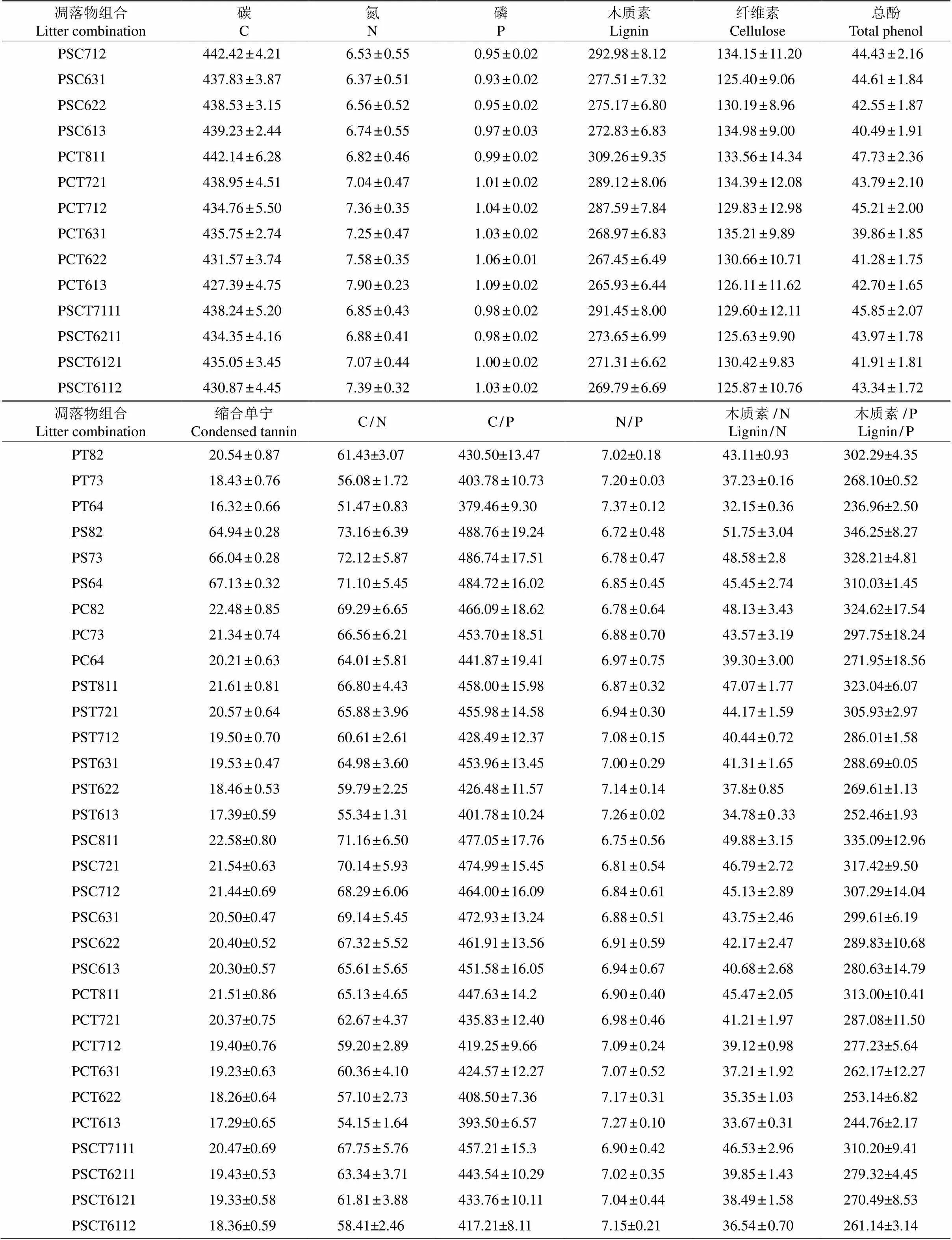
续表(Continued)
P: 马尾松; S: 檫木; C: 香樟; T: 香椿。
P:; S:; C:; T:.
2.2 凋落叶质量损失的动态变化
从表5可见,4个单树种凋落叶间的值存在显著差异(=60.54,<0.001),以香椿凋落叶的最高(1.27)、香樟(1.01)和檫木(0.88)次之,马尾松的最低(0.49)。此外,4个单树种凋落叶与部分混合凋落叶间的值差异显著(=17.00,<0.001)。
所有混合凋落叶的值均低于香椿和香樟凋落叶,而高于马尾松凋落叶。PSCT6112的值(0.89)略高于檫木(0.88),显著高于64.53% (20/31)的混合处理。此外,PT64 (0.87)、PST613 (0.86)、PST622 (0.85)、PC64 (0.83)、PT73 (0.82)、PCT613 (0.81)和PSCT6121 (0.81)的值略低于檫木凋落叶(= 1.79,=0.54),且显著高于29.03% (9/31)的混合处理。在所有混合凋落叶中,马尾松占总质量60%的组合(马尾松∶阔叶=6∶X,下同)的值均高于马尾松占总质量70% (7∶X)和80% (8∶X)的组合。
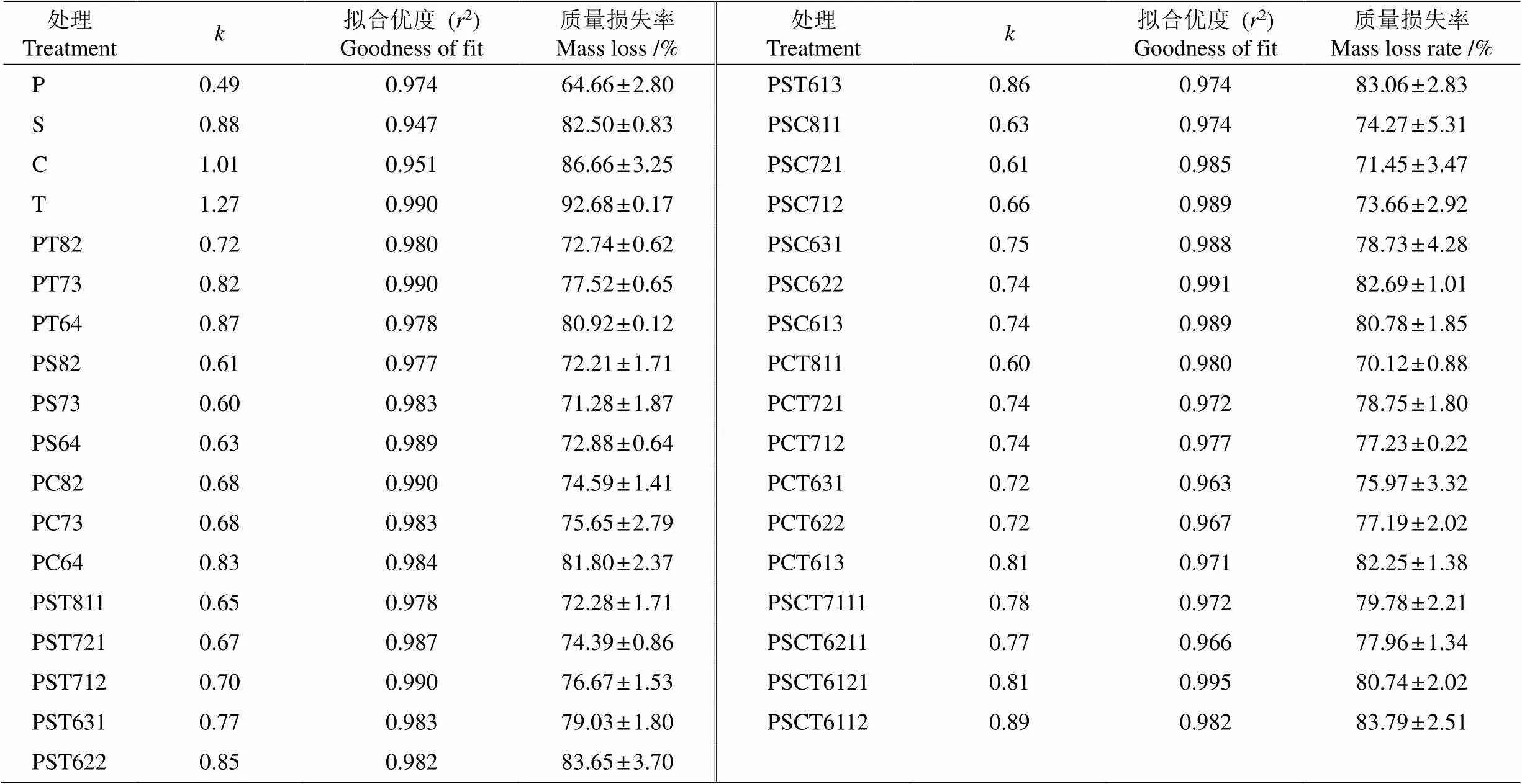
表5 凋落叶的年分解系数(k)和分解末期(24个月)凋落叶的质量损失率
P: 马尾松; S: 檫木; C: 香樟; T: 香椿。
P: Pinus massoniana; S: Sassafras tzumu; C: Cinnamomum camphora; T: Toona sinensis.
2.3 木质素含量的动态变化
由图1: H可知,分解3~9个月后,4个单树种凋落叶的木质素含量均有所升高,表现出富集现象,而除香椿和马尾松凋落叶在分解15~18个月升高外,其他时期不断降低。檫木和香樟凋落叶分解3个月后的木质素含量分别升至311.95和333.54 g/kg。
31个混合凋落叶木质素含量在分解过程中均表现出富集现象,但发生的时间有所差异,PT组合经0~6个月分解木质素含量不断升高,分解6~9个月时降低;PT82和PT73经9~12个月分解木质素含量升高;分解9~24个月,3个混合比例的PT组合木质素含量均不断降低(图1: A)。PC组合在0~9个月表现出富集现象,9~15个月出现迅速降低, 之后缓慢降低(图1: C)。而PS、PST、PSC、PCT和PSCT组合的木质素含量动态变化与4个单树种的基本一致,即在0~6或0~9或15~18个月表现出富集现象,其余时期木质素含量不断降低。
2.4 木质素降解的混合效应
混合凋落叶在分解3个月后就有41.94%的组合表现为非加和效应中的协同效应(观测值-预期值>0,<0.05),且随分解时间呈先降低后升高再降低的趋势(图2)。分解9个月(图2: C)和12个月(图2: D)后,分别有58.06%和64.52%的混合处理表现出协同效应,分解15个月后(图2: E),协同效应最弱,仅有16.13% (5/31)。而其他分解时期,分别有41.94% (图2: A,13/31)、35.48% (图2: B,11/31)、38.71% (图2: F,12/31)、48.39% (图2: G,15/31)和38.71% (图2: H,12/31)的混合凋落叶表现出协同效应。此外,除分解9个月外,其他7个分解时期均有个别混合处理(3.23%~6.45%,1/31~2/31)表现出拮抗效应(观测值-预期值<0,<0.05)。总体上看,不同处理凋落叶的混合效应在第1年分解期的春季(图2: C)和夏季(图2: D)较强,分解15个月后(图2: E)协同效应减弱,之后增强。
8次采样中,不同树种间木质素降解的混合效应表现为:PSCT、PC、PS的协同效应较强,分别为68.75% (22/32)、66.67% (16/24)和50.00% (12/24), 其次为PSC (43.75%, 21/48),而PT、PCT、PST的协同效应则相对较弱,分别为37.50% (9/24)、27.08% (13/48)和25.00% (12/48)。其中PS、PC和PSCT组合在分解9和12个月后,PT和PC组合在分解18个月后,PC组合在分解21个月后以及PT组合在分解24个月后均表现出协同效应。在31个混合处理中, PSCT6121 (8/8)、PSC622 (7/8)、PS64 (6/8)、PC64 (6/8)在≥75% (6/8)的分解时期表现出协同效应。
变量重要性指标(VIP值)用来测度每个自变量在解释因变量(木质素混合效应)的作用(图3)。凋落叶的初始物质含量和化学计量比对解释木质素混合效应的作用依次为木质素/N>木质素>N>总酚>C> C/N>单宁>木质素/P>C/P>N/P>P>纤维素。其中,凋落叶中木质素的混合效应与N含量呈显著正相关,与木质素/N和木质素含量呈显著负相关。
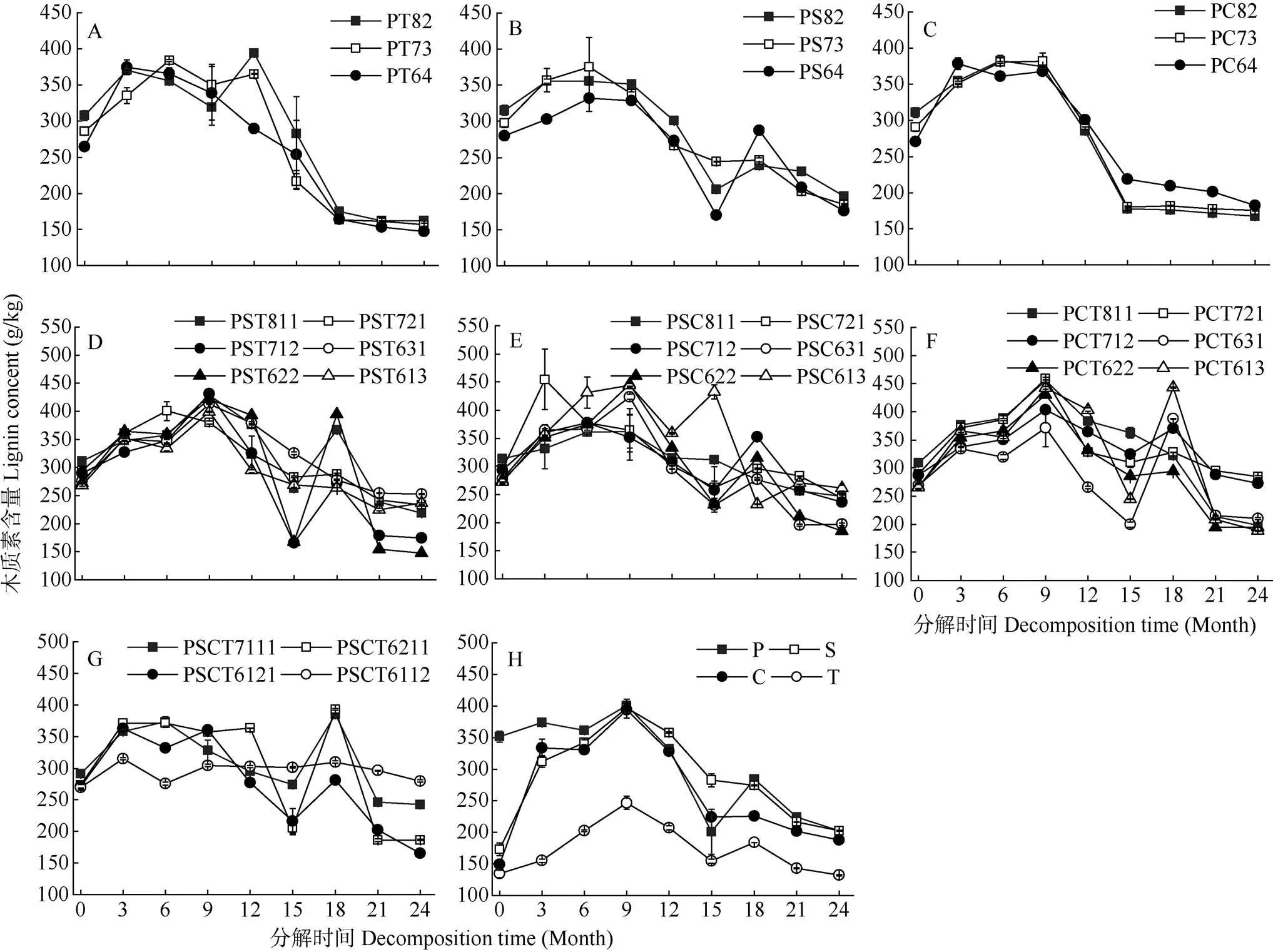
图1 凋落叶中木质素含量的动态变化。P: 马尾松; S: 檫木; C: 香樟; T: 香椿。
3 结论和讨论
凋落物质量通常用凋落物中养分含量的多少衡量,并以含碳化合物量(如木质素、纤维素等)与养分元素含量(N、P、K等)的比来表示,其中木质素/N和木质素含量是决定凋落物质量和分解快慢的关键指标[18-19]。总体上看,在前6或9个月的分解中,凋落叶的木质素含量随分解时间延长而升
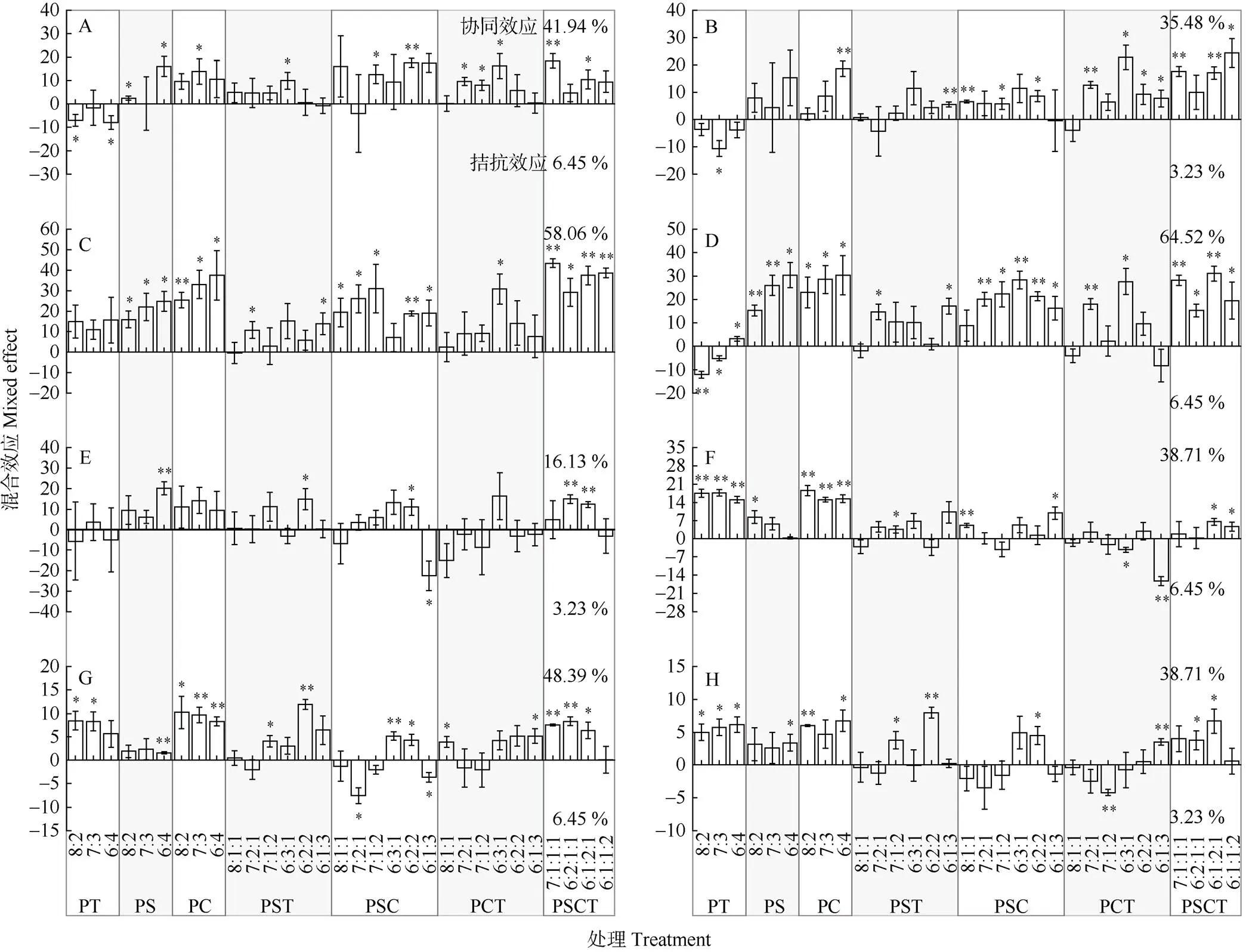
图2 混合凋落叶的木质素降解混合效应。P: 马尾松; S: 檫木; C: 香樟; T: 香椿。A~H分别表示分解3、6、9、12、15、18、21和24个月; *: P<0.05; **: P<0.01。
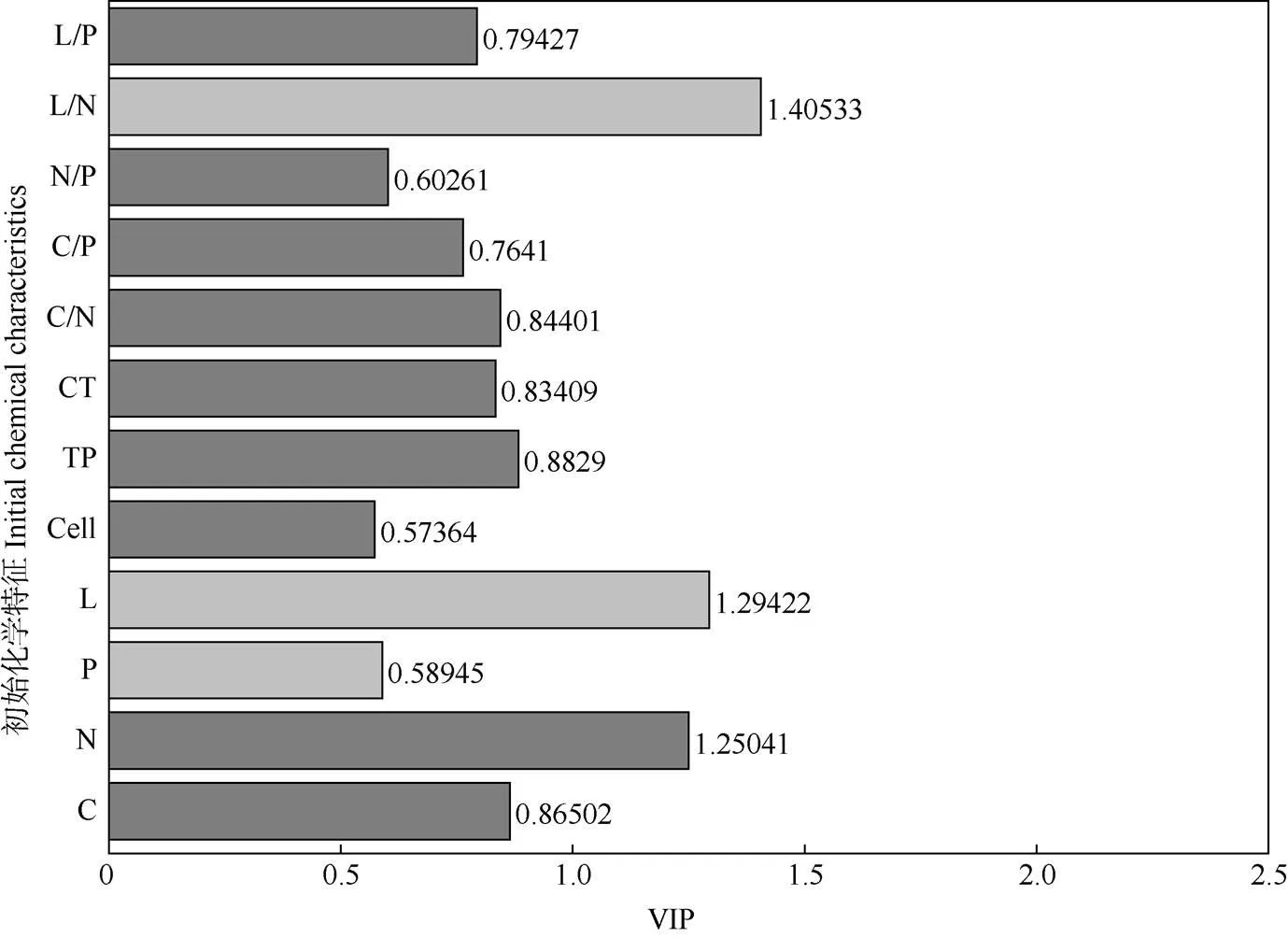
图3 凋落叶初始化学特征与木质素混合效应的偏最小二乘回归分析。L: 木质素; Cell: 纤维素; TP: 总酚; CT: 缩合单宁;L/N: 木质素/N; L/P: 木质素/P。
高,表现出富集现象,之后缓慢降低。这可能是因为木质素在凋落物分解中被认为是一种“难降解物质”,其与大分子物质(纤维素、蛋白质等)结合后, 形成一种类似于“屏障”的结构,限制土壤动物和微生物进入并抑制了分解者的生理代谢活动[20]。同时, 凋落物中易分解组分(可溶性碳、氮、磷等)在分解初期快速损失,导致易分解组分在凋落物中所占比例迅速降低,使得木质素等难分解组分所占比例有所提高。木质素的分解主要是由白腐菌、软腐菌和褐腐菌等3种分解者来实现的[21]。凋落物放置地面后,要经过一段时间的菌群转换才能形成适合分解木质素的主要真菌群落并产生对应的酶[22],这也是导致木质素分解前期富集的原因。在之后的分解过程,微生物将木质素分子结构破坏,凋落物失去外围“屏障”的保护,凋落叶分解也处于水分、温度等条件适宜的环境中,开始进入快速分解的阶段,木质素浓度也开始降低。因此混合凋落叶中木质素含量表现出前期富集,后期净释放的变化规律。此外,2 a分解期内,凋落叶中木质素的协同效应在春季和夏季较强,这可能是因为随着分解时间的进行, 凋落物基质质量和分解环境不断变化,土壤生物群落结构也在不断改变:经过几个月的旱季后,凋落叶迎来水热条件较好的雨季(春季和夏季),土壤生物迅速生长繁殖,能够降解木质素的胞外酶(如锰氧化物酶、漆酶等)被大量分泌[23],木质素经过酶解后被降解为小分子的芳香族化合物, 从而有利于进一步的分解。
木质素是地球上含量最丰富的芳香族化合物, 其降解是维持森林生态系统中碳素平衡不可缺少的物质循环过程。木质素是由结构复杂、稳定、多样的无定型三维体形大分子构成,且能与纤维素、蛋白质等大分子物质结合形成类似“屏障”的结,因此其降解相对较慢,被认为是凋落物中的难降解物质[24]。本研究结果表明,马尾松和3种乡土树种凋落叶按不同比例混合分解2 a后,不同树种组合PSCT、PC、PS和PSC的协同效应较强,而PT、PCT、PST组合的相对较弱。其中,PSCT6121的木质素降解率始终表现出协同效应,PSC622、PS64、PC64在≥75% (6/8)的时期表现出协同效应,可见, 凋落物的树种组成和混合比例对木质素降解均有显著影响。相关分析表明,凋落叶的初始物质含量和化学计量比对解释木质素混合效应的作用依次为: 木质素/N>木质素>N>总酚>C>C/N>单宁>木质素/P>C/P> N/P>P>纤维素含量。其中,木质素混合效应与凋落叶初始N含量呈显著正相关,与初始木质素含量和木质素/N呈显著负相关,这与前人[25-28]的研究结果一致,即N的含量与凋落叶的分解速率呈正相关,N在一定程度上越高越有利于凋落物分解。这是因为生物降解是亚热带林区凋落物木质素降解的主要途径[29],而N是土壤生物生存不可缺少的营养元素,也是生物细胞形成所必需的元素,较高的N含量能增强土壤生物的生理代谢活动,有利于土壤生物的生长和繁殖,改善凋落叶分解的微环境,使得凋落物中木质素的降解也随之加快。近期有研究表明, 单一马尾松凋落叶会造成森林土壤酸化,恶化凋落物分解微环境,从而难以形成群落结构稳定、功能丰富的微生物群落[30]。同时马尾松纯凋落叶分解过程中会释放化感物质,进而让凋落叶附近的土壤动物群落单一[31-32],不利于木质素降解。本研究将马尾松与阔叶树种凋落物混合后,增加了凋落物多样性,丰富的凋落物种类为土壤生物提供了充足的养分来源,部分土壤生物(如真菌)可以使木质素从坚固的结构转化为可溶性的聚合物状态[33-34],进而促进了凋落物中木质素的降解。而单宁与蛋白质可以形成不易消化复合物,使蛋白质不能发生正常的新陈代谢,从而抑制微生物活性[35]或降低微生物群落组成的多样性[36]。而本研究中阔叶占比较高的混合凋落物, 其缩合单宁含量相对较低, 因此凋落物中微生物的活性则相对较强,更能促进凋落物中木质素的降解。此外,线虫是土壤动物中数量最多的一种[37],凋落叶中酚类物质可以影响土壤线虫的活性和数量[38], 而食细菌线虫能通过选择性取食而对微生物数量、活性和组成产生影响[39],土壤线虫总数和食细菌线虫总数整体随酚类物质含量的下降呈上升的趋势[40]。本研究中阔叶占比较高的混合凋落物的酚类物质相对较低,导致土壤线虫和食细菌线虫总数相对更高,从而促进了凋落叶中木质素的降解。因此,这些马尾松占比较低的混合处理(PSCT6121、PSC622、PS64和PC64)的协同效应更强。
作为凋落物的主要组成部分之一,木质素在凋落物分解的过程中起着重要作用。马尾松与乡土阔叶树种混合分解后,不同分解时期的木质素降解率均表现出非加和效应,主要表现为协同效应,其中,木质素在夏季和冬季表现出较强的协同效应。在PSCT6121、PSC622、PS64和PC64混合处理中, 木质素降解的协同效应较强,表明凋落物的物种多样性和混合比例对木质素的降解均具有显著影响。在马尾松人工林改造过程中,可将檫木、香樟和香椿作为伴生树种,设置适当的混合比例,从而促进凋落叶中木质素的降解。由于本研究筛选出的混合效应较强的处理是凋落叶之间的比例,而不是树种之间的株数比,需要结合造林密度以及不同树种的年凋落量进行换算。
[1] DENG R J, YANG W Q, FENG R F, et al.Mass loss and element release of litter in the subalpine forest over one freeze-thaw season [J].Acta Ecol Sin, 2009, 29(10): 5730-5735.doi: 10.3321/j.issn:1000-0933.2009.10.065.
邓仁菊, 杨万勤, 冯瑞芳, 等.季节性冻融期间亚高山森林凋落物的质量损失及元素释放 [J].生态学报, 2009, 29(10): 5730-5735.doi: 10.3321/j.issn:1000-0933.2009.10.065.
[2] AUSTIN A T, BALLARÉ C L.Dual role of lignin in plant litter decomposition in terrestrial ecosystems [J].Proc Natl Acad Sci USA, 2010, 107(10): 4618-4622.doi: 10.1073/pnas.0909396107.
[3] ZENG F, QIU Z J, XU X Y.Review on forest litter decomposition [J].Ecol Environ Sci, 2010, 19(1): 239-243.doi: 10.3969/j.issn.1674-5906.2010.01.044.
曾锋, 邱治军, 许秀玉.森林凋落物分解研究进展 [J].生态环境学报, 2010, 19(1): 239-243.doi: 10.3969/j.issn.1674-5906.2010.01.044.
[4] LI X, ZHANG D J, ZHANG Y, et al.Forest gap size can efficiently promote litter decomposition and nutrient release in south-western China [J].S For J For Sci, 2019, 81(2): 185-194.doi: 10.2989/2070 2620.2018.1542564.
[5] LI M J, DU M F, NIE C J.Research advances in soil improvement ofplantation [J].World For Res, 2014, 27(5): 31-36.doi: 10.13348/j.cnki.sjlyyj.2014.05.006.李明军, 杜明凤, 聂朝俊.马尾松人工林地力维护研究进展 [J].世界林业研究, 2014, 27(5): 31-36.doi: 10.13348/j.cnki.sjlyyj.2014.05.006.
[6] LI X, ZHANG D J, ZHANG Y, et al.The edge effect of a forest gap on decomposition ofandleaf litter [J].Chin J Appl Environ Biol, 2017, 23(3): 570-578.doi: 10.3724/SP.J.1145.2017.02026.李勋, 张丹桔, 张艳, 等.林窗边缘效应对马尾松和香樟凋落叶分解的影响 [J].应用与环境生物学报, 2017, 23(3): 570-578.doi: 10.3724/SP.J.1145.2017.02026.
[7] LIU J, HONG W, WU C Z, et al.Biomass production and productivity of natural regenerated sassafras tzumu trees [J].J Trop Subtrop Bot, 2002, 10(2): 105-110.doi: 10.3969/j.issn.1005-3395.2002.02.002.
柳江, 洪伟, 吴承祯, 等.天然更新的檫木林的生物量和生产力 [J].热带亚热带植物学报, 2002, 10(2): 105-110.doi: 10.3969/j.issn.1005-3395.2002.02.002.
[8] QIN Y, ZHANG D J, LI X, et al.Changes of total phenols and condensed tannins during the decomposition of mixed leaf litter ofand broad-leaved trees [J].Chin J Appl Ecol, 2018, 29(7): 2224-2232.doi: 10.13287/j.1001-9332.201807.038.
覃宇, 张丹桔, 李勋, 等.马尾松与阔叶树种混合凋落叶分解过程中总酚和缩合单宁的变化 [J].应用生态学报, 2018, 29(7): 2224- 2232.doi: 10.13287/j.1001-9332.201807.038.
[9] LI X J.Studies on resistance ofto heavy metal [D].Tianjin: Tianjin University Technology, 2012: 1-81.李晓晶.香椿对重金属的抗性研究 [D].天津: 天津理工大学, 2012: 1-81.
[10] XIA J G, DENG L J, ZHANG L P, et al.Study on soil taxonomy in Sichuan [J].J Sichuan Agric Univ, 2002, 20(2): 117-122.doi: 10.3969/j.issn.1000-2650.2002.02.013.
夏建国, 邓良基, 张丽萍, 等.四川土壤系统分类初步研究 [J].四川农业大学学报, 2002, 20(2): 117-122.doi: 10.3969/j.issn.1000-2650.2002.02.013.
[11] ZHOU Y, CAI C F, WANG Q Y.Taxonomic nomenclature and analogy of soil in Hubei Province [J].Soil, 1997, 3(3): 130-136.
周勇, 蔡崇法, 王庆云.湖北省土壤系统分类命名及其类比 [J].土壤, 1997, 3(3): 130-136.
[12] CHEN D X, CHEN Q, MENG Y H, et al.High-efficient management technology of big-diameter timber of[J].J S Agric, 2015, 46(2): 293-298.doi: 10.3969/jissn.2095-1191.2015.2.293.
陈代喜, 陈琴, 蒙跃环, 等.杉木大径材高效培育技术探讨 [J].南方农业学报, 2015, 46(2): 293-298.doi: 10.3969/jissn.2095-1191.2015.2.293.
[13] LÜ S Y.Several basic viewpoints on construction of mixed forests [J].Yunnan For Sci Technol, 2001, 1(1): 26-28.doi: 10.3969/j.issn.1672- 8246.2001.01.005.
吕树英.关于营造混交林的几个基本观点 [J].云南林业科技, 2001, 1(1): 26-28.doi: 10.3969/j.issn.1672-8246.2001.01.005.
[14] VAN DE RBILT K L, WHITE C S, HOPKINS O, et al.Aboveground decomposition in arid environments: Results of a long-term study in central New Mexico [J].J Arid Environ, 2008, 72(5): 696-709.doi: 10.1016/j.jaridenv.2007.10.010.
[15] SCHOFIELD J, HAGERMAN A E, HAROLD A.Loss of tannins and other phenolics from willow leaf litter [J].J Chem Ecol, 1998, 24(8): 1409-1421.doi: 10.1023/A:1021287018787.
[16] COQ S, SOUQERT J M, MEUDEC E, et al.Interspecific variation in leaf litter tannins drives decomposition in a tropical rain forest of French Guiana [J].Ecology, 2010, 91(7): 2080-2091.doi: 10.1890/ 09-1076.1.
[17] TARDIF A, SHIPLEY B.The relationship between functional dis- persion of mixed-species leaf litter mixtures and species’ interactions during decomposition [J].Oikos, 2015, 124(8): 1050-1057.doi: 10.1111/oik.01686.
[18] SCHLESINGER W H, HASEY M M.Decomposition of chaparral shrub foliage: Losses of organic and inorganic constituents from deciduous and evergreen leaves [J].Ecology, 1981, 62(3): 762-774.doi:10.2307/1937744.
[19] MELILLO J M, ABER J D, MURATORE J F.Nitrogen and lignin control of hardwood leaf litter decomposition dynamics [J].Ecology, 1982, 63(3): 621-626.doi: 10.2307/1936780.
[20] FENG L, SONG S H, ZHAO L, et al.Progress in plant polyphenols and their physiological functions [J].Acta Agric Jiangxi, 2007, 19(10): 105-107.doi: 10.3969/j.issn.1001-8581.2007.10.035.
冯丽, 宋曙辉, 赵霖, 等.植物多酚种类及其生理功能的研究进展 [J].江西农业学报, 2007, 19(10): 105-107.doi: 10.3969/j.issn.1001- 8581.2007.10.035.
[21] FENG L, HU B, ZHAO Y L, et al.Study of disposing city defoliation by cellulose degradation bacteria system [J].J Anhui Agric Sci, 2009, 37(6): 2399-2401.doi: 10.3969/j.issn.0517-6611.2009.06.028.
冯蕾, 胡滨, 赵艳丽, 等.复合纤维素降解菌对城市落叶的降解研究 [J].安徽农业科学, 2009, 37(6): 2399-2401.doi: 10.3969/j.issn.0517-6611.2009.06.028.
[22] HUANG X P, MONK C.Purification and characterization of a cellulase (CMCase) from a newly isolated thermophilic aerobic bacteriumgen.nov.sp.Nov [J].World J Microbiol Biol, 2004, 20(1): 85-92.doi: 10.1023/B:WIBI.000001 3316.12730.e7.
[23] PUCHETA E, LLANOS M, MEGLIOLI C, et al.Litter decomposition in a sandy Monte desert of western Argentina: Influences of vegetation patches and summer rainfall [J].Aust Ecol, 2006, 31(7): 808-816.doi:10.1111/j.1442-9993.2006.01635.x.
[24] OSONO T, HIRADATE S, HOBARA S.Bleaching of leaf litter accelerates the decomposition of recalcitrant components and mobi- lization of nitrogen in a subtropical forest [J].Sci Rep, 2021, 11(1): 1787.doi: 10.1038/s41598-021-81206-7.
[25] GUO Z L, ZHENG J P, MA Y D, et al.Researches on litterfall decomposition rates and model simulating of main species in various forest vegetations of Changbai Mountains, China [J].Acta Ecol Sin, 2006, 26(4): 1037-1046.doi: 10.3321/j.issn:1000-0933.2006.04.009.
郭忠玲, 郑金萍, 马元丹, 等.长白山各植被带主要树种凋落物分解速率及模型模拟的试验研究 [J].生态学报, 2006, 26(4): 1037- 1046.doi: 10.3321/j.issn:1000-0933.2006.04.009.
[26] LI Y N, ZHOU X M, ZHANG N L, et al.The research of mixed litter effects on litter decomposition in terrestrial ecosystems [J].Acta Ecol Sin, 2016, 36(16): 4977-4987.doi: 10.5846/stxb201501200165.
李宜浓, 周晓梅, 张乃莉, 等.陆地生态系统混合凋落物分解研究进展 [J].生态学报, 2016, 36(16): 4977-4987.doi: 10.5846/stxb2015 01200165.
[27] LI Z A, ZOU B, DING Y Z, et al.Key factors of forest litter decom- position and research progress [J].Chin J Ecol, 2004, 23(6): 77-83.
李志安, 邹碧, 丁永祯, 等.森林凋落物分解重要影响因子及其研究进展 [J].生态学杂志, 2004, 23(6): 77-83.
[28] LIN K M, ZHANG Z Q, CAO G Q, et al.Decomposition character- ristics and its nutrient dynamics of leaf litter mixtures of both Chinese fir and[J].Acta Ecol Sin, 2006, 26(8): 2732-2738.doi: 10.3321/j.issn:1000-0933.2006.08.041.
林开敏, 章志琴, 曹光球, 等.杉木与楠木叶凋落物混合分解及其养分动态 [J].生态学报, 2006, 26(8): 2732-2738.doi: 10.3321/j.issn: 1000-0933.2006.08.041.
[29] MARTÍNEZ Á T, SPERANZA M, RUIZ-DUEÑAS F J, et al.Biode- gradation of lignocellulosics: Microbial, chemical, and enzymatic aspects of the fungal attack of lignin [J].Int Microbiol, 2005, 8(3): 195-204.doi: 10.1016/j.ibiod.2005.05.002.
[30] MO J M, PENG S L, FANG Y T, et al.A preliminary study on the dynamics of bio-available nitrogen in soils of pine-broadleaf mixed forest in Dinghuashan Biosphere Reserve [J].Acta Ecol Sin, 2001, 21(3): 492-497.doi: 10.3321/j.issn:1000-0933.2001.03.024.
莫江明, 彭少麟, 方运霆, 等.鼎湖山马尾松针阔叶混交林土壤有效氮动态的初步研究 [J].生态学报, 2001, 21(3): 492-497.doi: 10.3321/j.issn:1000-0933.2001.03.024.
[31] WANG W J, YANG W Q, TAN B, et al.Contributions of soil fauna to litter decomposition in subtropical evergreen broad-leaved forests in Sichuan basin [J].Ecol Environ Sci, 2013, 22(9): 1488-1495.doi: 10.3969/j.issn.1674-5906.2013.09.006.
王文君, 杨万勤, 谭波, 等.四川盆地亚热带常绿阔叶林土壤动物对几种典型凋落物分解的影响 [J].生态环境学报, 2013, 22(9): 1488-1495.doi: 10.3969/j.issn.1674-5906.2013.09.006.
[32] YANG D X, YANG M F, LÜ Z Y.Study on soil fauna in the burned area of natural pine forests in Karst Region [J].Guangdong Agric Sci, 2012, 39(22): 187-189.doi: 10.3969/j.issn.1004-874X.2012.22.061.
杨大星, 杨茂发, 吕召云.喀斯特马尾松自然林火烧迹地土壤动物调查 [J].广东农业科学, 2012, 39(22): 187-189.doi: 10.3969/j.issn.1004-874X.2012.22.061.
[33] HIGUCHI T.Microbial degradation of lignin: Role of lignin pero- xidase, manganese peroxidase, and laccase [J].Proc Jap Acad, 2004, 80(5): 204-214.doi: 10.2183/pjab.80.204.
[34] LI H T, YAO K, HE Q, et al.Biodegradation and applications of lignin [J].Leather Sci Eng, 2010, 20(6): 27-31.doi: 10.3969/j.issn.1004-7964.2010.06.007.
李海涛, 姚开, 何强, 等.木质素的生物降解及其应用 [J].皮革科学与工程, 2010, 20(6): 27-31.doi: 10.3969/j.issn.1004-7964.2010.06.007.
[35] SCHIMEL J P, CATES R G, RUESS R.The role of balsam poplar secondary chemicals in controlling soil nutrient dynamics through succession in the Alaskan Taiga [J].Biogeochemistry, 1998, 42(1/2): 221-234.doi: 10.1023/A:1005911118982.
[36] BAPTIST F, ZINGER L, CLEMENT J C, et al.Tannin impacts on microbial diversity and the functioning of alpine soils: A multi- disciplinary approach [J].Environ Microbiol, 2008, 10(3): 799-809.doi: 10.1111/j.1462-2920.2007.01504.x.
[37] VAN DE HOOGEN J, GEISEN S, ROUTH D, et al.Soil nematode abundance and functional group composition at a global scale [J].Nature, 2019, 572(7768): 194-198.doi: 10.1038/s41586-019-1418-6.
[38] HÄTTENSCHWILER S, JØRGENSEN H B.Carbon quality rather than stoichiometry controls litter decomposition in a tropical rain forest [J].J Ecol, 2010, 98(4): 754-763.doi: 10.1111/j.1365-2745.2010.01671.x.
[39] ZHOU J H.Impact on prometryne degradation in soil through the interactions between nematodes and microbes [D].Nanjing: Nanjing Agricultural University, 2011: 119.doi: 10.7666/d.Y2038607.
周际海.线虫与微生物相互作用及其对污染土壤扑草净解影响的研究 [D].南京: 南京农业大学, 2011: 119.doi: 10.7666/d.Y2038 607.
[40] LI H Q, LIU Q Z, ZHANG L L, et al.Accumulation of phenolic acids in the monocultured strawberry soils and their effect on soil nematodes [J].Chin J Ecol, 2014, 33(1): 169-175.doi: 10.13292/j.1000-4890.20131220.0023.
李贺勤, 刘奇志, 张林林, 等.草莓连作土壤酚酸类物质积累对土壤线虫的影响 [J].生态学杂志, 2014, 33(1): 169-175.doi: 10.13292/j.1000-4890.20131220.0023.
Mixed Effects on Lignin Degradation in the Litter Leaves ofand Native Broad-leaved Tree Species
LI Xun1, ZHANG Yan1, QIN Yu2, ZHANG Jian3*
(1.Research Center for Ecological Restoration and Characteristic Industry Cultivation in Hengduan Mountains Region, Sichuan Minzu College, Kangding 626000, Sichuan, China; 2.Aba Teachers University, Wenchuan 623002, Sichuan, China; 3.Key Laboratory of Forestry Ecological Engineering in Sichuan, Collaborative Innovation Center of Ecological Security in the Upper Reaches of Yangtze River, Sichuan Agricultural University,Chengdu 611130, China)
To understand the release rule of lignin in the decomposition process of forest leaf litter, the lignin degradation rate of leaf litter of(P),(S),(C) and(T) was studied.The results showed that lignin in most of mixed litter was enriched during decomposition.The lignin content in combination of PT and PC was high in the first year, and then decreased.However, the lignin content in combination of PS, PST, PSC, PCT and PSCT were enriched at 0-6, 0-9 and 15- 18 months, and decreased at other periods.At all decomposition stages, the degradation rate of lignin in some mixed litters showed synergistic effect rather than additive effect, the synergistic effect was stronger in summer and winter than in other seasons.Besides, the lignin degradation rate of PSCT6121, PSC622, PS64 and PC64 showed a synergistic effect in most decomposition stages (≥6/8).Therefore, the mixed leaves ofand native broad-leaved trees promoted the degradation of lignin, the degradation of lignin in the leaf litter ofplantation could be promoted by proper mixing with native broad-leaved species.
; Native tree species; Litter; Lignin
10.11926/jtsb.4408
2021-03-09
2021-05-19
国家自然科学基金项目(31370628); 四川省科技支撑计划项目(12ZC0017); 四川民族学院自办科研项目(XYZB2003ZA, XYZB2016ZB); 四川民族学院特色科研孵化项目(KBFH2103); 四川省大学生创新创业训练计划项目(S202011661092, S202011661106, S202011661090)资助
This work was supported by the National Natural Science Foundation of China (Grant No.31370628), the Project for Science and Technology Support of Sichuan (Grant No.12ZC0017), the Project for Scientific Research of Sichuan Minzu College (Grant No.ZYZB2003ZA, XYZB2016ZB), the Project for Characteristic Research Incubation of Sichuan Minzu College (Grant No.KBFH2103), and the Project for Innovation and Entrepreneurship Training for College Students in Sichuan (Grant No.S202011661092, S202011661106, S202011661090).
李勋,男,博士,主要从事长江中上游马尾松低效人工林改造。E-mail:13980765825@qq.com
通信作者Corresponding author.E-mail:sicauzhangjian@163.com
