工作记忆训练诱发的神经可塑性——基于系列fMRI实验的脑区分布递减时空模型*
2022-02-19陈幸明伏云发李恩泽ZHANGJian陈盛强党彩萍
陈幸明 付 彤# 刘 昌 张 宾 伏云发 李恩泽 ZHANG Jian 陈盛强 党彩萍,
·元分析(Meta-Analysis)·
工作记忆训练诱发的神经可塑性——基于系列fMRI实验的脑区分布递减时空模型*
陈幸明1付 彤1#刘 昌3张 宾1伏云发4李恩泽5ZHANG Jian6陈盛强2党彩萍1,2
(1广州医科大学附属脑科医院, 广州 510370) (2广州医科大学卫生管理学院, 广州 511436)(3南京师范大学心理学院, 南京 210024)(4昆明理工大学信息工程与自动化学院, 昆明理工大学医学院, 昆明 650031)(5南方医科大学南方医院精神心理科, 广州 510515) (6天津医科大学, 天津 300070)
工作记忆训练(Working Memory Training, WMT)诱发神经可塑性, 但其具体机制尚不明晰。为探索WMT改变正常人群大脑功能的时空特性, 以“扩展的智力顶额整合理论”和“神经效率假说”为依据, 采用逐层递进的5种方法, 分6个步骤来查究近20年来正常人群WMT的37篇fMRI文献。第一步, 用叙述性综述、频数分析和卡方检验法比较脑区激活模式和脑网络功能连接在WMT前后发生的改变, 发现WMT改变了大脑的5个联合区、7个宏观区和3个子区。其中, 额上回、顶下小叶和扣带回这3个子区各自激活减弱的报道文献数量多于其激活增强的, 且这种差异分别具有统计学意义。第二步, 采用激活似然估计法对其中26篇开展元分析, 发现大脑的3个子区激活减弱水平在WMT前后的差异具有统计学意义, 即额中回(BA6和8)、额上回(BA6)和前扣带回(BA24和32)。第三步, 综合定性和定量分析结果, 提出WMT脑区分布递减时空模型, 产生5个结果和讨论。第四步, 采用非参数检验进一步追踪WMT效应的调节因素, 发现训练的任务类型和时间分别对脑区激活的影响具有统计学意义。第五步, 针对正常人群WMT诱发神经可塑性的时空特性, 得出3个结论:第一, WMT改变了正常人群相应脑区的神经活动, 表现为减弱或增强, 但减弱更加突出, 且更新和较短时间的WMT倾向于诱发较多减弱; 第二, 这些神经活动变化主要发生在额顶叶联合区, 但也包括分别以颞叶、枕叶、扣带回及纹状体为主的联合区, 在一定范围内体现了整脑功能联合。这体现了WMT诱发神经可塑性的空间特性, 且符合“扩展的智力顶额整合理论”; 第三, 额中回、额上回、顶下小叶和扣带回(尤其前扣带回)这4个子区在激活减弱水平上重点展示了WMT神经可塑性的时间特性, 且符合“神经效率假说”, 恰好体现出“聪明的大脑更懒惰”。第六步, 指出WMT诱发神经可塑性的未来研究可能关注脑可塑性中的低活跃性、辨析额中回、额上回、顶下小叶和扣带回(尤其前扣带回)这4个子区在激活减弱水平上体现的时间特性、找寻训练减弱或增强大脑活动的综合性影响因素。
工作记忆训练, 神经可塑性, 脑区分布递减时空模型, 额中回, 额上回, 前扣带回
日益高压的社会环境和人口老龄化带来的认知老化问题使人们愈发关注认知绩效的提升。工作记忆(Working Memory, WM)作为一种对信息进行暂时贮存和加工的容量有限的记忆系统, 由于是人类高级认知功能的核心, 从而成为提升执行功能这类干预研究的一个关键靶点(Constantinidis & Klingberg, 2016)。而产生于21世纪早期、通过程序化WM任务来循序渐进提升WM容量及与之密切相关认知功能的训练活动即为工作记忆训练(Working Memory Training, WMT)。短期的WMT可以在个体发育成熟期过后继续诱发神经可塑性(neuroplasticity), 但那些变化呈现多样性, 甚至互相矛盾(Jolles et al., 2010; Schneiders et al., 2012)。因此, WMT的脑机制依然不够明晰(Salmi et al., 2019)。那么, 伴随WMT经验产生的神经可塑性, 其时间和空间特性是怎样的?
WM与智力密切相关, 故WMT神经可塑性的空间特性可追溯到智力的顶额整合理论(Parieto- Frontal Integration Theory, P-FIT) (Jung & Haier, 2007)。P-FIT被系列实验检验, 经微调和优化后, 发展为“扩展的顶额整合理论” (Extended PFIT, ExtPFIT) (Gur et al., 2020)。ExtPFIT将智力扩展到WM等高级认知活动, 关注高级认知与大脑结构和功能的多模态参数间的关系。它除了证明P-FIT明确的智力脑区, 即背外侧前额叶(the dorsolateral prefrontal cortex)、顶下和顶上小叶(the inferior and superior parietal lobule)、前扣带回(the anterior cingulate)、颞叶(temporal lobes)、枕叶(occipital lobes)以及包含弓状束的白质区(white matter region) (Jung & Haier, 2007), 还新增了纹状体(striatum)、边缘系统(limbic system)和小脑(cerebellar) (Gur et al., 2020)。那么, WMT改变的脑区是否也包含在ExtPFIT?故依据ExtPFIT和P-FIT, 假设WMT改变脑区的空间特点有:改变的是系列脑区, 而非某一局部; 系列脑区中, 额顶叶网络(frontal- parietal network, FPN)的效应可能最为突出; 系列脑区对WMT效应的体现不同, 呈递减分布。
ExtPFIT进一步显示, 较好的WM成绩连接着PFIT脑区内较高的脑容量、较大的灰质密度和较多的脑区激活(Gur et al., 2020)。但是, “神经效率假说” (neuro efficiency hypothesis)多次解释了“聪明的大脑更懒惰”。比如, 高智商人在推理时, 脑皮层激活却很少, 其神经元活动以一种稀疏而有效的方式组织(Genç et al., 2018)。依据这两种理论, 对WMT诱发神经可塑性的时间特性提出假设:训练减弱或增强了脑区活动, 二者并存于已有研究中, 但激活减弱是主导。
采用逐层递进的5种方法——叙述性综述(narrative review)、频数分析(frequency analysis)、元分析(meta-analysis)、建模(modeling)和非参数检验(non-parametric test), 分六步来查究近20年来正常人群WMT功能磁共振成像(fMRI)实验的37篇文献(42个实验), 集中比较WMT前后被试执行WM任务时的大脑活动, 从而探索WMT诱发的大脑功能可塑性(functional plasticity)在脑区的时空变化, 并进一步追溯可能影响其训练效应的调节因素。
1 WMT改变脑区活动的叙述性综述
1.1 WMT改变脑区激活模式
归纳32篇文献(36个实验, 见表1)后发现, WMT引起激活模式发生变化的主要脑区是分别以额叶(frontal lobe)、额顶叶(frontal-parietal lobe)、额顶颞叶(frontal-parietal-temporal lobe)、额顶枕叶(frontal-parietal-occipital lobe)和纹状体(striatum)为主的5个联合区, 它们在WMT后发生激活减弱或增强或重排。
1.1.1 WMT减弱或增强额叶联合区激活
短期的更新、抑制和言语类WMT减弱或增强了额叶为主联合区的激活。视觉更新训练减弱了成年人背外侧前额叶皮层的激活(Miró-Padilla et al., 2020); 听觉更新训练减弱了青年人右额下回(Schneiders et al., 2012)、老年人右额中回和尾侧额上沟(Heinzel et al., 2016)的激活; 抑制训练使青年人背外侧前额叶皮层激活缓慢减弱和前扣带回激活急剧下降 (Milham et al., 2003); 而言语WMT既减弱了青年人双侧背外侧前额叶、右额上回、右额极区和辅助运动区的激活水平(Jansma et al., 2001), 也增强了内侧前额叶皮层的激活(Jolles et al., 2010, 实验1)。这表明, WMT可能提高了视听觉模态和言语WM的神经效率。
1.1.2 WMT诱发额顶叶联合区激活减弱、增强或重排
顶叶与额叶联合参与WMT, 且短期的视听觉更新、空间和言语WMT会减弱或增强额顶叶为主的联合脑区激活。
视觉更新训练减弱了成年人右额中回和右后顶小叶(Schneiders et al., 2011, 实验1)、成年人双侧额上回及顶叶右侧缘上回和角回(Clark et al., 2017)以及老年人额顶叶的WM网络(Heinzel et al., 2014)激活; 听觉更新训练也减弱了大学生右额中回和右后顶小叶激活(Schneiders et al., 2011, 实验2); 言语WMT减弱了中年人额叶−顶叶−小脑回路和皮质下区域激活(Emch et al., 2019); 综合性WMT减弱了大学生左侧额顶叶脑区和左后侧脑岛激活(Takeuchi et al., 2014)。
但视空间和言语WMT增强了青年人前额叶和顶叶的激活(Olesen et al., 2004, 实验1)、以及额中回、额下回与顶叶上、下和内侧的激活(Westerberg & Klingberg, 2007); 视觉抑制训练增强了青年人额顶叶为主的一些核心抑制脑区的激活(Kelly et al., 2006); 言语更新训练增强了成年人的左额下回和右后顶叶皮层的激活(Beatty et al., 2015)。
另外, 一些视空间WMT诱发脑区激活减弱的同时也伴有激活增强, 产生激活重排(redistribution)。首先, 珠心算(Abacus-based mental calculation, AMC)训练增强了大学生额顶叶脑区持续激活, 但减弱其短暂激活。在记忆维持阶段, 额中回、顶上小叶激活增强, 但右侧顶下小叶、内侧前额叶、右前额叶和右眶额叶、后扣带回激活持续减弱; 在操作阶段, 双侧额下回、辅助运动前区等脑区激活短暂减弱, 但内侧前额叶上部激活增强(Zhou et al., 2019); 其次, 空间更新训练诱发青年人的右额下回和右顶内沟激活增强, 但训练4周后, 这些脑区却出现激活减弱, 呈倒U型变化(Hempel et al., 2004); 视空间WMT增强了青年人额中回、顶叶的上、下和内侧激活, 但减弱了右侧额下沟、扣带沟和左侧中央后回的激活(Olesen et al., 2004, 实验2); 视觉更新训练减弱了额上回激活, 但增强了左侧顶叶上部激活(Nęcka et al., 2021); 再次, 时间相对较长的视空间WMT减弱了青年人额叶(额中回、中央前回)、顶叶(顶下小叶、楔前叶)、前扣带回和枕叶的激活(Garavan et al., 2000, 实验2); 但是, 时间较短的WMT除了减弱以上脑区外, 还减弱了额下回、额上沟、顶内沟和后扣带区域的激活(Garavan et al., 2000, 实验1)。Garavan的这两个实验反映了激活位置在训练前后产生重组(reorganization), 也间接支持了激活重排。对比二者结果显示, 激活减弱更可能是神经效率提高的表现, 练习可能干扰训练效应(Garavan et al., 2000)。那么, 脑区激活减弱或增强是否受WMT时长调控?这有待进一步验证。
1.1.3 WMT减弱额顶颞叶联合区激活
颞叶联合额顶叶脑区参与自上而下的自适应加工活动, 具有较好的可塑性(Landau et al., 2007)。而短期的项目再认和更新WMT可减弱这三个脑区的激活。比如, 延迟面孔再认训练减弱了青年人的额、顶、颞叶皮层下区域激活(Landau et al., 2004); 延迟客体或空间位置识别训练减弱了青年人额叶内外侧、顶叶、颞叶(Landau et al., 2007)和左额中回、右顶下小叶、颞外侧、颞上叶皮层(Sayala et al., 2006)激活; 视觉n-back训练减弱了成年人额上/额中、顶下小叶、前扣带回和颞中皮层激活, 且在训练结束5周后, WMT效应依然稳定(Miró-Padilla et al., 2019); 视觉情绪双重n-back训练减弱了成年人腹外侧和背外侧前额叶、顶下小叶、颞叶以及扣带回激活(Schweizer et al., 2013)。这些变化反映了WMT较大范围地提高了神经加工效率。
1.1.4 WMT增强额顶枕叶联合区激活
枕叶作为视觉中枢, 较多参与视空间WMT, 它与额叶或顶叶联合被激活。时间相对较长的珠心算训练增强了小学生执行数字WM时诱发的额叶右侧辅助运动区、右侧后顶上小叶和枕上回激活(Li, Hu, et al., 2013), 以及执行视空间更新任务诱发的额、顶和枕叶激活, 并提示额中回是珠心算训练向视空间WM迁移的神经基础(Wang et al., 2019)。可见, WMT对枕叶的改变相对较少。
1.1.5 WMT诱发纹状体为主联合区的激活减弱、增强或重排
纹状体作为大脑基底核之一, 参与更新活动, 以它为主的系列脑区同样受WMT影响。转换训练减弱了老年人的纹状体激活(Dörrenbächer et al., 2020); 视空间和言语更新训练减弱了青年人纹状体和海马的激活(Flegal et al., 2019)。
但是, 视空间、更新和言语WMT还可增强纹状体为主的额顶颞枕叶脑区激活。视听觉双重n-back训练(Salminen et al., 2016)和视空间WM广度训练(宋杰等, 2020)都增强了纹状体激活; 视觉更新训练增强了老年人的左侧纹状体以及左额叶、双侧顶叶、右颞叶和左小脑的激活(Dahlin et al., 2008, 实验1); 言语WMT增强了青年人的纹状体激活, 且较小程度增强了左外侧前额叶、双侧背外侧前额叶和左顶上叶皮层的激活(Jolles et al., 2010, 实验2)。
而且, WMT同时诱发纹状体与额顶枕颞叶发生激活重排。数字和空间更新训练先增强青年人的双侧纹状体(具体在壳核)激活, 随后减弱右侧纹状体激活(Kühn et al., 2013); 视觉更新训练增强青年的左右侧纹状体、右枕叶和右颞叶激活, 但也减弱了其右侧额叶和右侧顶叶激活(Dahlin et al., 2008, 实验2)。
1.1.6 WMT改变脑区激活的小结
可见, WMT主要诱发分别以额叶、额顶叶、额顶颞叶、额顶枕叶和纹状体为主的五个联合区发生激活改变。较多脑区的参与, 充分支持了WM的训练效应, 在一定范围内体现了整脑功能联合。而且, 脑区激活的减弱或增强现象并存于已有实验报告中。另外, 个体在训练中逐渐学会新的记忆策略, 在使用新记忆策略前后, 大脑可能产生激活重排或功能重组, 它可能体现了脑区产生的一般性加工和特异性加工(Buschkuehl et al., 2012)。那么, 在时间和空间上, WMT对脑区的改变表现在哪些典型性脑区和脑区的典型性变化上?
1.2 WMT改变脑网络功能连接
WMT会改变脑内自发产生的高振幅、低频率BOLD信号间的时间相关, 即功能连接(Functional Connectivity, FC), 且主要发生在静息态或任务态下的两个网络(见表2)。
1.2.1 WMT增强额顶叶网络的FC
额顶叶网络(frontal-parietal network, FPN)包含2个与WM联系密切的网络, 即执行控制网络(executive control network, ECN)和背侧注意网络(dorsal attention network, DAN) (Thompson et al., 2016)。而更新、言语、视觉记忆、数学计算和珠心算的WMT会增强这些网络的FC。比如, 视听觉双重更新训练增强了青年人执行2-back任务激活的额顶叶兴趣区的FC, 即ECN节点间、DAN节点间、以及两个网络之间的FC, 且这种连通性的增强程度与WM成绩改善程度呈正相关(Thompson et al., 2016); 言语WMT增强了老年人FPN及默认模式网络和WM网络内部的FC (Iordan et al., 2020); 数学计算和视觉WMT增强了注意网络的FC, 具体为右额中回与右颞叶脑区节点的FC, 且这种FC与抑制控制分数提高密切相关(Sánchez- Pérez et al., 2019); 珠心算训练增强了儿童右侧额下回和右侧辅助运动区间的FC (Li, Hu, et al., 2013)。
1.2.2 WMT改变默认模式网络的FC
有些脑区在静息态下比在任务态下产生更强的自发性活动, 常处于任务诱导失活状态, 从而构成一个支持脑功能默认模式的网络, 即默认模式网络(default mode network, DMN)。而WMT可改变DMN的FC。首先, 言语WMT减弱了青年人DMN网络与颞叶脑区的FC, 即内侧前额叶皮层和右侧后颞中回间的FC, 且DMN的FC减弱程度与WM行为成绩的提高幅度呈负相关(Jolles et al., 2013, 实验1); 其次, WMT既减弱DMN与额顶叶脑区的FC, 又增强DMN内部网络的FC。视听觉WMT减弱了青年人外围注意系统关键节点间的静息FC, 即内侧前额叶与右侧后顶叶/右外侧前额叶的静息FC, 同时增强了DMN重要节点间的静息FC, 即内侧前额叶与楔前叶之间的静息FC (Takeuchi et al., 2013); 再次, WMT未改变儿童DMN的FC (Jolles et al., 2013, 实验2), 表明WMT诱发DMN的FC产生变化具有年龄依赖性。
1.2.3 WMT改变脑网络FC小结
WMT不仅改变了多个脑区激活, 也改变了额顶叶和默认模式网络内神经网络的连接状态。这体现了WMT对脑网络FC的可塑性, 也提供了WMT改善神经水平的潜在信号。而且, FC变化与激活模式变化对应的脑区总体上具有一致性, 均以额顶叶网络为WMT诱发的主要脑区。减弱和增强这两个相反的现象分别存在于文献中, 这可能涉及到具体脑区对WMT敏感性差异。但是, 这些差异的统计检验效应如何?这有待下一步探索。
2 WMT改变脑区活动的频数分析和卡方检验
依据WMT诱发脑区神经活动的减弱和增强维度来重新分析文献。
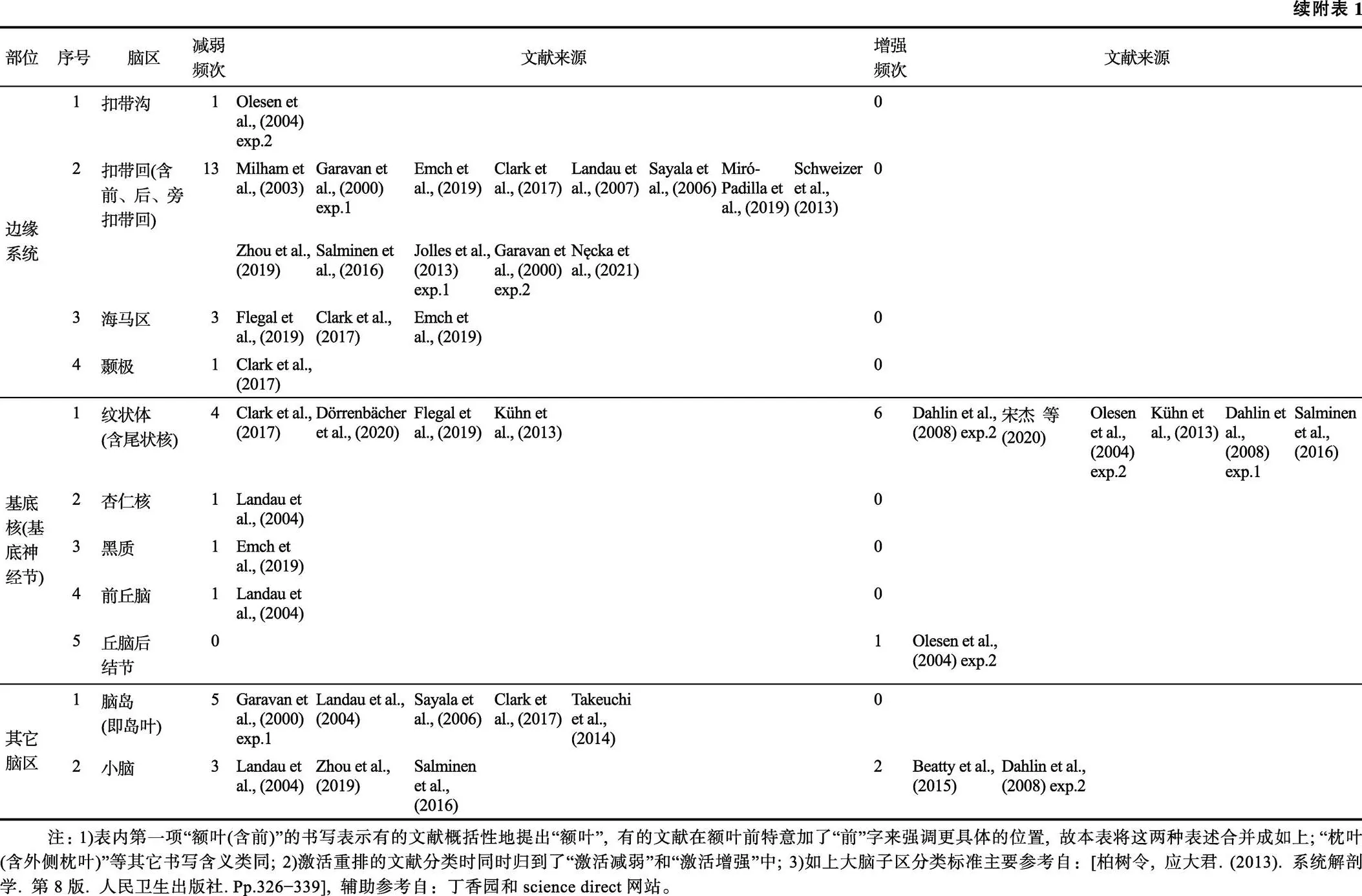
首先, 对37篇文献(42个实验)做频数分析, 结果(见网络版附表1)显示, WMT诱发7个宏观脑区发生激活减弱或增强(分别含重排), 即额区、顶区、颞区、枕区、边缘系统、基底核及其它脑区, 它们包含的大脑子区共有44个。具体而言, 第一, WMT改变了额区的15个子区, 即额叶(含前额叶)、背外侧前额叶、腹外侧前额叶、内侧前额叶、外侧前额叶、额上回(含额内回)、额中回、额下回、中央前回、辅助运动区(含前运动皮层)、额极、额下沟、额上沟、额叶眼动区、眶额皮层; 第二, WMT改变了顶区的6个子区, 即顶叶、顶内沟、顶上小叶、顶下小叶(含缘上回、角回)、中央后回、楔前叶; 第三, WMT改变了颞区的6个子区, 即颞叶、颞外侧、颞上回、颞中回、颞下回、梭状回; 第四, WMT改变了枕区的6个子区, 即枕叶(含外侧枕叶)、枕上回、枕中回、枕下回、楔叶(Cuneus)、距状回(Calcarine gyrus); 第五, WMT改变了边缘系统的4个子区, 即扣带沟、扣带回(含前、后、扣带旁回)、海马区、颞极; 第六, WMT改变了基底核(基底神经节)的5个子区, 即纹状体(含尾状核)、杏仁核、黑质、前丘脑、丘脑后结节; 第七, WMT改变了其它脑区的脑岛(即岛叶)和小脑这2个子区。
其次, 从中筛选出8个出现次数大于等于9的子区(来自28篇文献, 32个实验), 即:额中回(17)、额上回(10)、额下回(11)、顶下小叶(17)、顶上小叶(11)、顶内沟(9)、扣带回(13)和纹状体(10), 且除额上回和扣带回仅表现为激活减弱而没有增强外, 其余脑区均减弱和增强并存(见表3和图1)。而且, 这8个脑区减弱与增强的比率差异具有统计学意义(χ2= 18.35,= 7,= 0.010)。
最后, 用卡方检验分别检验大脑的8个子区激活减弱和增强比率的差异性(见表4), 发现顶下小叶的差异具有统计学意义(χ2= 4.77= 1,= 0.049), 其激活减弱次数(13)显著多于激活增强次数(4)。结合额上回和扣带回仅表现为激活减弱次数较多(大于10)而增强次数为0, 可得结论:WMT诱发额上回、顶下小叶和扣带回这3个脑区的激活减弱明显多于其激活增强。
可见, 如上分析充分支持了ExtPFIT提到的脑区。而且, 脑区激活减弱和增强分别符合“神经效率假说”和ExtPFIT, 且“神经效率假说”得到更多支持。
3 WMT改变脑区活动的元分析
3.1 方法
3.1.1 文献检索和纳入标准
限定文献检索的时间周期为2000年1月1日到2021年3月20日, 内容为在标题或摘要中同时包含关键词“fMRI”和“working memory training”或“training working memory”。以此为条件在Pubmed上检索出73篇文章。为避免遗漏, 在2篇综述文献(Buschkuehl et al., 2012; Constantinidis & Klingberg, 2016)中追溯找到16篇, 故合计89篇。
以如下标准对文献做“异质性”处理, 保证数据来源和统计指标的一致性。筛选可纳入的文献法为:(1)文章为实证研究; (2)研究内容为WMT的训练效应; (3)研究人群为健康人群; (4)实验技术采用fMRI; (5)研究指标为全脑分析WMT前后差异的峰值激活脑坐标; (6)研究的空间坐标体系为MNI或Talairach。据此得到26篇。其中, 2000~2010年的有10篇; 2011~2015年的有7篇; 2016~2021年的有9篇。被试总数为614人, 坐标数量为294, MNI坐标系为25篇, Talairach坐标系为6篇。逐步排除文献数量及标准见图2, 纳入文献数据详见表5。
3.1.2 元分析算法
采用激活似然估计法(Activation Likelihood Estimation, ALE) (Turkeltaub et al., 2012)检验fMRI数据反映的WMT前后脑激活水平差异。先依据特定优化算法, 对文献中的脑空间坐标进行自动计算, 然后采用置换检验或多重比较校正进行推断统计。
3.1.3 工具与数据处理
采用GingerALE 2.3.6 (http://www.brainmap. org/ale/)软件处理数据、Mango 4.1 (http://ric.uth- scsa.edu/mango)软件呈现结果图像。若文献结果为Talairach坐标, 则使用icbm2tal函数将其转为MNI坐标(Lancaster et al., 2007)。最后, 全部文献的元分析都在MNI坐标系进行。参考Ginger ALE手册提供的算法和参数设置建议, 选择FDR pID算法, 临界统计值为0.05, 最小体积为300 mm (Eickhoff et al., 2009)。
3.2 结果和讨论
如表6和图3所示, 对WMT后脑区激活增强的文献做元分析, 结果无显著激活簇。但对WMT后脑区激活减弱的文献做元分析, 得到激活减弱的两个激活簇, 一个是以MNI坐标(x = 36, y = 20, z = 40)为中心的额中回(Middle Frontal Gyrus, MFG) (100%)的区域, 体积为352 mm3; 另一个是以MNI坐标(x = –2, y = 12, z = 52)为中心、包含额上回(Superior Frontal Gyrus, SFG) (93.3%, BA6, 其中额内回占该激活簇39.2%)和腹侧前扣带回(Anterior Cingulate Cortex, ACC) (6.8%)的区域, 体积为1024 mm3。可见, 在激活减弱水平上, WMT前后差异具有统计学意义的脑区为额中回、额上回和前扣带回。
由此表明, 已有文献汇报的WMT增强神经活动的具体脑区差异较大。当对已有统计结果做再统计时, 那些已报告的具体脑区由于异质性太大而未能显示出更稳定的空间特性。但是, WMT减弱神经活动的具体脑区, 却在这种荟萃分析中凸显出来。可见, 这间接支持了观点:认知训练使人在执行相关任务过程中消耗更少的能量、诱发更少的大脑活动。
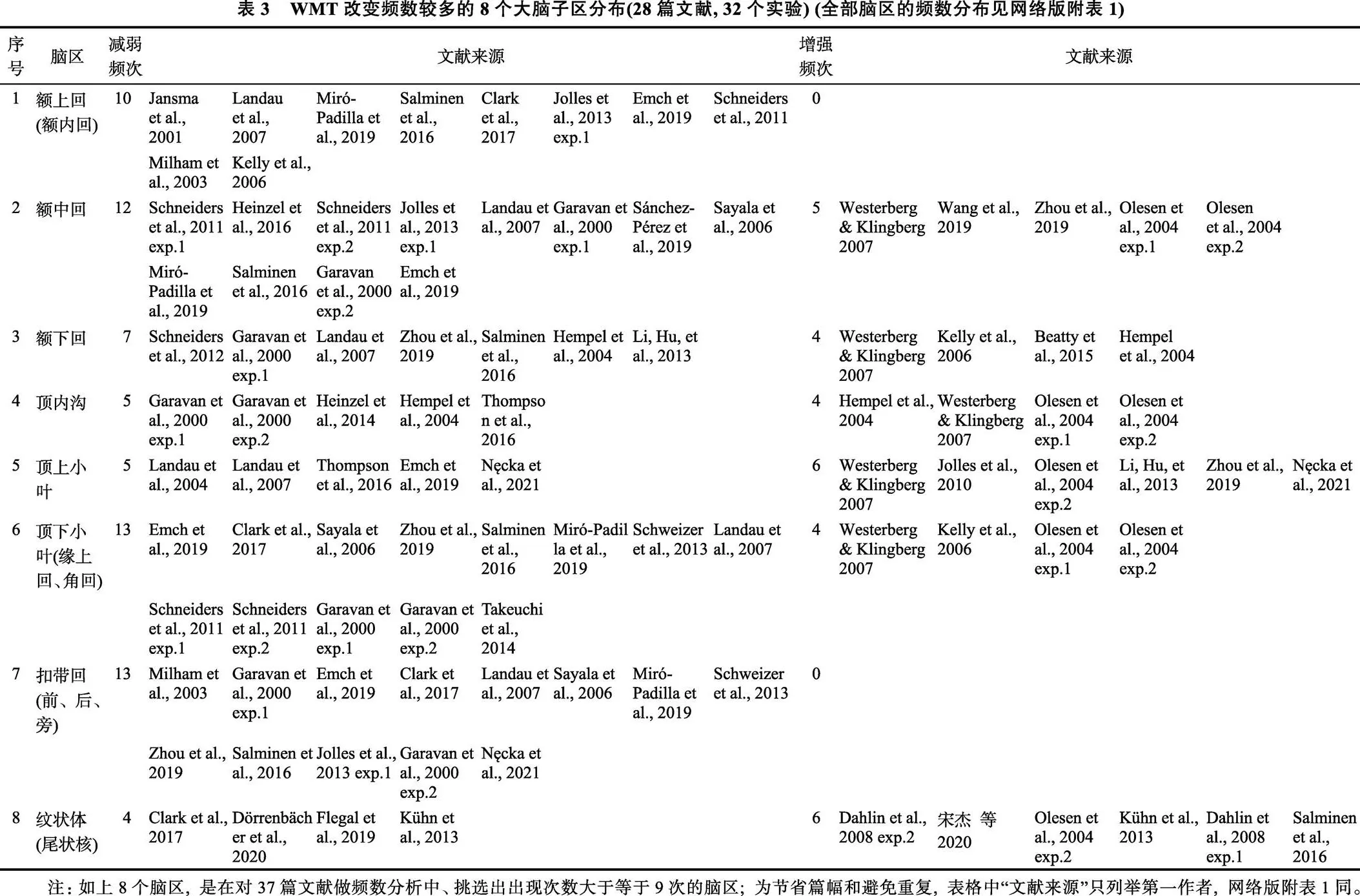
图1 WMT诱发神经活动减弱与增强的具体脑区频数分布(37篇文献, 42个实验)
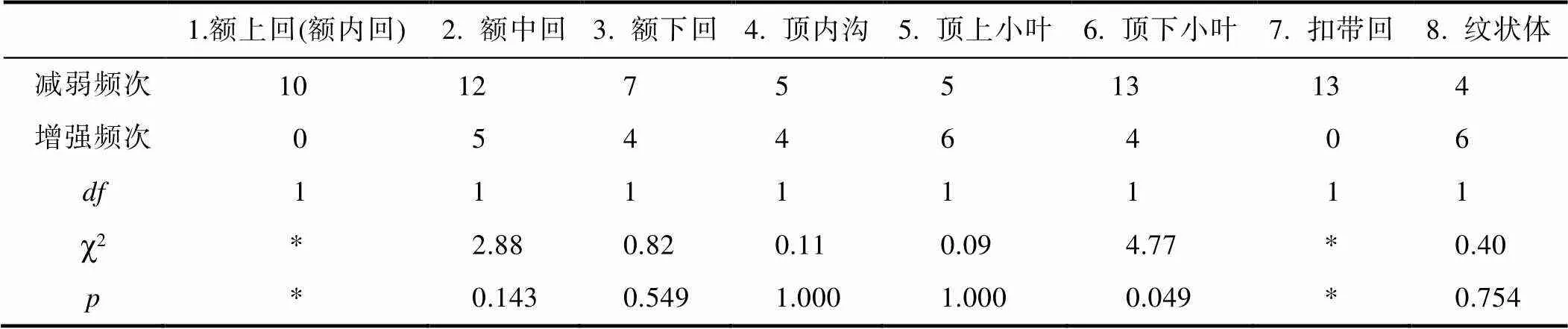
表4 WMT诱发大脑子区改变的卡方检验(28篇文献, 30个实验)
注:第1和7脑区存在单元格频数为0情况, 用*表示无法进行卡方检验; 其余脑区存在样本量<40且单元格期望频数小于5并大于1的情况, 故采用Fisher校正检验。参考自:[孙振球, 徐勇勇. (2010).. 人民卫生出版社. p.98]。
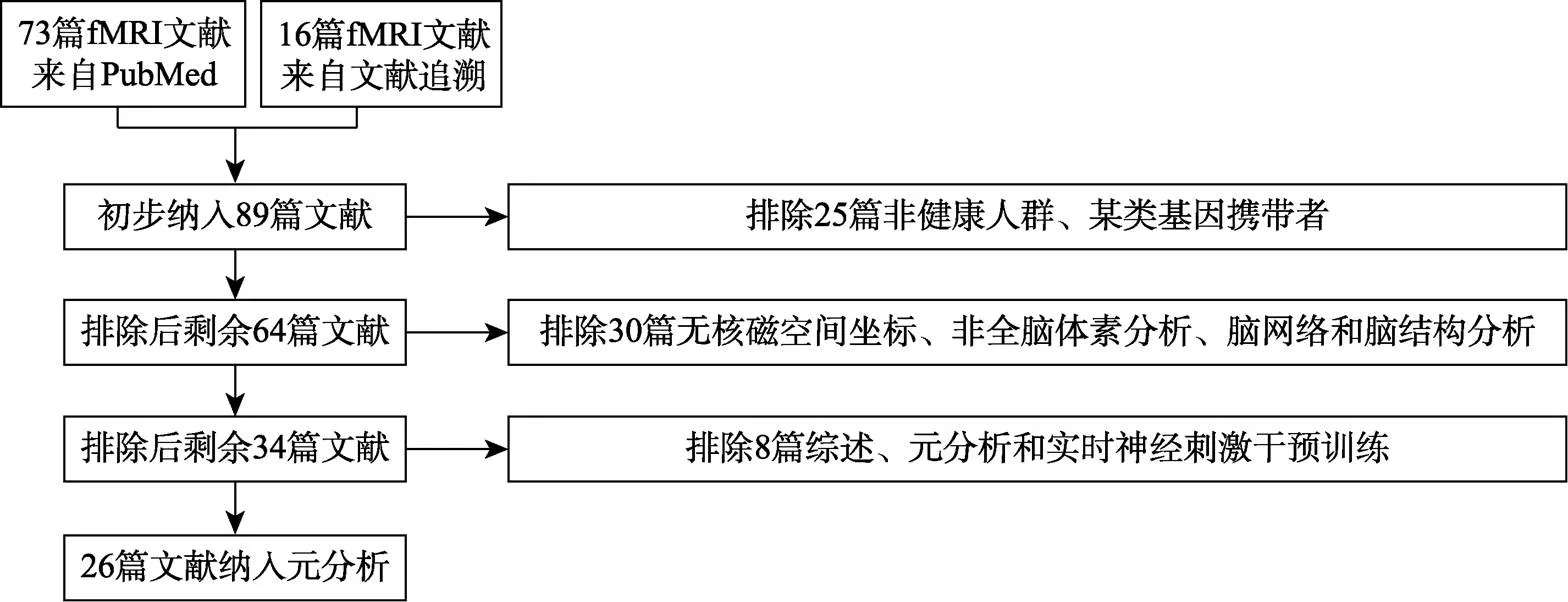
图2 逐步排除和纳入元分析的标准

表5 纳入元分析文献的基本信息(共26篇文献, 31个实验)
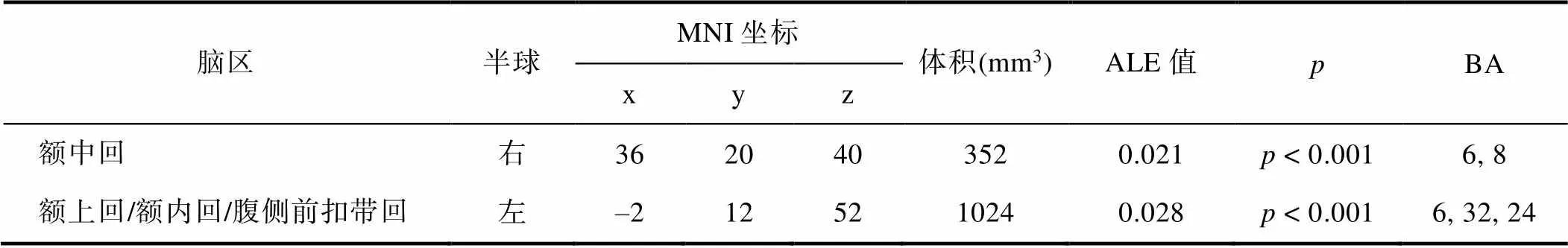
表6 WMT减弱脑区激活的元分析结果(共26篇文献, 31个实验)
注:(1)坐标为激活簇中心脑坐标; (2)ALE为在该激活簇内最大的激活似然值,值为训练诱发该脑区激活减弱的零假设概率; (3)BA为Brodmann分区。BA6既包括额中回, 也包括额上回; BA8为额中回; BA32为背侧前扣带回, 属于内侧额叶(额内回); BA24为腹侧前扣带回。
4 WMT脑区分布递减时空模型的结果和讨论
综合以上定性和定量研究结果, 主要对频数分析和元分析结果做建模, 围绕脑区的空间和时间变化, 提出WMT脑区分布递减时空模型(见图4)。据此得到5个研究结果和对应的讨论。其中, 前三个结果由描述统计得到; 第四和五个结果由推断统计得到, 它们与描述统计结果呼应, 解释了WMT敏感的大脑子区及其变化。
第一, WMT效应主要体现在脑区神经活动的减弱或增强上, 且减弱效应更突出。首先, 脑区神经活动减弱可以用“神经效率假说”解释, 被认为是神经加工或神经回路效率提高的表现(Brooks et al., 2020; Constantinidis & Klingberg, 2016)。持续训练可能促使被训练者的信息加工方式由控制加工过渡到较大程度的自动加工, 从而降低对注意控制的需求, 优化认知资源, 产生更有效的一般性神经加工(Schneiders et al., 2011)。其次, 脑区激活增强反映了参与信息加工的神经组织增多、神经元活性增强(Buschkuehl et al., 2012), 或在某个脑区诱发了更强烈的脑响应(Kelly et al., 2006)。因为WMT引起树突、轴突等细胞成分形成的增加, 这可能构成了皮层活动增强的神经基础, 从而诱发神经元反应增强和神经元数量增加, 并引起局部BOLD信号增强。另外, 减弱和增强现象并存, 可能由于在不同的WMT阶段, 被训练者接受不同的训练强度或训练内容, 由此采用不同的认知策略来解决问题, 从而诱发不同的神经机制。未来需设计更精细的实验, 来辨析这种神经机制差异。

图3 WMT减弱脑区激活的2个激活簇区域(共26篇文献, 31个实验)
第二, WMT效应体现在额顶叶联合脑区、边缘系统和基底神经核等多个联合脑区上, 且额顶叶联合区尤为突出。
以往神经成像研究也揭示, WM的神经基质分布在复合的皮层区域, 尤其在前额叶、后顶叶和前扣带皮层(Alagapan et al., 2019)。其中, 额顶叶网络及其子区域可以依据任务目标来改变它与其它网络节点的FC, 其连接模式既可以反映具体任务的参与, 也能促进新任务的学习(Zanto & Gazzaley, 2013)。额顶叶网络参与大量认知过程, 比如WM、流体智力(Assem et al., 2020)、情景记忆和来源记忆(Eschmann et al., 2020)、视空间注意(Lobier et al., 2018)、珠心算(Li, Hu, et al., 2013)和外显情绪调节(Pozzi et al., 2020)等, 这间接支持了额顶叶联合脑区是WMT效应的主要脑区, 其激活减弱反映了自动化加工的增多。尤其, 顶下小叶(inferior parietal lobule)较明显地体现WMT效应。其参与WM网络的注意控制过程, 是更新训练范式激活脑区中常见的部位(Dahlin et al., 2008; Schneiders et al., 2011)。同时, 顶下小叶可能属于抑制网络的重要脑区, 抑制活动会增强该区激活, 反映了抑制过程中信息维持能力的增强(Kelly et al., 2006)。人脑的功能作用, 不是由相对分开的区域所完成。故整个皮层结构是如何协同整合WMT效应的, 这需要进一步探索。
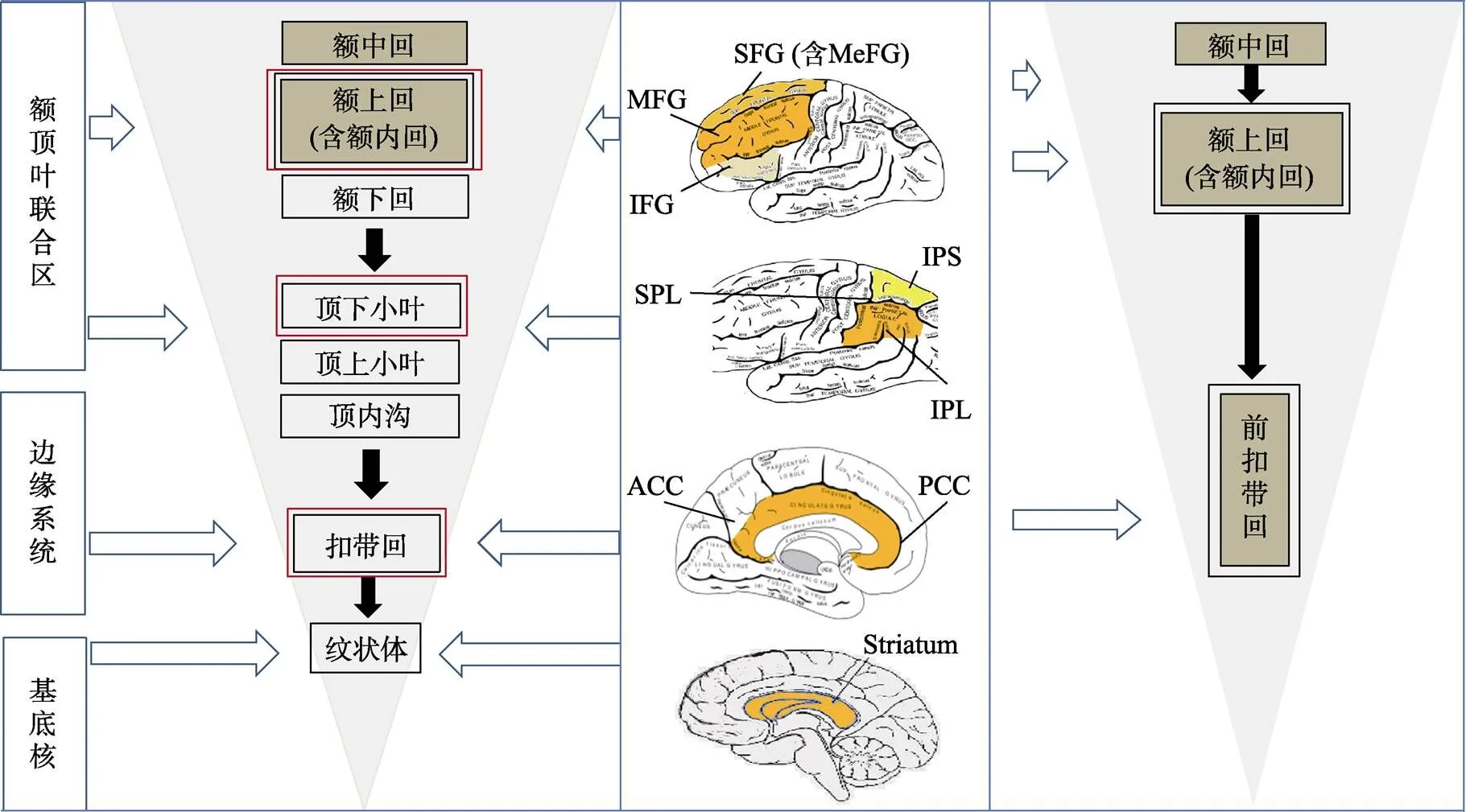
图4 WMT脑区分布递减时空模型(倒金字塔图)
注:左侧倒三角为频数分析结果(37篇文献, 42个实验), 右侧倒三角为元分析结果(26篇文献, 31个实验); 棕色框内的脑区为元分析差异具有统计学意义; 双黑线框内的脑区为只出现激活减弱, 单黑线框内的脑区为激活减弱或增强并存; 红线框内的脑区为激活减弱明显多于激活增强。SFG = Superior Frontal Gyrus (额上回), MeFG = Medial Frontal Gyrus (额内回), MFG = Middle Frontal Gyrus (额中回), IFG = Inferior Frontal Gyrus (额下回), SPL = Superior Parietal Lobule (顶上小叶), IPL = Inferior Parietal Lobule (顶下小叶), IPS = Intraparietal Sulcus (顶内沟), ACC = Anterior Cingulate Gyrus (前扣带回), PCC = Posterior Cingulate Gyrus (后扣带回)。图中间位置的4个脑区模型图片来自:A+医学百科网(www.a-hospital.com), 作者在其上新增了色彩和英文缩写。
第三, 纹状体(Striatum)明显体现了WMT效应。纹状体激活减弱也符合神经效率假说, 反映了更新的自动化加工增强(Kühn et al., 2013)和跨模态(一般控制)的神经加工效率提高(Brooks et al., 2020); 但纹状体激活增强可能反映的也是WMT提高了WM信息的维持和更新效率(Dahlin et al., 2008; Kühn et al., 2013; Salminen et al., 2016); 而更新训练可促进纹状体内多巴胺的释放, 增强纹状体激活(Bäckman et al., 2017)。而纹状体激活和多巴胺释放如何改善认知行为, 仍需进一步研究。
第四, 额中回和额上回体现的WMT效应具有统计学意义。这两个脑回一直被认为是参与WM等多种心理活动的重要脑部位, 它们与WM激活的顶内沟(即背侧顶叶皮层)存在相关(Harms et al., 2013), 但额中回参与WM的证据更为广泛(Alagapan et al., 2019)。
额中回的激活强度和皮层厚度分别与WM成绩呈正相关和负相关(Owens et al., 2018), 它通过参与WM中的注意活动来影响WM成绩。额中回参与多种注意加工, 其激活强度与注意信息调节活动密切相关(Zhu et al., 2018; Japee et al., 2015), 在调节注意网络(Song et al., 2019)、维持注意网络完整性(Gogulski et al., 2017)、调控持续性注意(Han et al., 2018; Song et al., 2019; Neale et al., 2015)和警觉(Neale et al., 2015)方面也发挥重要作用。而且, 右侧额中回被认为是背侧和腹侧注意网络(the dorsal and ventral attention networks)的交汇位置, 它通过发挥“断路器” (circuit-breaker)的功能来中断内源性注意和重新定位外源性注意, 在调节外源性注意转向内源性注意的活动中扮演重要角色(Japee et al., 2015)。对变化事件的预测活动会在对视觉信息进行储存、维持和更新的三个WM环节中全部选择性地激活额中回(Heinzel et al., 2016)。总之, 额中回激活减弱体现了对注意控制加工需求的降低和一般控制加工的灵活转换; 而少数情况下额中回激活增强, 则反映了任务相关的神经元活动增强或参与加工的树突、轴突等神经结构数量增多(Westerberg & Klingberg, 2007)。而额中回与记忆的直接关系, 仍需进一步探索。
额上回分三个亚区:前内侧(SFGam)、背外侧(SFGdl)和后内侧(SFGp)。静息态FC分析表明, SFGam主要与认知控制网络和DMN(前扣带皮层和中扣带皮层)相关; SFGdl与认知执行网络和DMN(额中回)相关; SFGp与感觉运动相关脑区(中央前回、尾状核、丘脑和额叶岛盖)相关(Li, Qin, et al., 2013)。可见, 额上回是WM脑网络中的一个重要节点(Alagapan et al., 2019), 参与WM的多种活动(Briggs et al., 2020)。额上回的BA8区与空间WM的客体信息维持活动相关更为密切(Briggs et al., 2020); 另外, 当左侧额上回在theta和alpha频段表现出的内源性振荡与WM刺激频率相匹配(尤其在编码阶段)时, 就会缩短被训练者执行WM任务的反应时, 这支持了该脑区对WM的调节作用(Alagapan et al., 2019)。由此, 额上回激活减弱体现了自上而下注意控制的减弱(Milham et al., 2003)和WM各类加工过程(如更新、集中注意、刺激选择和排列)效率的提升(Miró-Padilla et al., 2019)。额上回的功能具有复杂性。那么, 在亚区水平上(subregional level)辨析其在WMT中的不同作用, 可能是未来的研究方向之一。
第五, 前扣带回体现了显著的WMT效应。前扣带回与前额叶和后顶叶被合称为认知活动的脚手架(scaffolding) (Schneiders et al., 2011), 它着重参与言语WM (Emch et al., 2019)、注意(Miró- Padilla et al., 2019)、冲突加工(Kanske & Kotz, 2011)、监控竞争、动机、情绪冲突、错误检测和任务转换(Zhu et al., 2018)等。可见, 前扣带回具有复杂认知功能, 其激活减弱反映了WM能力的提升。但是, 前扣带回如何参与WMT中错误检测?而后扣带回如何通过预期或动机来间接参与WMT?可见, 扣带回的不同结构在WMT如何发挥不同作用, 其机制需要进一步探究。
5 WMT效应的调节因素
模型显示, WMT对脑区神经活动的改变具有多样性, 尤其表现出减弱和增强在不同实验中并存。那么, 对立现象并存的原因可能是什么?这需要追溯WMT效应可能的调节因素。
对其中33个实验(来自31篇文献)的调节因素做频数分析后(见图5)发现:第一, 训练人数变化较大, 跨度范围在3~41人, 以10~20人(15.2 ± 3.0)居多; 第二, 有2种神经活动分析指标, 以BOLD (21个)为主、FC (12个)为辅; 第三, 被训练者的年龄跨度范围较大, 以青年(24个)为主, 也包含少数儿童(3个)和中老年(6个); 第四, 训练任务具有多样性, 更新(17个)较多, 也有短时记忆(9个)、抑制(3个)、转换(1个)、计算(1个)和多种综合心理(2个)等; 第五, 训练时间具有多样性, 以历时4周以下居多(19个), 也包括4~6周(11个)和6周以上(3个), 但跨度范围很大(20分钟~5年)。
然后, 用非参数检验检验这33个实验中的调节因素是否影响WMT诱发的神经活动减弱或增强, 结果(见表7)显示:第一, 不同训练任务类型诱发的减弱与增强比率差异具有统计学意义(χ2= 10.56,= 5,= 0.023)。其中, 更新训练诱发减弱(14个)显著多于其增强(3个); 第二, 不同训练时长的减弱与增强比率差异具有统计学意义(χ2= 7.57,= 2,= 0.019)。其中, 训练周期为4周以下的减弱(15个)显著多于增强(4个)。可见, 更新任务和较短时间的WMT倾向于减弱脑区神经活动。
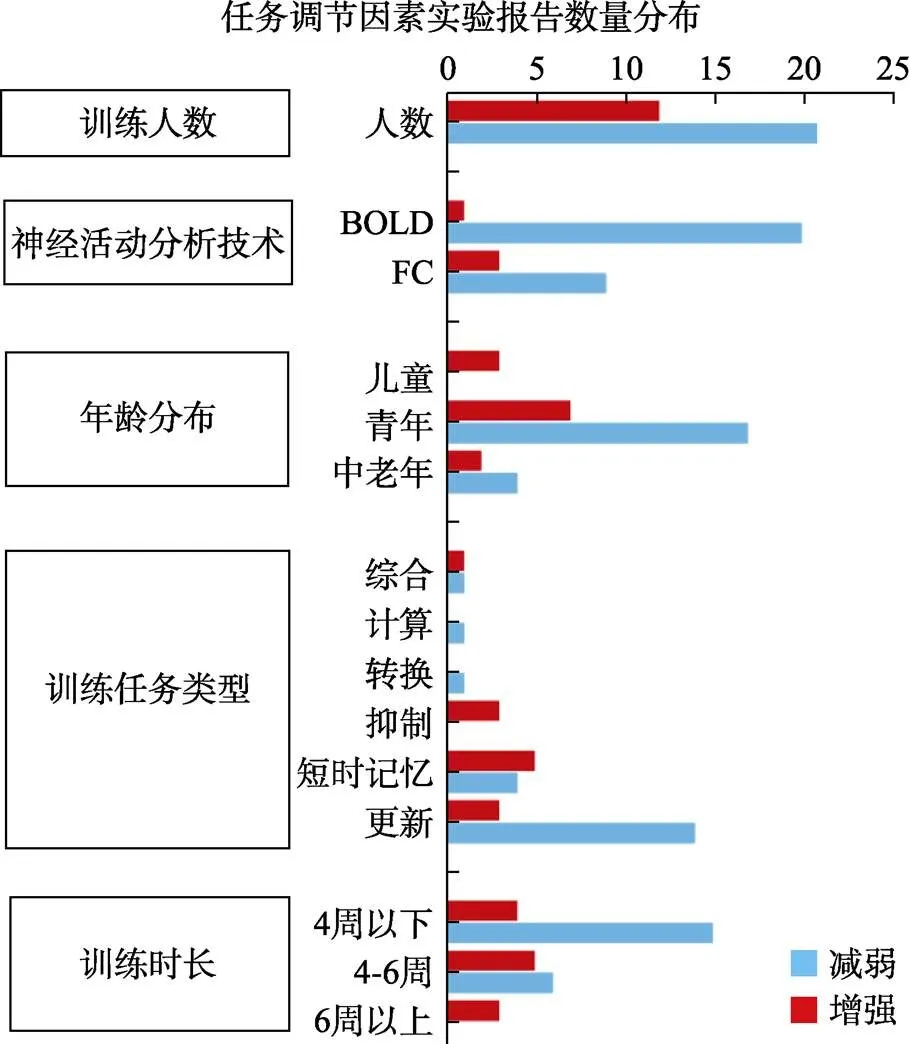
图5 WMT效应调节因素的神经活动减弱与增强频数分布(31篇文献。33个实验)
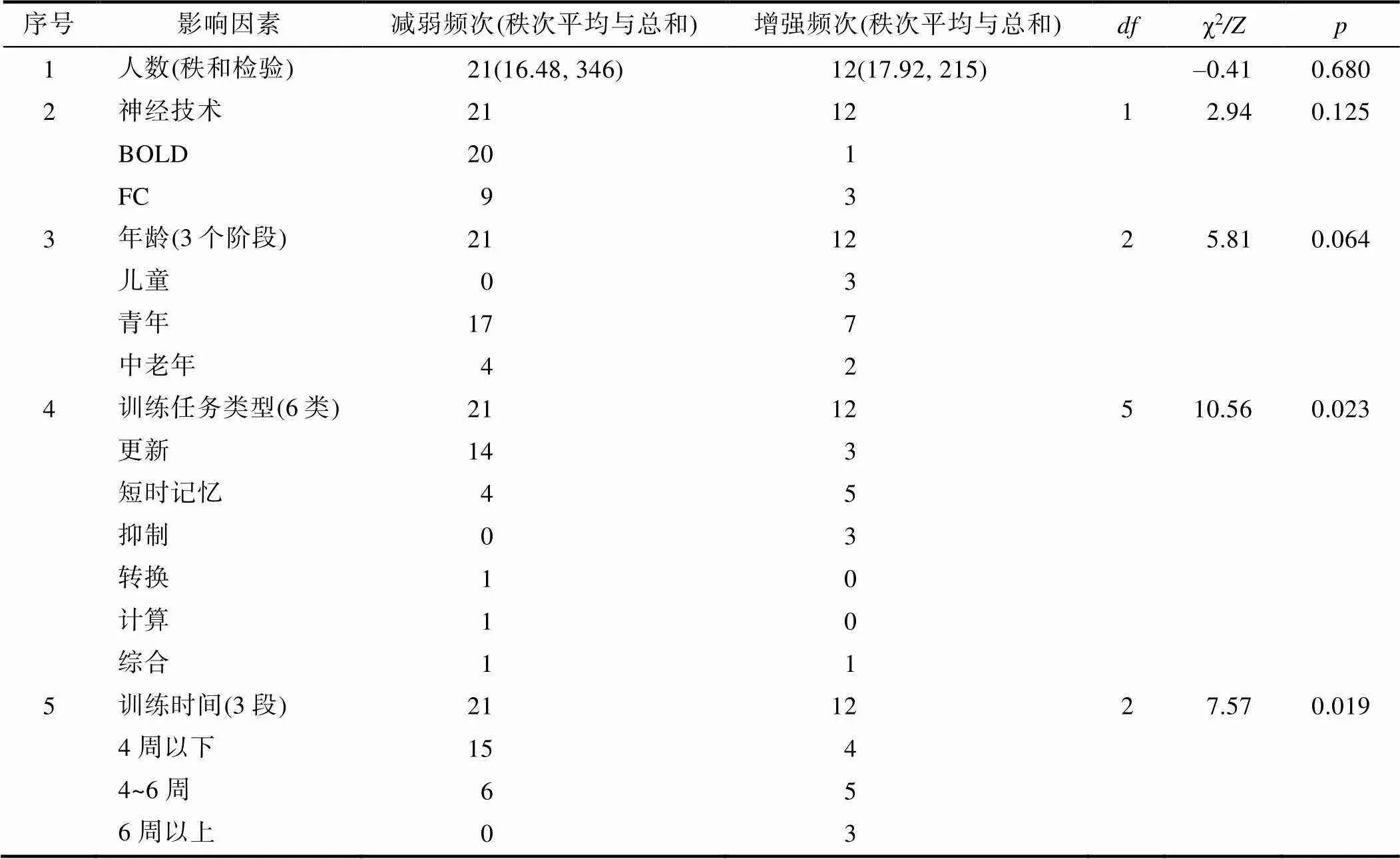
表7 WMT效应的调节因素影响神经活动改变的非参数检验(31篇文献, 33个实验)
注:第1项(人数)为定距变量, 且每一类样本容量小于30, 但至少有一类样本容量大于20, 故采用非参数检验-Mann- Whitney U Test中的Z值; 第2~5项为定类和定序变量, 且存在任一理论频数小于5, 故采用卡方检验的Fisher校正, [参考:邵志芳. (2017).(第三版, pp.349−350, 333). 中国轻工业出版社.]。
6 结论和展望
学习诱发的可塑性(learning-induced plasticity)在WMT中尤为明显。为探明WMT改变脑区神经活动的时空特性, 以ExtPFIT和“神经效率假说”为依据, 采用逐层递进方法, 查究近20年来正常人群WMT的37篇fMRI文献, 从脑区激活模式和脑网络FC上比较各个具体脑区在WMT前后发生的变化, 据此建立WMT脑区分布递减时空模型。最后, 综合定性和定量分析结果, 针对正常人群WMT诱发神经可塑性的时空特性, 得出3个结论:第一, WMT减弱或增强了正常人群脑区的神经活动, 但减弱活动更加突出, 且更新和较短时间的训练倾向于诱发较多激活减弱; 第二, 这些神经活动变化主要发生在额顶叶联合区, 但也包括颞叶、枕叶、扣带回及纹状体等联合脑区, 在一定范围内体现了整脑功能联合。这体现了WMT诱发神经可塑性的空间特性, 且符合“扩展的智力顶额整合理论”; 第三, 额中回、额上回顶下小叶和扣带回(尤其前扣带回)这4个子区减弱的神经活动重点展示了WMT诱发神经可塑性的时间特性, 且符合“神经效率假说”, 恰好体现出“聪明的大脑更懒惰”。
可见, WMT如何通过减弱或增强脑区神经活动来提升WM能力及相关认知功能, 是一个兼具研究价值与挑战性的问题。未来需进一步探究WMT诱发与学习有关的大规模通用神经网络机制(Salmi et al., 2018), 寻找WMT神经机制更具普适性的规律。尤其, 训练诱发的减弱和增强并存现象, 促使人思考:思维靠脑细胞“加班”, 还是靠“效率高”?Genc团队(2018)在《自然》子刊上报道, 用轴突定向分散和密度成像(Neurite Orientation Dispersion and DensityImaging, NODDI)这种新的弥散MRI技术, 准确检测到了大脑神经元树突分布密度, 发现在做智力测试时, 普通智商人的神经元之间“密集沟通”; 而高智商者的每个神经元表现出超强的目的性, 他们伸出寥寥无几的树突就能准确找到另一个神经元伙伴, 毫不费力就完成了任务, 真叫一个轻松惬意。故WMT诱发神经可塑性的未来研究可能关注脑可塑性中的低活跃性, 辨析额中回、额上回、顶下小叶和扣带回(尤其前扣带回)这4个子区在激活减弱水平上体现的时间特性, 找寻训练减弱或增强大脑活动的综合性影响因素。
*代表元分析用到的文献
宋杰, 赵琬, 张秋梅, 李君. (2020).大学生工作记忆广度训练迁移效应的行为学与影像学研究.,(10), 909–914.
Alagapan, S., Lustenberger, C., Hadar, E., Shin, H. W., & Frӧhlich, F. (2019). Low-frequency direct cortical stimulation of left superior frontal gyrus enhances working memory performance.,, 697–706.
Assem, M., Blank, I. A., Mineroff, Z., Ademoğlu, A., & Fedorenko, E. (2020). Activity in the fronto-parietal multiple-demand network is robustly associated with individual differences in working memory and fluid intelligence.,, 1–16.
Bäckman, L., Waris, O., Johansson, J., Andersson, M., Rinne, J. O., Alakurtti, K., … Nyberg, L. (2017). Increased dopamine release after working-memory updating training: Neurochemical correlates of transfer.(1), 7160.
*Beatty, E. L., Jobidon, M.-E., Bouak, F., Nakashima, A., Smith, I., Lam, Q., … Vartanian, O. (2015). Transfer of training from one working memory task to another: Behavioural and neural evidence.,, 86.
Briggs, R. G., Khan, A. B., Chakraborty, A. R., Abraham, C. J., Anderson, C. D., Karas, P. J., … Sughrue, M. E. (2020). Anatomy and white matter connections of the superior frontal gyrus.,(6), 823–832.
Brooks, S. J., Mackenzie-Phelan, R., Tully, J., & Schiöth, H. B. (2020). Review of the neural processes of working memory training: Controlling the impulse to throw the baby out with the bathwater.,, 512761.
Buschkuehl, M., Jaeggi, S. M., & Jonides, J. (2012). Neuronal effects following working memory training.,, 167–179.
*Clark, C. M., Lawlor-Savage, L., & Goghari, V. M. (2017). Functional brain activation associated with working memory training and transfer.,, 34–49.
Constantinidis, C., & Klingberg, T. (2016). The neuroscience of working memory capacity and training.,(7), 438–449.
*Dahlin, E., Neely, A. S., Larsson, A., Backman, L., & Nyberg, L. (2008). Transfer of learning after updating training mediated by the striatum.,(5882), 1510–1512.
Dörrenbächer, S., Wu, C., Zimmer, H., & Kray, J. (2020). Plasticity in brain activity dynamics after task-shifting training in older adults.,, 107285.
Eickhoff, S. B., Laird, A. R., Grefkes, C., Wang, L. E., Zilles, K., & Fox, P. T. (2009). Coordinate-based activation likelihood estimation meta-analysis of neuroimaging data: A random-effects approach based on empirical estimates of spatial uncertainty.(9), 2907–2926.
*Emch, M., Ripp, I., Wu, Q., Yakushev, I., & Koch, K. (2019). Neural and behavioral effects of an adaptive online verbal working memory training in healthy middle-aged adults.,, 300.
Eschmann, K. C. J., Bader, R., & Mecklinger, A. (2020). Improving episodic memory: Frontal-midline theta neurofeedback training increases source memory performance.,, 117219.
*Flegal, K. E., Ragland, J. D., & Ranganath, C. (2019). Adaptive task difficulty influences neural plasticity and transfer of training.,, 111–121.
*Garavan, H., Kelley, D., Rosen, A., Rao, S. M., & Stein, E. A. (2000). Practice-related functional activation changes in a working memory task.,(1), 54–63.
Genç, E., Fraenz, C., Schlüter, C., Friedrich, P., Hossiep, R., Voelkle, M. C., … Jung, R. E. (2018). Diffusion markers of dendritic density and arborization in gray matter predict differences in intelligence.(1):1905.
Gogulski, J., Zetter, R., Nyrhinen, M., Pertovaara, A., & Carlson, S. (2017). Neural substrate for metacognitive accuracy of tactile working memory.5343–5352.
Gur, R. C., Butler, E. R., Moore, T. M., Rosen, A. F. G., Ruparel, K., Satterthwaite, T. D., … Gur, R. E. (2020). Structural and functional brain parameters related to cognitive performance across development: Replication and extension of the Parieto-Frontal Integration Theory in a single sample.,(3), 1444–1463.
Han, Q., Zhang, Y., Liu, D. H., Wang, Y., Feng, Y. J., Yin, X. T., & Wang, J. (2018). Disrupted local neural activity and functional connectivity in subjective tinnitus patients: Evidence from resting-state fMRI study.,(11), 1193–1201.
Harms, M. P., Wang, L., Csernansky, J. G., & Barch, D. M. (2013). Structure-function relationship of working memory activity with hippocampal and prefrontal cortex volumes.,(1), 173–186.
Heinzel, S., Lorenz, R. C., Brockhaus, W.-R., Wüstenberg, T., Kathmann, N., Heinz, A., & Rapp, M. A. (2014). Working memory load-dependent brain response predicts behavioral training gains in older adults.,(4), 1224–1233.
*Heinzel, S., Lorenz, R. C., Pelz, P., Heinz, A., Walter, H., Kathmann, N., … Stelzel, C. (2016). Neural correlates of training and transfer effects in working memory in older adults.,, 236–249.
Hempel, A., Giesel, F. L., Garcia Caraballo, N. M., Amann, M., Meyer, H., Wüstenberg, T., … Schröder, J. (2004). Plasticity of cortical activation related to working memory during training.,(4), 745–747.
Iordan, A. D., Cooke, K. A., Moored, K. D., Katz, B., Buschkuehl, M., Jaeggi, S. M., … Reuter-Lorenz, P. A. (2020). Neural correlates of working memory training: Evidence for plasticity in older adults.,, 116887.
*Jansma, J. M., Ramsey, N. F., Slagter, H. A., & Kahn, R. S. (2001). Functional anatomical correlates of controlled and automatic processing.,(6), 730–743.
Japee, S., Holiday, K., Satyshur, M. D., Mukai, I., & Ungerleider, L. G. (2015). A role of right middle frontal gyrus in reorienting of attention: A case study.,, 23.
*Jolles, D. D., Grol, M. J., van Buchem, M. A., Rombouts, S. A. R. B., & Crone, E. A. (2010). Practice effects in the brain: Changes in cerebral activation after working memory practice depend on task demands.,(2), 658–668.
Jolles, D. D., van Buchem, M. A., Crone, E. A., & Rombouts, S. A. R. B. (2013). Functional brain connectivity at rest changes after working memory training.,(2), 396–406.
Jung, R. E., & Haier, R. J. (2007). The Parieto-Frontal Integration Theory (P-FIT) of intelligence: Converging neuroimaging evidence.,(2), 135–187.
Kanske, P., & Kotz, S. A. (2011). Emotion triggers executive attention: Anterior cingulate cortex and amygdala responses to emotional words in a conflict task.,(2), 198–208.
*Kelly, A. M. C., Hester, R., Foxe, J. J., Shpaner, M., & Garavan, H. (2006). Flexible cognitive control: Effects of individual differences and brief practice on a complex cognitive task.,(2), 866–886.
*Kühn, S., Schmiedek, F., Noack, H., Wenger, E., Bodammer, N. C., Lindenberger, U., & Lövden, M. (2013). The dynamics of change in striatal activity following updating training.,(7), 1530–1541.
Lancaster, J. L., Tordesillas-Gutiérrez, D., Martinez, M., Salinas, F., Evans, A., Zilles, K., … Fox, P. T. (2007). Bias between MNI and Talairach coordinates analyzed using the ICBM-152 brain template.(11), 1194–1205.
Landau, S. M., Garavan, H., Schumacher, E. H., & D’Esposito, M. (2007). Regional specificity and practice: Dynamic changes in object and spatial working memory.,, 78–89.
*Landau, S. M., Schumacher, E. H., Garavan, H., Druzgal, T. J., & D’Esposito, M. (2004). A functional MRI study of the influence of practice on component processes of working memory.,(1), 211–221.
Li, W., Qin, W., Liu, H. G., Fan, L. Z., Wang, J. J., Jiang, T. Z., & Yu, C. S. (2013). Subregions of the human superior frontal gyrus and their connections.,46–58.
*Li, Y. X., Hu, Y. Z., Zhao, M., Wang, Y. Q., Huang, J., & Chen, F. Y. (2013). The neural pathway underlying a numerical working memory task in abacus-trained children and associated functional connectivity in the resting brain.,, 24–33.
Lobier, M., Palva, J. M., & Palva, S. (2018). High-alpha band synchronization across frontal, parietal and visual cortex mediates behavioral and neuronal effects of visuospatial attention.,, 222–237.
*Milham, M. P., Banich, M. T., Claus, E. D., & Cohen, N. J. (2003). Practice-related effects demonstrate complementary roles of anterior cingulate and prefrontal cortices in attentional control.,(2), 483–493.
*Miró-Padilla, A., Bueichekú, E., & Ávila, C. (2020). Locating neural transfer effects of n-back training on the central executive: A longitudinal fMRI study.,(1), 5226.
*Miró-Padilla, A., Bueichekú, E., Ventura-Campos, N., Flores-Compañ, M.-J., Parcet, M. A., & Ávila, C. (2019). Long-term brain effects of N-back training: An fMRI study.,(4), 1115–1127.
Neale, C., Johnston, P., Hughes, M., & Scholey, A. (2015). Functional activation during the rapid visual information processing task in a middle aged cohort: An fMRI study.,(10), e0138994.
*Nęcka, E., Gruszka, A., Hampshire, A., Sarzyńska-Wawer, J., Anicai, A.-E., … Soreq, E. (2021). The effects of working memory training on brain activity.(2), 155.
*Olesen, P. J., Westerberg, H., & Klingberg, T. (2004). Increased prefrontal and parietal activity after training of working memory.,(1), 75–79.
Owens, M. M., Duda, B., Sweet, L. H., & MacKillop, J. (2018). Distinct functional and structural neural underpinnings of working memory.,, 463–471.
Pozzi, E., Vijayakumar, N., Rakesh, D., & Whittle, S. (2020). Neural correlates of emotion regulation in adolescents and emerging adults: A meta-analytic study.(2), 194–204.
Salmi, J., Nyberg, L., & Laine, M. (2018). Working memory training mostly engages general-purpose large-scale networks for learning.,, 108–122.
Salmi, J., Vilà-Balló, A., Soveri, A., Rostan, C., Rodríguez-Fornells, A., Lehtonen, M., & Laine, M. (2019). Working memory updating training modulates a cascade of event-related potentials depending on task load.,107085.
*Salminen, T., Kühn, S., Frensch, P. A., & Schubert, T. (2016). Transfer after dual n-back training depends on striatal activation change.,(39), 10198–10213.
Sánchez-Pérez, N., Inuggi, A., Castillo, A., Campoy, G., García-Santos, J. M., González-Salinas, C., & Fuentes, L. J. (2019). Computer-based cognitive training improves brain functional connectivity in the attentional networks: A study with primary school-aged children.,, 1–12.
*Sayala, S., Sala, J. B., & Courtney, S. M. (2006). Increased neural efficiency with repeated performance of a working memory task is information-type dependent.,(5), 609–617.
*Schneiders, J. A., Opitz, B., Krick, C. M., & Mecklinger, A. (2011). Separating intra-modal and across-modal training effects in visual working memory: An fMRI investigation.,(11), 2555–2564.
*Schneiders, J. A., Opitz, B., Tang, H., Deng, Y., Xie, C. X., Li, H., & Mecklinger, A. (2012). The impact of auditory working memory training on the fronto-parietal working memory network.,, 173.
*Schweizer, S., Grahn, J., Hampshire, A., Mobbs, D., & Dalgleish, T. (2013). Training the emotional brain: Improving affective control through emotional working memory training.,(12), 5301–5311.
Song, P., Lin, H., Liu, C., Jiang, Y., Lin, Y., Xue, Q., … Wang, Y. (2019). Transcranial magnetic stimulation to the middle frontal gyrus during attention modes induced dynamic module reconfiguration in brain networks.1–9.
Takeuchi, H., Taki, Y., Nouchi, R., Hashizume, H., Sekiguchi, A., Kotozaki, Y., … Kawashima, R. (2013). Effects of working memory training on functional connectivity and cerebral blood flow during rest.,(8), 2106–2125.
*Takeuchi, H., Taki, Y., Nouchi, R., Hashizume, H., Sekiguchi, A., Kotozaki, Y., … Kawashima, R. (2014). Working memory training improves emotional states of healthy individuals.200.
Thompson, T. W., Waskom, M. L., & Gabrieli, J. D. E. (2016). Intensive working memory training produces functional changes in large-scale frontoparietal networks.,(4), 575–588.
Turkeltaub, P. E., Eickhoff, S. B., Laird, A. R., Fox, M., Wiener, M., & Fox, P. (2012). Minimizing within-experimentand within-group effects in Activation Likelihood Estimation meta-analyses.(1), 1–13.
*Wang, C., Xu, T. Y., Geng, F. J., Hu, Y. Z., Wang, Y. Q., Liu, H. F., & Chen, F. Y. (2019). Training on abacus-based mental calculation enhances visuospatial working memory in children.,(33), 6439– 6448.
*Westerberg, H., & Klingberg, T. (2007). Changes in cortical activity after training of working memory—A single-subject analysis.,(1–2), 186–192.
Zanto, T. P., & Gazzaley, A. (2013). Fronto-parietal network: Flexible hub of cognitive control.,(12), 602–603.
Zhou, H., Geng, F. J., Wang, Y. Q., Wang, C. J., Hu, Y. Z., & Chen, F. Y. (2019). Transfer effects of abacus training on transient and sustained brain activation in the frontal– parietal network.,, 135–146.
Zhu, J., Li, J. X., Li, X. W., Rao, J., Hao, Y. R., Ding, Z. J., & Wang, G. P. (2018). Neural basis of the emotional conflict processing in major depression: ERPs and source localization analysis on the N450 and P300 components.,, 214.
附录:
文献中全部脑区做频数分析的完整版本见如下附表1。因为篇幅较长, 故在正文中对其作压缩删减后成为表3。

Neuroplasticity induced by working memory training: A spatio-temporal model of decreased distribution in brain regions based on fMRI experiments
CHEN Xingming1, FU Tong1, LIU Chang3, ZHANG Bin1, FU Yunfa4, LI Enze5, ZHANG Jian6, CHEN Shengqiang2, DANG Caiping1,2
(1Brain Hospital Affiliated to Guangzhou Medical University, Guangzhou 510370, China) (2School of Health Management, Guangzhou Medical University, Guangzhou 511436, China) (3School of Psychology, Nanjing Normal University, Nanjing 210024, China) (4School of Information Engineering and Automation, School of Medicine, Kunming University of Science and Technology, Kunming 650031, China) (5Department of Psychology, Nanfang Hospital Affiliated to Southern Medical University, Guangzhou 510515, China) (6Tianjing Medical University,Tianjing 300070, China)
Working memory training (WMT) induces neuroplasticity, but its specific mechanism remains unclear. In order to explore the spatio-temporal characteristics of brain function changes induced by WMT, the findings of 37 fMRI papers on WMT in normal population over the last 20 years were investigated with a step-by-step approach, including 5 methods and the following 6 steps, as well as with two assumptions based on the two theories— “Extended Parieto-Frontal Integration Theory” (ExtPFIT) and “neuro efficiency hypothesis”. Step 1, the changes of activation pattern and function connectivity in various brain regions before and after WMT among the 37 papers were compared using descriptive review, frequency analysis and Chi-square test. It was found that 5 association areas, 7 macro regions and 3 sub-regions of the brain were changed by WMT. Especially, the number of articles reporting decreased activation in 3 sub-regions— superior frontal gyrus, inferior parietal lobule and cingulate gyrus was more than that reporting increased activation in them, and this difference was statistically significant. Step 2, 26 out of the 37 papers were used to conduct meta-analysis with activation likelihood estimation method, and it showed that 3 sub-regions with the statistically significant effects in their decreased activation were middle frontal gyrus (BA6 and 8), superior frontal gyrus (BA6) and anterior cingulate gyrus (BA24 and 32). Step 3, based on the comprehensive results of qualitative and quantitative research analysises, a spatio-temporal model of decreased distribution in brain regions was proposed, and there were 5 results and their discussions obtained with the said model. Step 4, 33 out of the 37 papers were analyzed using non-parametric tests to check the factors that possibly regulated the training effect of WMT, which showed that the task type and time of the training had statistically significant effects on brain activation. Step 5, according to the above analysises, 3 conclusions were drawn regarding the neuroplasticity induced by WMT in normal population, i.e. 1) WMT can either weaken or enhance the brain activities, and the weakening effect appears more prominent. In addition, updating and shorter time of WMT tend to induce more weakening effects; 2) Such changes occur mostly in the frontal-parietal association area, but also possibly in the temporal lobe, occipital lobe, cingulate gyrus, striatum and other association areas, which reflects the whole brain functional association within a certain range. It highlights the spatial characteristics of neuroplasticity induced by WMT, which fits in easily with ExtPFIT; 3) The 4 sub-regions of the middle frontal gyrus, superior frontal gyrus, inferior parietal lobule and cingulate gyrus (anterior in particular) in decreased activation mainly reflect the temporal characteristics of neuroplasticity induced by WMT, which fits in easily with “neural efficiency hypothesis” and “smarter brains may be slightly lazier”. Step 6, it was proposed that further research of neuroplasticity induced by WMT was to possibly focus on low brain activity on its plasticity, distinguish the temporal characteristics of the training effect reflected in 4 sub-regions of middle frontal gyrus, superior frontal gyrus, inferior parietal lobule and cingulate gyrus (anterior in particular) in decreased activation as well as to probe the comprehensive influencing factors that increase or decrease the brain activities.
working memory training, neuroplasticity, a spatio-temporal model of decreased distribution in brain regions, middle frontal gyrus, superior frontal gyrus, anterior cingulate gyrus
B842
2021-02-17
* 国家自然科学基金资助项目(31400874), 广州市哲学社会科学规划课题(15G99), 广州医科大学高水平大学建设项目(02-410-B205001186)。
#为并列第一作者。
党彩萍, E-mail: dcp619@163.com
