SbGA20ox1•SbGA20ox3和SbGA2ox3基因对高粱品种赤霉素表达水平的影响
2022-02-19商靖陆劲羽逄洪波李雪梅王艳秋李玥莹
商靖 陆劲羽 逄洪波 李雪梅 王艳秋 李玥莹



摘要 [目的]研究SbGA20ox1、SbGA20ox3和SbGA2ox3基因对高粱品种赤霉素表达水平的影響。[方法]以不同高粱品种为试材(9544.7057为父本与不同母本进行杂交得到子代:辽夏梁1号、辽2297、辽2697、辽5397、01-26B;以及01-26A为母本与不同父本进行杂交得到子代:辽杂35、0-01、9544.7057、3550、辽2697),利用实时荧光定量PCR技术比较不同时期(拔节期、抽穗期、成熟期),不同品种的高粱之间SbGA20ox1基因、SbGA20ox3基因、SbGA2ox3基因表达含量规律。[结果]SbGA20ox1基因在同一父本不同母本高粱组的基因表达规律基本符合抽穗期>成熟期>拔节期;SbGA2ox3基因在同一父本不同母本高粱组的基因表达规律基本符合抽穗期>成熟期>拔节期,SbGA2ox3基因在同一母本不同父本高粱组的基因表达规律基本符合拔节期>成熟期>抽穗期。而SbGA20ox1基因在同一母本不同父本高粱组的基因表达以及SbGA20ox3基因在各品种高粱的基因表达并未发现明显规律,各时期基因的相对表达量大部分品种间无明显差异。[结论]该研究一定程度上揭示了SbGA20ox1、SbGA20ox3和SbGA2ox3基因在多个高粱品种各时期的基因表达规律,为进一步研究调控赤霉素水平的小基因家族的作用机制提供启示。
关键词 高粱;相对表达量;实时荧光定量;赤霉素;小基因家族
中图分类号 S 188文献标识码 A文章编号 0517-6611(2022)02-0105-04
doi:10.3969/j.issn.0517-6611.2022.02.028
开放科学(资源服务)标识码(OSID):
Effects of SbGA20ox1,SbGA20ox3 and SbGA2ox3 Genes on the Expression of Gibberellin in Sorghum Varieties
SHANG Jing,LU Jin-yu,PANG Hong-bo et al (School of Life Sciences,Shenyang Normal University,Shenyang,Liaoning 110034)
Abstract [Objective]To study the effects of SbGA20ox1,SbGA20ox3 and SbGA2ox3 on the expression of gibberellin in sorghum varieties.[Method]Different sorghum varieties were used as test materials (9544.7057 was used as the male parent and different female parents were crossed to obtain offspring:Liaoxialiang 1,Liao 2297,Liao 2697,Liao 5397,01-26B;and 01-26A was used as the female parent crossing with different male parents to obtain offspring:Liaoza 35,0-01,9544.7057,3550,Liao 2697). Real-time fluorescent quantitative PCR technology was used to compare the expression levels of SbGA20ox1,SbGA20ox3,and SbGA2ox3 genes between different periods (joining stage,heading stage,maturity stage) and different varieties of sorghum.[Result] The gene expression pattern of SbGA20ox1 gene in the same male parent and different female parent sorghum groups basically accorded with heading date>mature stage>joining stage;the gene expression law of SbGA2ox3 gene in the same male parent and different female parent sorghum groups basically accorded with heading date>mature stage>joining stage,the gene expression pattern of SbGA2ox3 gene in the same female parent and different male parent sorghum groups basically accorded with jointing stage>mature stage>heading stage. However,the gene expression of SbGA20ox1 gene in the same female parent and different male parent sorghum groups and the gene expression of SbGA20ox3 gene in various varieties of sorghum did not find obvious regularities,and the relative expression levels of genes in each period showed no significant difference among most varieties.[Conclusion]This study revealed to a certain extent the gene expression regularity of SbGA20ox1,SbGA20ox3 and SbGA2ox3 genes in multiple sorghum varieties at various stages,and provided enlightenment for further research on the mechanism of the small gene family that regulates gibberellin levels.
Key words Sorghum;Relative expression level;Real-time fluorescence quantification;Gibberellin;Small gene family
基金项目
辽宁省教育厅重点攻关项目(LZD201901);辽宁省教育厅自然科学重点攻关项目(LZD202004);财政部和农业农村部国家现代农业产业技术体系(CARS-06)。
作者简介 商靖(1996—),女,辽宁北票人,硕士研究生,研究方向:植物基因工程。*共同通信作者:王艳秋,研究员,硕士,从事高粱育种与栽培工作;李玥莹,教授,博士,从事植物基因工程研究。
收稿日期 2021-04-16
高粱属禾谷类作物,是世界第五大作物[1-2]。作为粮食,高粱的籽粒内含有丰富的营养物质,如淀粉、蛋白质、铁、镁等[3],且在边缘土地上,相较于其他可作为粮食的作物,生产和生物量更高[4],在非洲和很多发展中国家的许多干旱及半干旱地区都是重要食物来源[5]。高粱作为粮食已有悠久的历史。除了可作为粮食,高粱还具有许多其他用途。比如作为饲料,可以最小化差异地替代玉米,进而更节约成本[6]。在酿酒中,相较于小麦、大米、糯米等,高粱整体协调风格评价最高[7];此外,高粱还可以作为能源[8],具有很高的利用价值 [9-10]。
赤霉素是植物激素,属于双萜类化合物 [11],是高等植物重要的五大内源激素之一[12] ,也是重要的植物生长调节剂[13]。赤霉素能够促进细胞的伸长[14],并参与植物生长发育的许多方面,如种子萌发、茎的伸长以及植物籽粒生物量的提高等[4]。
植物中赤霉素的水平受许多小基因家族的表达的调控。如2-氧化戊二酸-依赖加双氧酶赤霉素20-氧化酶1(SbGA20ox1)基因和2-氧化戊二酸-依赖加双氧酶赤霉素20-氧化酶3(SbGA20ox3)基因对赤霉素水平有正调节作用,2-氧化戊二酸-依赖加双氧酶 赤霉素2-氧化酶3(SbGA2ox3)基因对赤霉素水平有负调节作用[15]。
该试验探究SbGA20ox1、SbGA20ox3和SbGA2ox3基因在不同品种高粱各时期的基因表达量规律,为进一步探究赤霉素受小基因家族的调控机制提供参考。
1 材料与方法
1.1 材料 供试高粱品种由辽宁省农科院提供,分别是以9544.7057为同一父本的子代,辽夏梁1号、辽2297、辽2697、辽5397、01-26B 5个高粱品种,以及以01-26A为同一母本的子代,辽杂35、0-01、9544.7057、3550、辽2697 5个高粱品种。
1.2 方法
1.2.1 RNA的提取及cDNA的合成。取约30 mg样品于已放入20颗研磨用小钢珠的2 mL离心管中,使用研磨仪进行样品研磨,随后根据RNA提取试剂盒 (Promega) 说明书,进行总RNA的提取,随后根据 PrimeScript RT Master Mix (TaKaRa) 试剂盒说明书,进行 RNA 的反转录,合成 cDNA 备用。
1.2.2 引物的选取。参照文献[16],选取SbActin基因为内参基因,另选取 SbGA2ox 1、 SbGA20ox 3、SbGA20ox 3为引物,对影响赤霉素水平的SbGA20ox1、SbGA20ox3和SbGA2ox3基因进行基因表达检测。引物由生工生物工程上海 (股份) 有限公司合成,具体见表1。
1.2.3 PCR扩增。使用SYBR Premix Ex Taq Ⅱ (TaKaRa) 进行 RT—PCR 扩增。
PCR反应体系为20.0 μL:10.0 μL 2× SYBR Premix Ex Taq Ⅱ,0.8 μL Forward Primer (10 μmol/L),0.8 μL Reverse Primer (10 μmol/L),6.4 μL ddH 2O,2.0 μL cDNA模板。
PCR反应参数为:95 ℃ 30 s;95 ℃ 5 s,60 ℃ 30 s,72 ℃ 20 s 40次循環。反应结束后,电脑自动生成Ct值[17]。
1.2.4 基因表达检测。得到的Ct值用Comparative Delta-delta Ct法进行数据分析,使用IBM SPSS 23软件进行方差分析和显著性检验[18],使用Origin 8.6软件进行柱状图的绘制。
2 结果与分析
2.1 SbGA20ox1基因在不同高粱品种各时期的基因表达规律 对同一父本9544.7037的辽夏梁1号、辽2297、辽2697、辽5397、01-26B的测定分析结果表明,拔节期,SbGA20ox1基因表达量在这5个高粱品种中的表达量都非常低,且品种间无显著差异。抽穗期,辽2697的SbGA20ox1基因表达量最高(6.28),辽5397的SbGA20ox1基因表达量最低(1.5)。成熟期,辽2297的SbGA20ox1基因表达量最高(7.99),辽5397的SbGA20ox1基因表达量最低(0.4)。试验结果还显示,除品种辽2297外,辽夏梁1号、辽2697、辽5397、01-26B,4个品种SbGA20ox1基因表达量均符合抽穗期>成熟期>拔节期。而品种辽2297的SbGA20ox1基因表达量为成熟期>抽穗期>拔节期(图1 a)。
对同一母本01-26A的辽杂35、0-01、9544.7057、3550、辽2697的测定分析结果表明,拔节期,9544.7057的SbGA20ox1基因表达量最高(6.02),显著高于其他4个品种,0-01的SbGA20ox1基因表达量最低(0.05)。抽穗期,3550的SbGA20ox1基因表达量最高(1.0),0-01的SbGA20ox1基因表达量最低(0.09)。成熟期,辽2697的SbGA20ox1基因表达量最高(2.6),辽杂35的SbGA20ox1基因表达量最低(0.16)。辽杂35、9544.7057在拔节期的基因表达量尤其高外,其他品种均为成熟期的基因表达量最高,但相比于其他2个时期没有很大差异(图1 b)。
2.2 SbGA20ox3基因在不同高粱品种各时期的基因表达规律 对同一父本9544.7037的辽夏梁1号、辽2297、辽2697、辽5397、01-26B的测定分析结果表明,拔节期,辽2697的SbGA20ox3基因表达量最高(2.0),辽2297的SbGA20ox3基因表达量最低(0.31)。抽穗期,辽2697的SbGA20ox3基因表达量最高(7.84),与其他4个品种表达量相差较大,辽2297的SbGA20ox3基因表达量最低(0.24)。成熟期,辽2697的SbGA20ox3基因表达量最高(1.52),略高于01-26B,辽5397的SbGA20ox3基因表达量最低(0.23),略低于辽夏梁1号(图2 a)。
对同一母本01-26A的辽杂35、0-01、9544.7057、3550、辽2697的测定分析结果表明,拔节期,0-01的SbGA20ox3基因表达量最高(13.43),显著高于其他品种,辽杂35的SbGA20ox3基因表达量最低(1.29)。抽穗期,辽杂35的SbGA20ox3基因表达量最高(3.45),9544.7057的SbGA20ox3基因表达量最低(0.39)。成熟期,0-01的SbGA20ox3基因表达量最高(6.06),辽2697的SbGA20ox3基因表达量最低(0.79)(图2 b)。
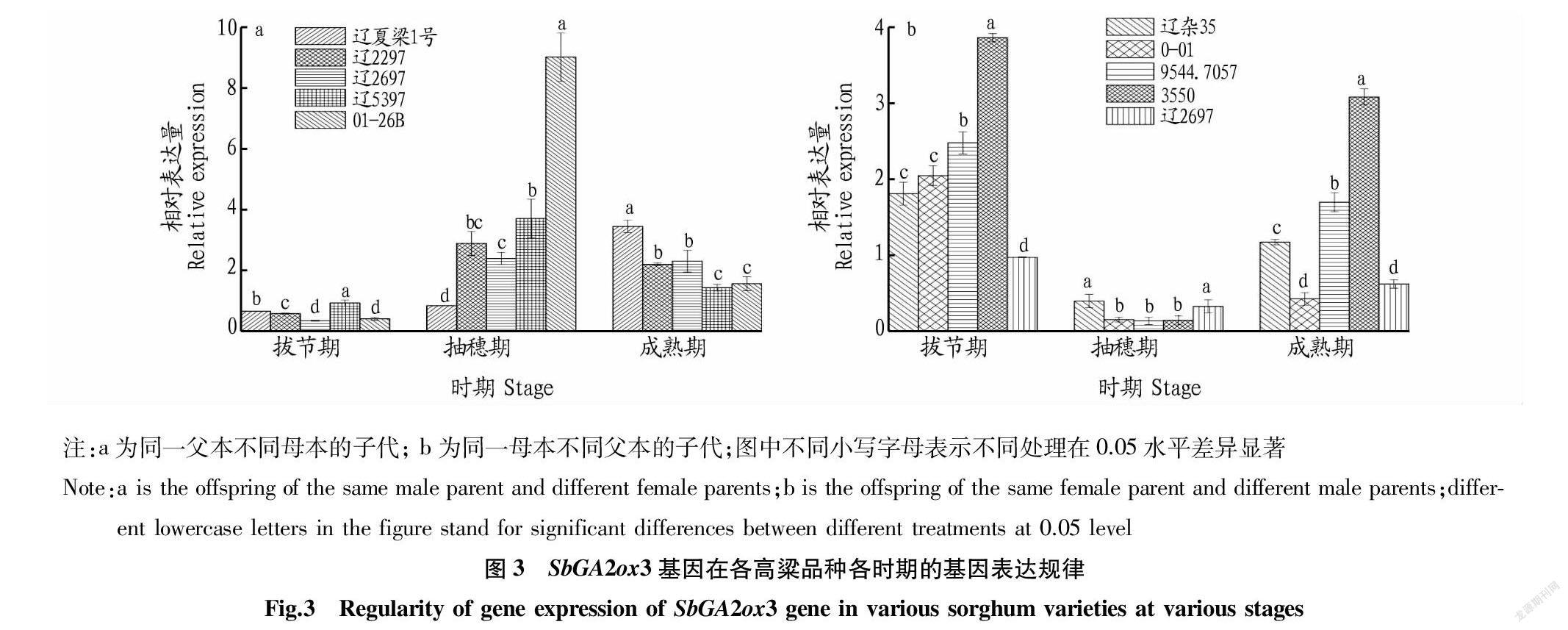
2.3 SbGA2ox3基因在不同高粱品种各时期的基因表达规律 对同一父本9544.7037的辽夏梁1号、辽2297、辽2697、辽5397、01-26B的测定分析结果表明,拔节期,辽5397的SbGA2ox3基因表达量最高(0.95),辽2697的SbGA2ox3基因表达量最低(0.37)。抽穗期,01-26B的SbGA2ox3基因表达量最高(9.01),显著高于其他4个品种;辽夏梁1号的SbGA2ox3基因表达量最低(0.86)。成熟期,辽夏梁1号的SbGA2ox3基因表达量最高(3.46),辽5397的SbGA2ox3基因表达量最低(1.45)。试验结果还显示,除辽夏梁1号外,辽2297、辽2697、辽5397、01-26B的基因表达量均符合抽穗期>成熟期>拔节期。而辽夏梁1号的基因表达量符合拔节期<抽穗期<成熟期(图3 a)。
对同一母本01-26A的辽杂35、0-01、9544.7057、3550、辽2697的测定分析结果表明,拔节期,3550的SbGA2ox3基因表达量最高(3.86),辽2697的SbGA2ox3基因表达量最低(0.98)。抽穗期,辽杂35的SbGA2ox3基因表达量最高(0.40),9544.7057的SbGA2ox3基因表达量最低(0.14),但与品种3550、0-01的SbGA2ox3基因表达量相差无几。成熟期,3550的SbGA2ox3基因表达量最高(3.09),0-01的SbGA2ox3基因表达量最低(0.43)。试验结果还显示,辽杂35、0-01、9544.7057、3550、辽2697 5个品种的高粱SbGA2ox3基因表达量均符合拔节期>成熟期>抽穗期(图3 b)。
3 讨论
高粱可以用作多种营养食品,其功能特性和作为食品的应用已广为人知[5]。 赤霉素在非生物胁迫耐受性中起重要作用[19]。已有研究表明,赤霉素能促进细胞的分裂和伸长,且在植物的整个生命周期内都起作用[9]。而赤霉素在植物各时期的水平受许多小基因家族调控,其中SbGA20ox1基因和SbGA20ox3基因对于赤霉素水平有正调节作用,SbGA2ox3基因对于赤霉素水平有负调节作用。
该研究表明,SbGA20ox1基因在相同父本不同母本高粱组,拔节期的基因表达量很低,且除辽2297品种外均符合抽穗期>成熟期>拔节期规律。SbGA20ox1基因在相同母本不同父本高粱组的3个时期,表达量差异不大,无明显规律。SbGA20ox3基因在相同父本不同母本高粱组,除品种辽2297外的品种,拔节期的基因表达量最高,但各時期间基因表达量的差异并不大。SbGA20ox3基因在相同母本不同父本高粱组,除辽杂35外的高粱品种在拔节期的基因表达量最高。SbGA2ox3基因在相同父本不同母本高粱组,拔节期的基因表达量偏低,除辽夏梁1号外,其余高粱品种基因表达量均符合抽穗期>成熟期>拔节期。SbGA2ox3基因在相同母本不同父本高粱组,抽穗期的基因表达量很低,5个高粱品种基因表达量均符合拔节期>成熟期>抽穗期。
参考文献
[1]
李玥莹,李春阳,邢慧清,等.高粱抗丝黑穗病SCAR标记的建立[J].沈阳师范大学学报(自然科学版),2016,34(4):468-472.
[2] 邓小锋,彭秋,李青风,等.高粱炭疽病抗性机理研究进展[J].贵州农业科学,2019,47(11):68-74.
[3] 张瑞栋,张壮,岳忠孝,等.播期对高粱营养与食味品质的调控效应[J].中国生态农业学报,2020,28(3):386-394.
[4] WANG Y L,SUN J Z,ALI S S,et al. Identification and expression analysis of Sorghum bicolor gibberellin oxidase genes with varied gibberellin levels involved in regulation of stem biomass[J/OL].Industrial crops and products,2020,145[2020-11-07].https://doi.org/10.1016/j.indcrop.2019.111951.
[5] ADEBO O A.African sorghum-based fermented foods:Past,current and future prospects[J].Nutrients,2020,12(4):1111.
[6] THOMAS L L,ESPINOSA C D,GOODBAND R D,et al. Nutritional evaluation of different varieties of sorghum and the effects on nursery pig growth performance [J].Journal of animal science,2020,98(5):1-16.
[7] 江伟,韦杰,李宝生,等.不同原料酿造单粮白酒风味物质特异性分析[J].食品科学,2020,41(14):234-238.
[8] 艾雪莹.种植密度和配置方式对高粱生长发育及微环境的影响[D].沈阳:沈阳农业大学,2019.
[9] 李佳衡.高粱形态及生理特性对不同肥料的响应[D].沈阳:沈阳农业大学,2019.
[10] 寇兴凯,徐同成,宗爱珍,等.高梁的营养价值以及应用现状[J].安徽农业科学,2015,43(21):271-273.
[11] 高秀华,傅向东.赤霉素信号转导及其调控植物生长发育的研究进展[J].生物技术通报,2018,34(7):1-13.
[12] 邢丹,黄勇兰,赵洪,等.参薯赤霉素途径基因的克隆与表达分析[J].分子植物育种,2020,18(6):1755-1761.
[13] 安迪.赤霉素对兔眼蓝莓生长发育的影响研究[D].杭州:浙江农林大学,2018.
[14] 陈子奇. 利用钝化内源赤霉素策略创制半矮秆玉米[D].哈尔滨:哈尔滨师范大学,2019.
[15] GAO S J,XIE X Z,YANG S G,et al.The changes of GA level and signaling are involved in the regulation of mesocotyl elongation during blue light mediated de-etiolation in Sorghum bicolor[J].Molecular biology reports,2012,39(4):4091-4100.
[16] RODRGUEZ M V,MENDIONDO G M,CANTORO R,et al. Expression of seed dormancy in grain sorghum lines with contrasting pre-harvest sprouting behavior involves differential regulation of gibberellin metabolism genes[J].Plant and cell physiology,2012,53(1):64-80.
[17] 高华晨.不同高粱品种抗氧化指标及内源激素分析[D].沈阳:沈阳师范大学,2019.
[18] 郑月英,魏胜娟,董书圣,等.梅山豬前体脂肪细胞的分离培养及分化相关基因的表达规律研究[J].畜牧与兽医,2017,49(6):9-14.
[19] EMAMVERDIAN A,DING Y L,MOKHBERDORAN F.The role of salicylic acid and gibberellin signaling in plant responses to abiotic stress with an emphasis on heavy metals[J].Plant signaling & behavior,2020,15(7):1-10.
