热带桉树树干液流的时滞效应分析
2022-02-25王城城叶文伟赵从举陈丽艳
王城城,叶文伟,赵从举,陈丽艳
(海南师范大学 地理与环境科学学院,海口 570100)
0 引 言
【研究意义】蒸腾是植物将体内的水分以水蒸气的形式蒸发到大气中的过程,是植物耗水的主要方式,不仅反映了植物的水分状况与环境间的关系,还影响生态系统的水量平衡[1-3]。研究液流的变化规律及其与环境因子间的关系是揭示植物蒸腾作用对区域生态环境水量平衡的重要手段。植物树干液流是光合有效辐射、饱和水气压亏损、土壤水分等多个环境因子耦合作用的结果,但因受植物自身根系吸水、木质部输水效率、茎干储水等因素的影响,基于植物树干液流测定值所表征的蒸腾速率与环境因子间的变化动态存在不等的时间差,即为液流的时滞效应[4-5]。若忽略时滞效应,通过树干液流测定值及相关环境因子所构建的蒸腾耗水模型会产生高达30%的误差[6]。因此,开展植物液流对环境因子的时滞效应研究有助于建立更精准的区域耗水模型,对深入理解植物蒸腾作用与生态系统水量平衡间的关系具有重要意义[7-8]。
桉树(Eucalyptusspp.)因其速生性、轮伐期短,成为我国南方的重要战略树种之一[9]。海南中西部为我国热带半湿润半干旱特殊生境区,也是重要的桉树浆纸林栽植区[10]。【研究进展】桉树在海南的大面积种植,产生了巨大经济效益的同时,其蒸腾耗水及产生的水生态问题备受关注[11-12]。深入理解桉树蒸腾耗水特征,建立更精准的桉树蒸腾耗水模型有助于定量回答桉树栽植对区域水量平衡的影响。王志超等[9]关于雷州半岛尾叶桉蒸腾耗水的研究结果表明,尾叶桉年蒸腾耗水量占同期年降水量的34.2%,主要影响因子为相对湿度、饱和水汽压亏损和光合有效辐射等。周翠鸣等[13]关于广西尾巨桉液流特征的研究结果表明,光合有效辐射和饱和水汽压亏损是影响树木蒸腾的主要动力。【切入点】以往多数研究主要集中在桉树林蒸腾耗水特征及其与环境因子间的关系,较少考虑植物液流的时滞效应。通过研究桉树液流对环境因子的时滞效应,有助于提高蒸腾耗水模型精度,进一步明确桉树水分供给规律及其对区域水量平衡的影响[14]。【拟解决的关键问题】为此,本研究选取海南西北部儋州林场桉树人工林作为研究对象,采用Granier热扩散探针技术,运用时间错位对比法,分析桉树生长季(5—10月)液流与环境因子间的时滞效应,揭示不同月份下液流的时滞效应差异,理清桉树液流的时滞效应与树木生长状况、土壤含水率间的关系,旨在弥补以往桉树人工林蒸腾耗水研究的不足,以期为准确评价人工林蒸腾耗水效应对区域水量平衡的影响提供科学依据。
1 材料与方法
1.1 试验地概况
研究地位于海南省儋州市西北部的儋州林场(19°43′3″N,109°20′3″E),该区域属热带季风气候,多年平均气温为23.8 ℃,年降水量为800~1 200 mm,年蒸发量为1 200~2 500 mm;干湿季分明,5—10月为雨季,降水量占全年降水量的70%~90%,且台风频繁,11月至次年4月为旱季,干旱少雨,季节性干旱严重;研究区地势平坦,以台地、平原为主,平均海拔33 m;土壤主要为砂质砖红壤,肥力较低。林场桉树栽植于2011年,株行距为1.5 m×3.0 m,主要栽种尾叶桉(Eucalyptus urophyllaS. T. Black)、细叶桉(Eucalyptus tereticornisSmith)和赤桉(Eucalyptus camaldulensisDehnh.)等种类,研究地林下植被稀疏,主要分布有飞机草(Eupatorium odoratumL.)、黄花稔(Sida acutaBurm. f.)和雀稗(Paspalum thunbergiiKunth ex Steud.)等植物。
1.2 材料选择
2016年5—10月在研究区随机建立一块桉树林固定样地,大小为40 m×30 m,样地内的桉树种类、林分密度,郁闭度、生长情况与研究地基本一致,同时对样地进行围栏围封。样地内总调查桉树植株100棵,平均树龄为6 a,平均胸径为9.35 cm,平均树高为12.2 m,按2 cm为1个径级单位在样地中选取处于生长期,长势良好、树干通直、无病虫害、人为影响因素小的5株桉树作为标准木;其中,8.0 cm径级植株较少,标准木为1株,10.0、12.0 cm径级标准木均为2株(表1)。样本的胸径范围7.52~12.80 cm,覆盖林场样方中78.10%的桉树胸径范围,基本可以代表研究区桉树总体样本数据。
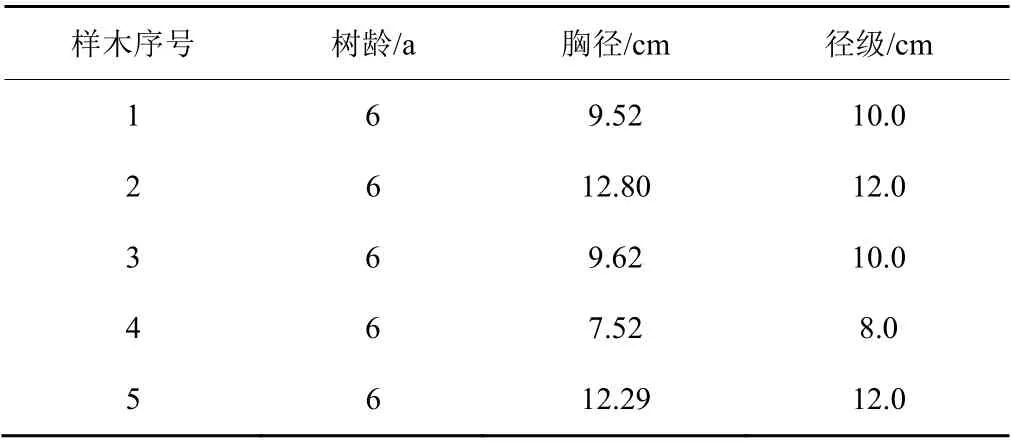
表1 桉树标准木树形特征Table 1 Type trees of Eucalyptus’ characteristics
1.3 树干液流测定
采用Granier热扩散探针技术(Dynamax Inc,USA)对样木树干液流进行连续监测。在样木的北向方位按照安装技术标准安装探针,在最外层裹上涂有反光材料(PVC)的防护罩,并用胶带固定,并用玻璃胶封口,以减少雨水、太阳辐射等外界因素对传感器的干扰;同时,为降低探针对树干液流传输的影响,本试验每2个月定期更换试验标准木,所更换标准木生长状况、树形特征等与前者基本相同。使用数据采集器收集和存贮热扩散探针的输出信号,数据采集时间间隔为30 s,每30 min记录1次平均值,每周定期下载数据。
1.4 环境因子测定
在距离观察点50 m处布设小型气候观测塔(Watch Dog 1000),连续监测林冠层附近的光合有效辐射(Photosynthetically active radiation,PAR)、大气温度(Ta)、相对湿度(Rh);0~60 cm土壤含水率(Soil water content,SWC)由林间布设的TDR采集,TDR探头分别随机埋藏于样地植株的行间与株间。本研究中的SWC是样地行间与株间0~10、10~30 cm和30~60 cm土层的SWC均值。饱和水汽压亏损(Vapor pressure deficit,VPD)由大气温度和相对湿度计算获得,计算式为[15]:
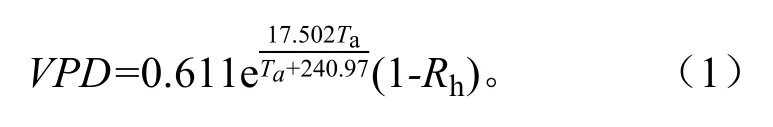
环境因子数据采集间隔与液流一致。
1.5 桉树液流速率测定
获得原始数据后,利用Dynamax Inc(USA)公司提供的数据处理软件对液流数据进行处理,液流速率计算式为[16]:
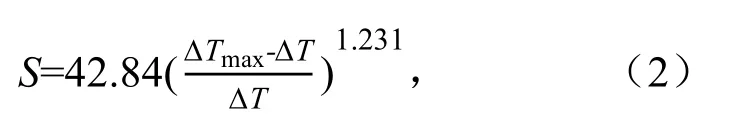
式中:S为树干中液流速率(mL/(cm2·h));ΔT为加热探针与数据传感探针之间的温度差值(℃);ΔTmax为空气湿度达到100%且为时长达48 h或树干直径停止变化、处于相对稳定状态时算得的最大温度差ΔT(℃)。
1.6 数据处理
本文选取生长季(5—10月)典型晴天白天的桉树液流数据进行分析,其中白天定义为PAR>5 μmol/(m2·s)的时段,并采用5月3日、7月1日和9月5日(全天晴天无云,PAR走势呈“单峰型”曲线)的数据的均值代表典型晴天下桉树液流速率的日变化特征,这是因为生长季内晴天正午时分天气为多云的时间较多,致午间PAR偏小,若采用整个研究期所有晴天的液流数据,其结果可能会掩盖实际的液流日变化特征。
采用时间错位对比法分析桉树与环境因子间的时滞效应,即按数据观测时刻顺序,建立各样木液流速率与对应PAR、VPD的数据列,并将液流速率分别与PAR、VPD逐次进行-4 h(提前4 h)或+4 h(滞后4 h)的错位移动,对错位后的数据进行线性相关性分析,将相关性系数最大时所对应的错位时刻确定为该环境因子的时滞时间[17]。
分别选取5、6月和9、10月典型晴天下的SWC数据,因5、6月降水较少,SWC较低,9、10月的降水较多,SWC较高,并要求5、6月所选时间的SWC均值低于SWC月均值,9、10月所选时间的SWC均值高于SWC月均值,以此分析SWC对桉树液流与环境因子间时滞效应的影响,其中5、6月共选取20 d,9、10月共19 d。5、6月20 d的SWC均值为10.45%,代表SWC低值时段;9、10月19 d的SWC均值为15.42%,代表SWC高值时段。低值时段与高值时段的SWC存在极显著差异(P<0.001),而5株样木胸径均值无显著差异(P>0.05)。
采用偏相关分析桉树液流速率与不同环境因子间的关系,单因子方差分析SWC、径级对液流速率与环境因子间时滞效应的影响水平。利用Dynamax Inc(USA)公司提供的数据处理软件进行数据预处理,使用SPPS 23.0软件进行数据的统计与分析,采用Excel 2010作图。
2 结果与分析
2.1 桉树液流速率与环境因子日变化
典型晴天(5月3日、7月1日和9月5日),桉树液流的日变化特征与PAR、VPD相似(图1),均存在明显昼夜节律,日间数值明显高于夜间,白天液流日变化特征表现为不规则“双峰型”曲线,说明桉树午间可能存在“光合午休”。日间液流启动时间为06:30—07:00,随后液流速率与PAR、VPD不断上升,但不同参数抵达峰值时间存在差异,液流速率抵达峰值时间为11:30,与PAR(12:30)、VPD(12:30)相比均提前60 min。另外,整个研究期间晴天桉树液流速率抵达峰值时间与环境因子间亦存在较大差异,其中液流速率抵达峰值时间平均为11:50,分别较PAR和VPD提前约35 min和75 min,这说明桉树液流对环境因子的响应存在时滞,且对不同环境因子的响应敏感度有较大差异。
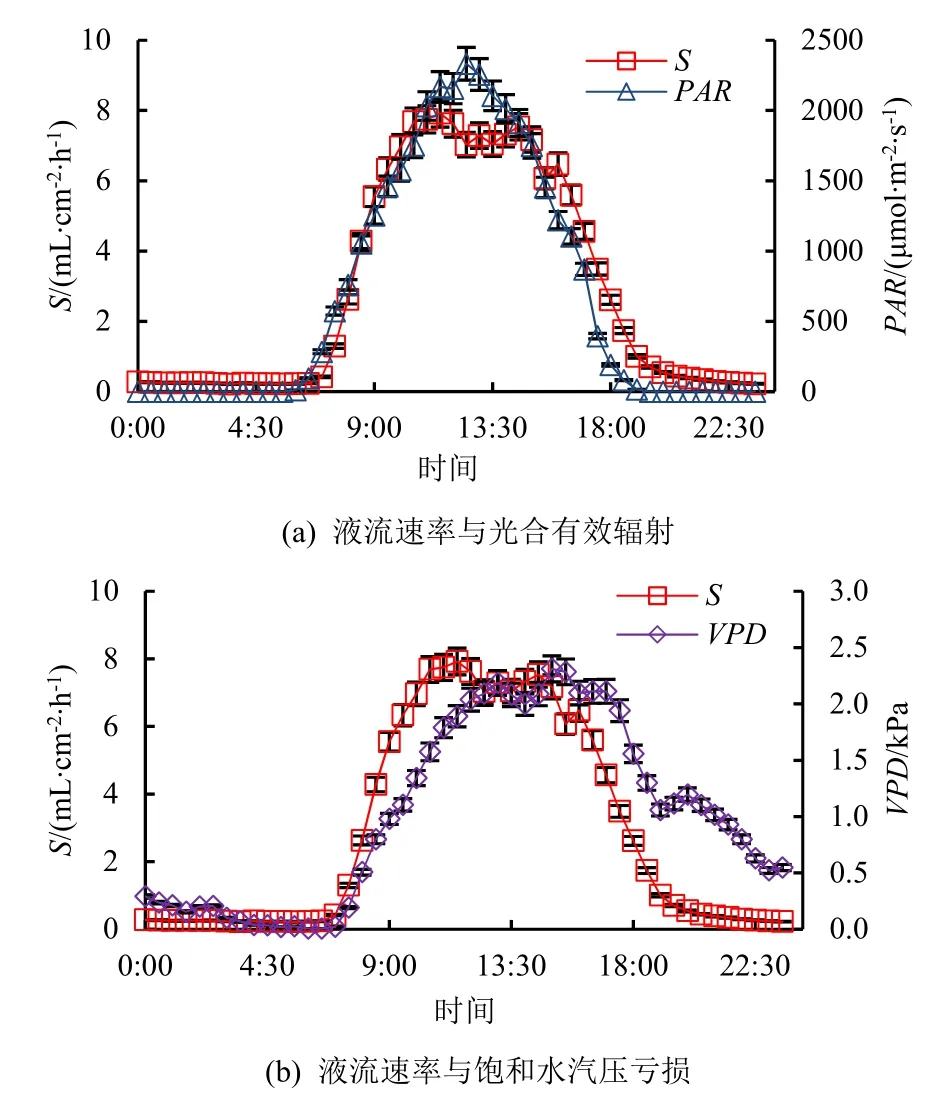
图1 典型晴天下桉树液流速率与环境因子日变化Fig.1 Diurnal variation characteristic of Eucalyptus sap flow density and environmental factors in a typical sunny day
液流速率与PAR、VPD的相关性均达到极显著水平(P<0.001),其中液流速率与PAR的相关性(0.876)高于VPD(0.650),说明日尺度上影响桉树液流速率的主要环境因子为PAR和VPD,其中最重要的为PAR。
2.2 生长季桉树液流对环境因子的时滞
典型晴天,桉树液流速率与PAR和VPD间呈“迟滞回环”现象(图2),即随着时间推移,当环境因子对液流的驱动作用逐渐增强时,液流速率持续升高一段时间后不再升高;而当环境因子的驱动作用逐渐减弱时,液流速率衰减的比例和原本增强的比例不一致,导致午后液流速率的衰退路径与上午液流速率增强路径存在差异。本研究中,桉树液流与PAR构成的迟滞回环呈逆时针,表明相同PAR下,上午液流速率低于下午;液流与VPD构成的迟滞回环呈顺时针,说明在相同VPD情况下,上午液流速率高于下午。不同时段液流速率与环境因子间表现出对称性,表明一天内上午和下午桉树液流速率对相同环境因子的响应敏感度存在较大差异,也说明桉树液流速率对PAR、VPD的响应存在时滞性。
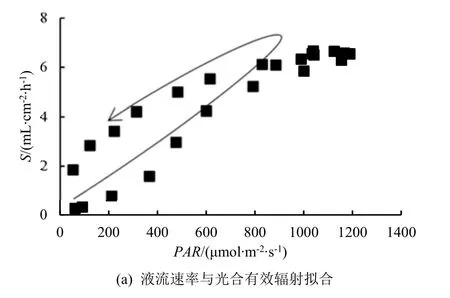
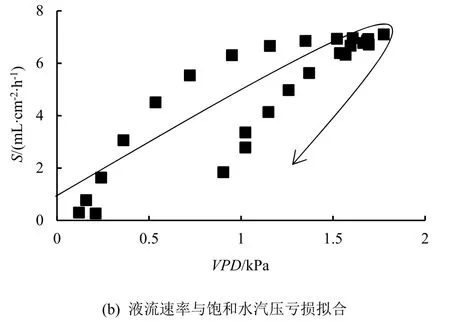
图2 生长季白天桉树液流速率与环境因子的曲线拟合Fig.2 Curve fitting of sap flow density with environmental factors of Eucalyptus on the daytime in growing season.
采用时间错位对比法分析桉树液流速率与不同环境因子间的时滞长度(图3),结果显示,整个观测期液流速率对PAR和VPD的响应均存在不同时长的时滞。桉树液流对PAR的时滞平均为-30 min(提前30 min);其中5、8月液流与实时的PAR相关系数最高,最高值平均为0.897,6、7月液流与PAR间的相关系数最高值平均为0.755,此时液流对PAR的响应提前约30 min,9、10月液流与PAR间的相关系数最高值平均为0.944,此时液流对PAR的响应提前约60 min;观测期桉树液流对VPD的时滞平均为-50 min;其中5、9月液流与VPD间的相关系数最高值平均为0.775,此时液流对VPD的响应提前约30 min,6、7、8、10月液流与VPD间的相关系数最高值平均为0.874,此时液流对VPD的响应提前约60 min。不同月份桉树液流速率与同一环境因子间时滞长度存在差异,相同月份不同环境因子间时滞长度亦有不同,表明存在某种变量影响桉树液流与环境因子间的时滞,且不同环境因子对该变量的响应程度存在差异。
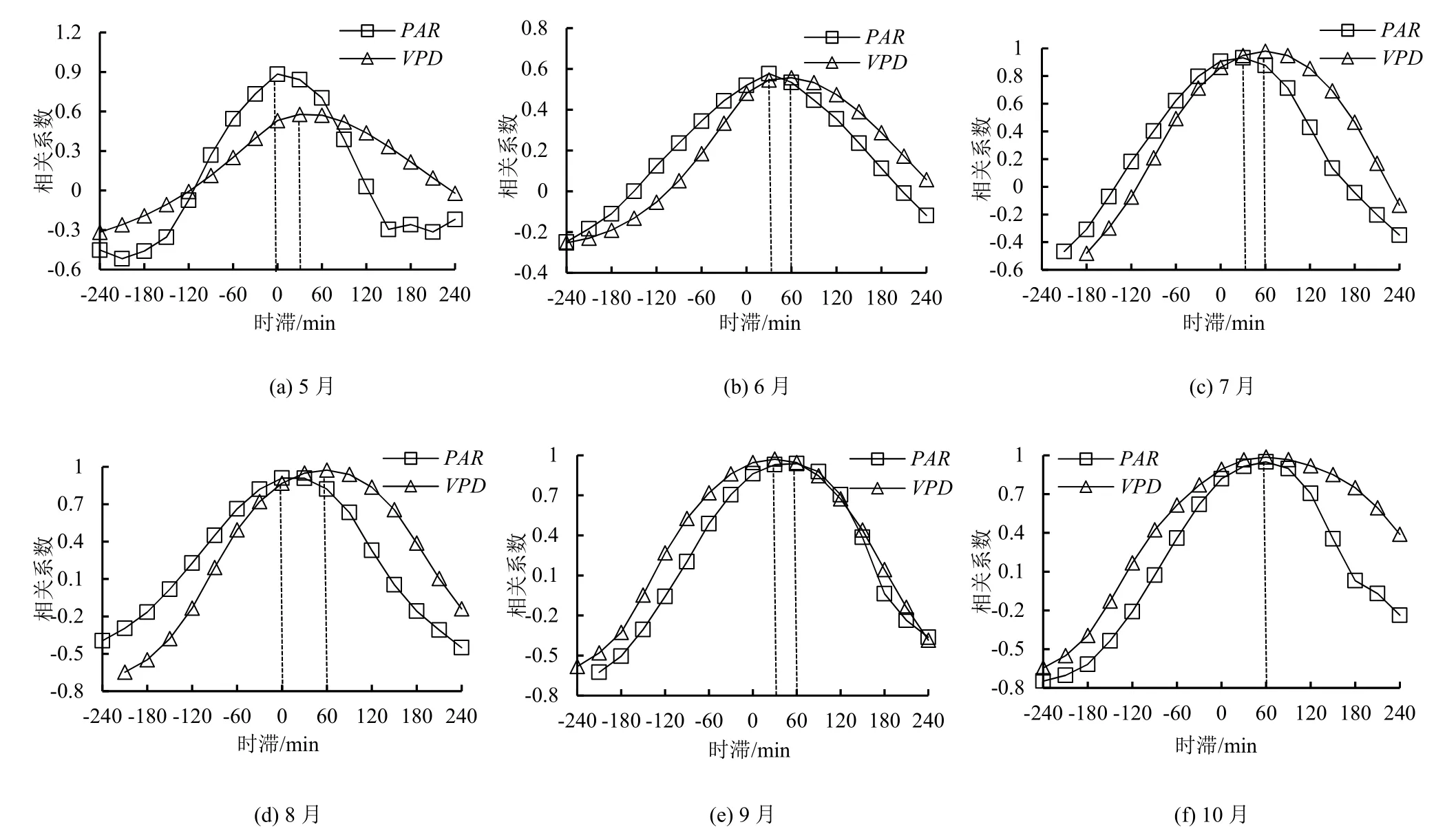
图3 生长季不同月份桉树液流速率与环境因子间相关系数变化Fig.3 Correlation coefficients of the sap flow density with environmental factors of Eucalyptus in growing season
2.3 土壤含水率和径级对桉树液流时滞效应的影响
采用时间错位对比法分析了SWC对液流速率与环境因子间时滞效应的影响(图4)。结果表明,在SWC低值时段(5、6月),液流速率与实时(时滞为0 min)的PAR间的相关系数最高,值为0.857;该时段液流速率与VPD间的相关系数最高值为0.847,此时对VPD的时滞为-60 min(提前60 min);在SWC高值时段(9、10月),液流速率与PAR间的相关系数最高值为0.948,此时液流对PAR的时滞为-60 min;而该时段液流速率与VPD间的相关系数最高值为0.964,此时液流对VPD的时滞为-30 min。One-way ANOVA分析结果显示(表2),对于2个环境因子(PAR和VPD)而言,在不同土壤水分条件下的液流速率对相同环境因子的时滞长度差异均达到显著性水平(P<0.05),但PAR(0.049)显著性水平较VPD(0.026)差,说明桉树液流与PAR、VPD间的时滞效应对SWC的响应存在差异。SWC高值时段的液流速率对PAR的时滞长度大于低值时段,VPD则相反,这表明SWC的大小会影响液流速率对蒸腾驱动因子的敏感度,SWC高值时段桉树液流对VPD的敏感性更高,而低值时段对PAR的敏感性较高。
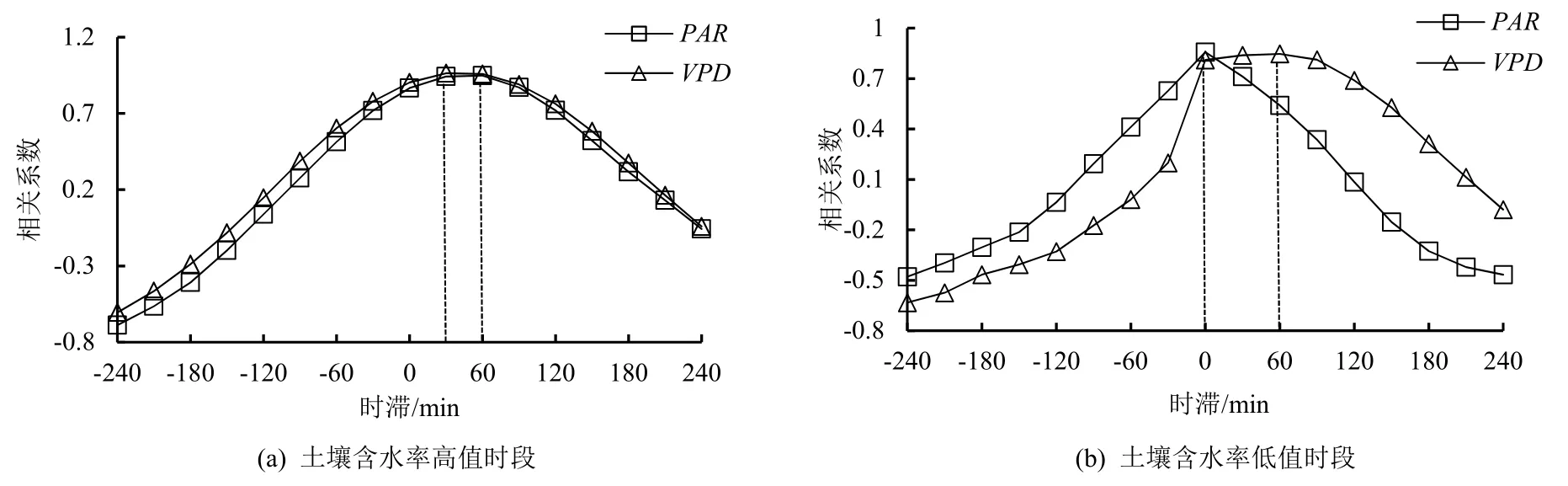
图4 不同土壤含水率条件下桉树液流速率与环境因子间相关系数变化Fig.4 Correlation coefficients between sap flow density and environmental factors of Eucalyptus under different soil water contents
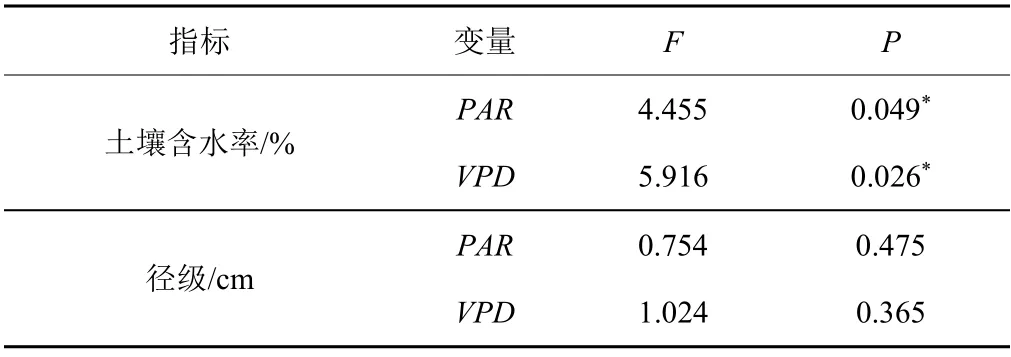
表2 土壤含水率和径级对桉树液流速率与环境因子间时滞效应影响程度的单因素方差分析(n=17)Table 2 Factor analysis of variance for the effects of soil water content and diameter classes on the time lag between sap flow density and environmental factors in Eucalyptus
由图5可知,不同径级样木的液流速率与PAR间的相关系数最大值差异较小(0.830~0.847),且液流速率对PAR的时滞均为-30 min;但不同径级样木液流速率对VPD的时滞长度存在差异,8、12 cm径级样木的液流速率对VPD时滞均为-30 min,相关系数最大值分别为0.741、0.764,而10 cm径级样木液流速率与实时(时滞为0 min)的VPD相关性最强,系数最高为0.817。One-way ANOVA分析结果显示(表2),径级对桉树液流时滞效应的影响未达显著性水平(P>0.05),说明径级对桉树液流与环境因子间的时滞效应影响较小。


图5 生长季各径级桉树液流速率与环境因子间相关系数变化Fig.5 Correlation coefficients between sap flow density with environmental factors of Eucalyptus under different diameter classes in growing season
3 讨 论
3.1 桉树液流的时滞效应
学者们研究指出,大部分植物树干液流与环境因子(PAR、VPD)间均存在时滞效应,且液流速率滞后于PAR、提前于VPD的情况较多。孙迪等[18]的研究结果表明,杨树树干液流速率滞后于净辐射约10~40 min,提前于VPD约50~120 min。赵春彦等[19]对胡杨(Populus euphratica)树干液流的研究表明,生长季液流速率滞后于太阳辐射约60 min,提前于VPD约120 min;姚增旺等[20]对生长季梭梭(Holoxylon ammodendron)液流的研究发现,液流速率滞后于PAR约80 min,提前于VPD约114 min;Wang等[21]也报道了青杨(Populus cathayana)树干液流速率滞后于太阳辐射,提前于VPD的现象。但也有部分研究表明液流速率提前于PAR和VPD。杨洁等[14]对黄土丘陵区辽东栎(Quercus liaotungensis)树干液流时滞效应研究发现,辽东栎液流速率分别提前于PAR、VPD的时间约为12.4、118.2 min;结果指出,生长季典型晴天桉树液流速率提前于PAR、VPD约60 min,这可能与桉树液流存在“光合午休”有关。桉树光合日变化呈“双峰型”曲线,且第1个液流峰值高于第2个,第1个峰值出现时间约在11:00—12:00间,早于PAR峰值出现时间(12:00—13:00),进而导致液流速率滞前于PAR。另外,偏相关结果表明,液流速率与PAR(0.876)的相关性高于VPD(0.650),说明桉树液流速率对不同环境因子的响应存在差异,PAR对桉树液流速率的影响更大。
3.2 桉树液流时滞效应的形成
有关植物液流时滞效应的发生机制主要包括夜间蒸腾储存水和树干与叶片间的运输距离延迟2种[22-23]。夜间蒸腾指植物夜间液流分为蒸腾与补水二部分,其中树干夜间补水有助于清晨植物冠层蒸腾作用的启动,进而影响植物液流的时滞效应,而夜间补水量大小主要与土壤含水率有关,即土壤含水率会影响植物的时滞效应[22]。树干与叶片间的运输距离延迟是指植物树干内部的水分通过导管运输至叶片,再通过气孔渠道返回大气的过程存在一定时间的延迟,导致了植物液流的时滞效应[24]。Granier等[25]的研究指出,植物体内水分的吸收和释放是针叶树树干液流产生时滞效应的主要原因。Chuang等[26]的研究也指出植物体内的储存水是导致液流与环境因子间产生时滞效应的主因。然而,Gonzalez-Benecke等[27]的研究表明,长叶松(Pinus palustris)和湿地松(Pinus elliottii)的树干储水量对液流的时滞效应影响有限。本研究结果显示,不同土壤水分状况下桉树液流与环境因子间的时滞呈显著差异(P<0.05),且当土壤含水率较高时桉树液流速率对PAR的时滞长度较大(-60 min),明显高于土壤含水率较低时的时滞长度(0 min),这说明桉树液流时滞效应可以用夜间蒸腾储存水的作用来解释。夜间桉树通过液流补水来弥补日间蒸腾所引起的水分亏缺,并在体内储存了一定量的水分来促进第二天清晨蒸腾作用的启动;同时土壤含水率的升高有利于植物对水分的吸收和传输,使夜间补水量上升,体内储存水含量亦升高,日间液流峰值的提前抵达,进而引起日间液流时滞长度的延长。当土壤含水率较低时,植物夜间补水不足,体内储水量较低,日间液流峰值则抵达较晚,故而缩短了液流速率与PAR间的时滞长度。然而,本研究区桉树夜间液流是否存在补水现象,以及影响夜间补水的主要因子有哪些,有待进一步研究。
3.3 桉树液流时滞效应的影响因素
前人研究表明,植物液流的时滞效应会受到生物与非生物因素的影响,其中生物因素主要包括胸径大小、树种类型、气孔导度等,非生物因素主要有土壤含水量、气候和季节等[14,28-30]。Goldstein等[31]研究指出,树木的大小对植物的储水量有重要影响,相同条件下,大树储存的水分较小树多,进而延长植物液流速率与环境因子间的时滞长度。Horna等[32]对热带雨林7个树种液流速率的研究表明,液流速率与环境因子间的时滞长度与树形特征成正比。王华等[33]对北京城区常见的树种树干液流时滞效应的研究指出,树干液流时滞长度和胸径、树高、边材面积及夜间补水量正相关,但与树种无关。杨洁等[14]关于黄土丘陵区辽东栎和刺槐树干液流时滞效应的研究结果显示,树种会影响液流与环境因子间的时滞长度,日尺度下辽东栎液流速率峰值提前于PAR约12.4 min,而刺槐则滞后于PAR约68.5 min。但也有部分研究结果与之相反。Phillips等[6]的研究结果显示,植物液流时滞长度与树高和树冠的暴露程度无关。姚增旺等[20]研究也表明,树形因子(地径、50 cm处直径、树高等)并不能解释梭梭树干液流的时滞特征。本研究结果表明,径级不能解释生长季桉树液流的时滞长度的变化,这可能是因为研究区日间光合有效辐射过强(午间PAR可达2 400 μmol/(m2·s)),即使夜间不同径级的树木补水量存在差异,大树虽夜间补水、体内储存水较多,但日间蒸腾耗水速率亦较快,小树虽夜间补水、体内储存水较少,但日间蒸腾耗水速率亦较慢[30]。因此,日间不同径级树木消耗夜间储存水的速度(单位时间内消耗的储存水的比例)差异较小,进而缩小了不同径级树木的液流速率与环境因子间的时滞长度。
4 结 论
1)生长季(5—10月),热带桉树树干液流日变化特征呈“双峰型”曲线,存在光合午休;影响桉树液流日变化特征的主要环境因子为PAR和VPD。
2)日尺度下,桉树液流与PAR和VPD间存在“迟滞回环”现象,不同时段(上午和下午)液流速率与环境因子间关系表现出对称性;生长季桉树液流与环境因子间存在明显的时滞效应,分别提前于PAR和VPD约30 min和50 min,相同月份下液流与不同环境因子间时滞长短存在差异。因此,在研究桉树液流与环境因子间时滞效应时应对不同环境因子进行综合考虑。
3)桉树液流时滞效应的主要驱动因子为SWC,径级对桉树液流时滞效应的影响较小。SWC较高时液流速率对PAR时滞较长,而SWC较低时液流速率对VPD的时滞较长,但SWC对桉树液流时滞效应的影响机制尚不明确,未来有必要加强相关研究。
