创造性思维的酝酿效应*
2022-02-18李子逸
李子逸 张 泽 张 莹 罗 劲
·研究前沿(Regular Articles)·
创造性思维的酝酿效应*
李子逸 张 泽 张 莹 罗 劲
(首都师范大学心理学院, 北京 100048)
酝酿效应是指当人们遭遇无法解决的难题时, 暂时将其搁置, 转而去完成其他无关任务, 反而有利于原有问题解决的现象。近年来, 研究者对创造性问题解决过程中的酝酿效应及其影响因素进行了广泛的研究, 同时提出了较多理论试图进一步解释酝酿效应。酝酿效应的代表性理论可分为“选择性遗忘理论”、“激活扩散和线索同化理论”、“无意识加工理论”等, 它们从不同角度解释了酝酿效应的机制, 并对其影响因素和有关现象各有预测。对心智游移和睡眠这些特殊形式的酝酿的研究进一步揭示了酝酿效应的机制。认知神经研究表明, 酝酿期的基本认知成分, 即表征重构和激活扩散, 涉及到前额叶、纹状体、海马体等多个脑区的协同合作。未来研究需要以深化理论建设为基础, 利用认知神经方法进一步揭示酝酿效应的机制, 从而更为深刻地认识创造性酝酿的本质。
酝酿效应, 无意识思维, 创造力
1 引言
“我离开了当时居住的卡昂, 在矿业学校的赞助下进行了一次地质考察。旅行的变动使我忘记了我的数学工作。抵达库唐斯后, 我们坐上一辆公共汽车要到某个地方去。就在我踏上车的那一霎那, 我突然有了一个想法, 这个想法在我此前的思想中没有任何铺垫, 那就是我用来定义富克斯函数的变换与非欧几何的变换是相同的。[…]回到卡昂后, 我不慌不忙地验证了结果。” 1910年, 庞加莱在一文中回忆了这段经历。基于对自身经验的反思, 他将这种突然的顿悟视为一段长期潜意识工作的结果(Poincaré, 1910)。
这段神秘的时期, 被定义为“酝酿期”。在20世纪前半叶, 华莱士(Wallas)提出的创造力的四阶段模型得到了广泛的认可, 他认为创意产生需要经历4个时期:准备期, 酝酿期, 明朗期和验证期(Wallas, 1926)。在酝酿期, 个体将难以解决的问题暂时放到一边, 转而从事一些其他无关的活动, 当再次尝试解决这个问题时, 会发现问题的答案逐渐明朗, 这一现象被称为“酝酿效应”。20世纪下半叶开始, 心理学家开始以实证研究的方式检验酝酿效应是否存在, 及其影响因素。随着实证研究数量的积累, Sio和Ormerod (2009)针对117个酝酿效应研究进行了元分析, 证实了酝酿效应的存在, 也发现了一些调节酝酿效应的因素, 例如准备期的长度、酝酿期内插任务的认知负荷、目标任务类型等。此外, 对酝酿效应的理论探讨也有丰富的成果。研究者或称酝酿效应来自于固着的遗忘(Smith & Blankenship, 1991), 或称在酝酿后被试对问题有了崭新的视域(Segal, 2004; Smith, 1995; Storm & Angello, 2010), 或称酝酿改变了对问题的心理表征(Brodt et al., 2018), 或称在酝酿期发生了有利于问题解决的激活扩散(Seifert et al., 1995; Yaniv & Meyer, 1987)。此外, 对酝酿效应神经机制的研究也方兴未艾。例如, Haarmann等人(2012)发现, 在酝酿期增加被试右后脑区的α波有助于随后远距离联想测验(RAT; remote associate task)的解决, 原因可能是警醒而放松的状态有利于远距离联系的内隐激活。值得注意的是, 近年来酝酿效应研究开始与心智游移, 睡眠相结合, 为我们进一步认识酝酿效应提供了新的视角。例如, Yamaoka和Yukawa (2016)发现在酝酿阶段较多的走神预示着在创造性问题解决中更高的灵活性。
对酝酿期这一创造性思维关键阶段的研究, 既有利于揭示创造力产生过程的奥秘, 也有利于促进创造力研究的应用, 因此, 本文基于过去20年间创造力酝酿研究领域取得的研究进展, 从不同理论视角对创造性思维过程中的酝酿效应及其影响因素进行综述, 并从理论基础、研究内容、研究手段以及提高研究结果的生态效度等方面对未来研究方向进行了展望, 以便更好的理解创造力问题解决过程中的酝酿效应及其机制。
2 酝酿效应的理论
现存的酝酿效应理论主要包括遗忘固着理论(Smith & Blankenship, 1989)、注意撤回理论(Segal, 2004)、间断性有意识工作理论(Weisberg, 2006)、疲劳恢复理论(Smith & Dodds, 1999)、激活扩散理论(Yaniv & Meyer, 1987)、线索同化理论(Seifert et al., 1995)、无意识工作理论(Ritter & Dijksterhuis, 2014)等。绝大多数学者认可从无意识角度研究酝酿效应的取向, 认为间断性有意识工作或从疲劳中恢复并非酝酿效应之来源(Gilhooly, 2016; Sio & Ormerod, 2009)。因此本文将评述本领域有代表性的几个理论, 并以之为中心总结相关研究结果(见表1)。
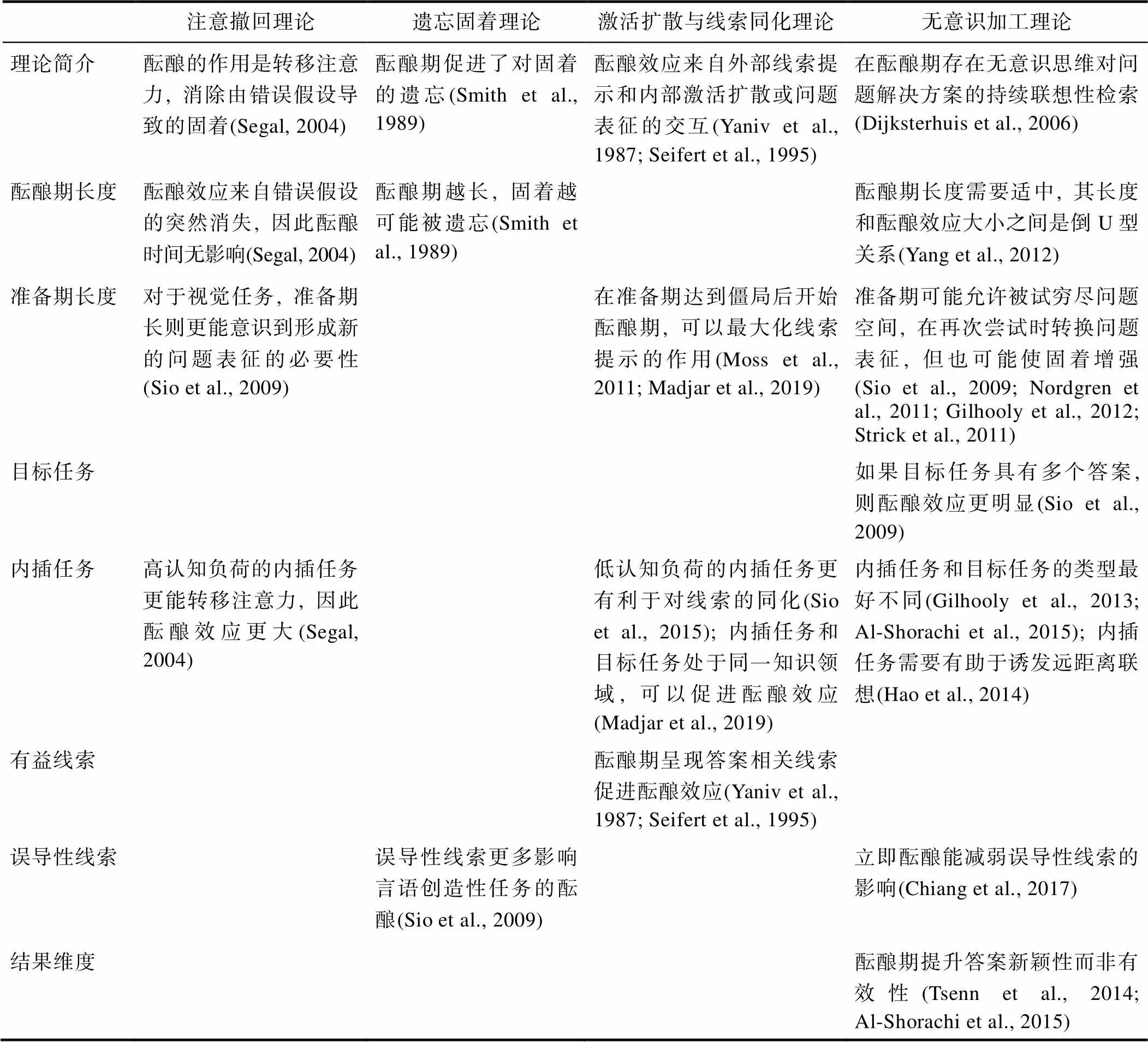
表1 酝酿效应理论及相关实证研究
2.1 选择性遗忘理论
选择性遗忘理论包括注意撤回理论和遗忘固着理论, 这些理论都认为酝酿期的作用是让个体遗忘掉前期解决问题所提出的错误假设和无效信息所导致的思维固着, 因此酝酿后个体能够抛弃旧的策略, 摆脱思维定势, 以更新颖有效的方法解决问题。
2.1.1 注意撤回理论
Segal (2004)通过分析顿悟谜题(insight puzzle)的解决过程, 指出在初次遇到问题时, 个体会形成假设, 将问题的全部要素结合起来, 以形成一个封闭的问题空间。当假设错误时, 个体的注意力受困于错误的问题空间, 从而产生固着。为了解决问题, 个体需要将由错误假设主导的注意力撤回, 以便遗忘旧假设, 重组问题空间。Segal认为在酝酿阶段什么也没有发生, 酝酿期的功能仅是将注意力转移, 并因此降低错误假设的激活, 使问题的元素重新处于无组织的状态, 由于此前的错误假设已导致其陷入思维僵局, 因此酝酿后个体可能会换用全新的假设, 形成全新的问题表征, 这一全新的问题表征更有利于随后的问题解决。
这一理论预期:相比于低认知负荷的内插任务, 高认知负荷的内插任务更能增加酝酿效应。Segal的实验发现当内插任务占用个体较多认知资源, 造成更大认知负荷时, 产生的酝酿效应更大。这是因为相比于那些轻松的分心任务, 占用较多认知资源的高负荷任务更有助于转移被试注意力, 从而遗忘错误假设, 促进酝酿效应的产生。
此外, Segal 认为酝酿期长度不影响酝酿效应。他发现, 较长或较短的酝酿期(12min, 4min)产生的酝酿效应基本相同, 造成这一现象的原因可能是酝酿期并不存在对原有任务的加工, 酝酿效应只来自错误假设的突然消失, 因此酝酿期长度并不会影响酝酿效应的强弱。
2.1.2 遗忘固着假说
Smith和Blankenship (1989)提出遗忘固着假说(forgetting-fixation hypothesis), 认为在初次问题解决尝试中, 个体从记忆中提取了错误的信息或策略, 而酝酿期减弱了错误信息的激活, 使得正确信息更加可及。
此理论认为酝酿期时间越长, 误导信息消退越多, 酝酿效应也越大。Smith等人将被试随机分配到不同的酝酿条件下, 分别是无酝酿期条件(酝酿时长0 min), 较短酝酿期条件(酝酿时长5 min)和较长酝酿期条件(酝酿时长15 min), 结果发现经历较长酝酿期的被试完成字谜任务(rebus puzzle)的成绩要显著好于短酝酿期条件和无酝酿条件的被试。这可能是是由于酝酿时间的增加, 被试对错误线索的记忆率降低, 酝酿效应是因为被试在酝酿期遗忘掉错误信息而产生的。
此外, 这一假说还预期:酝酿效应仅发生在被试陷入固着的情况下, 若未被错误线索固着, 则没有酝酿效应。这一假设虽然也得到了一些实证研究的支持(Choi & Smith, 2005; Smith & Blankenship, 1991), 但Sio等人(2009)的元分析显示, 是否给予误导性线索总体上并不影响酝酿效应。这可能是因为存在调节变量。在上述元分析中, 有25项研究运用言语创造性任务, 4项研究运用视觉创造性任务, 但是, 误导性线索的存在仅影响前者的酝酿效应, 即只有在使用言语创造任务时, 个体才通过遗忘错误信息来产生酝酿效应。这也说明当问题材料涉及语言而非视觉刺激时, 摆脱线索的误导对成功的问题解决更加关键。
2.2 激活扩散和线索同化理论
激活扩散理论和线索同化理论关系密切, 它们都强调外部环境和内部无意识过程的交互:在酝酿期可能既有原有问题表征的激活, 也有在语义网络中的扩散激活, 而此二者均可能与外部线索发生互动(Sio & Ormerod, 2015), 从而促进问题解决。
2.2.1 激活扩散理论
Yaniv等人(1987)提出酝酿的激活扩散说, 认为解题失败可能会部分激活对解决问题至关重要的记忆痕迹, 在酝酿期, 这些痕迹在语义网络中扩散, 从而激活先前被忽视的关键知识, 这些知识互相结合, 从而促进问题解决。
与遗忘−固着理论不同, 激活扩散说认为:较长的酝酿期不一定导致更强的酝酿效应, 最佳的酝酿期长度可能取决于激活项的数量及其衰减速率。Sio等人(2017)发现分散作业(被试同时解决3个远距离联想问题, 每个10 s)的效果优于集中作业(被试持续解决1个问题, 每个30 s), 且分散作业适合10分钟的酝酿期, 集中作业适合5分钟的酝酿期。这可能是因为同时呈现的项目虽然很快被激活到更高的水平, 但它们的衰减速度更快; 相反, 继时呈现的问题激活水平较低, 但其衰减得较慢。因此对于分散作业, 将远距离概念激活到足以促进问题解决的程度需要较长的酝酿期, 而对于集中作业, 较短的酝酿期则更有利。
2.2.2 线索同化理论
Seifert等人(1995)提出酝酿的线索同化说, 认为在准备期遇到的僵局使问题表征保持了一定程度的激活, 增加了同化外部线索的可能性, 当二者交互, 问题解决的可能性就会上升。
线索同化理论预期:在酝酿期呈现有益线索会提升酝酿效果。但Sio等人(2009)对10项相关研究的分析没有发现有益线索的调节作用。近年也有研究发现, 在第二次尝试解决问题的前3 s, 呈现与答案语义相关的线索并不能增加解题率(Smith et al., 2012)。可以确定的是, 呈现外部线索并不总是有利的, 存在许多调节变量。例如, Moss等人(2011)发现, 在准备期达到僵局不久后, 立刻开始酝酿, 可以最大化线索的提示作用, 因为此时个体最可能利用新信息以探索新的问题表征。反之, 过早酝酿可能导致问题表征未充分建立, 问题的激活程度不足。过晚酝酿则可能导致随着准备期的延长, 固着进一步增强。此外, Sio和Ormerod (2015)发现, 当尝试解决困难远距离联想问题时, 认知负荷低的内插任务更有利于线索发挥作用, 原因可能是高负荷的任务不利于无意识思维对线索加以处理。
线索同化理论还认为:较长的准备期, 以及内插任务和目标任务是否属于同一知识领域将影响酝酿效应强弱。Madjar等人(2019)让被试自主决定什么时候结束准备期。他们发现, 在目标任务处理过程的晚期开始酝酿, 可以获得更强的酝酿效应。说明个体需要足够长的准备期以陷入思维僵局, 这样才能增加在内插任务中同化偶然外部线索的可能性。另外, 他们认为, 内插任务和目标任务如果处于同一知识领域(例如设计新的营销活动和分析市场数据), 将促进远距离联结和信息整合。在同一知识领域中进行内插任务, 可以建立充分的远距离联想, 同时最小化重新执行目标任务的认知转换成本, 便于将激活的知识重新转移回目标任务, 因而能得到更强的酝酿效应。
2.3 无意识加工理论
Dijksterhuis和Nordgren (2006)提出无意识加工理论(unconscious-thought theory), 认为在酝酿期, 当意识被导向给分心任务时, 与目标任务有关的无意识思考过程依然在继续。Dijksterhuis将被试分配到(1)无酝酿组:要求被试在1分钟之内立刻解决发散性思维任务; (2)有意识思考组:被试看到目标任务后先有意识的思考3分钟, 随后再作答; (3)无意识思考组:被试看完目标任务后, 需要先完成另一项持续3分钟的分心任务, 之后再完成目标任务。结果发现无意识思考组的被试完成发散思维任务的表现显著好于其他两组(Dijksterhuis & Meurs, 2006; Study 3), 说明酝酿效应可能来自酝酿期被试对目标任务的无意识加工。Zhong等人(2008)用词汇决策任务(lexical decision task; Bargh et al., 1995)发现, 这种了解目标任务后立即开始的酝酿虽然没能增加远距离联想测验的解题率, 但是却内隐激活了未解题目的答案。Zhong因此认为:在酝酿期, 无意识思维促进对创造性答案的联想性检索, 进而不断增加答案的无意识激活程度, 直到进入意识层面使问题得到解决。
需要指出的是, 与以往的延迟酝酿范式(初次尝试−酝酿−二次尝试)不同, 依据无意识工作理论发展的立即酝酿范式(了解目标任务−酝酿−执行目标任务)抛弃了在酝酿前的第一次尝试。有研究者认为, 立即酝酿效应很好地驳斥了酝酿的遗忘理论, 因为在立即酝酿范式中, 并没有需要忘记的固着, 这就说明酝酿效应不是来源于固着的减弱(Gilhooly, 2016)。然而, Chiang等人(2017; Study 2)使用了立即酝酿范式, 却发现酝酿仍然能减弱错误线索, 即固着的影响。说明思维僵局可能在早期知觉阶段, 即观看目标任务时, 而不是在晚期加工阶段, 即在准备期陷入僵局时, 就已经产生(沈汪兵等, 2012)。
无意识加工理论可以解释一些酝酿效应的影响因素。第一是目标任务的类型。此理论假设:如果目标任务具有多个答案, 则擅长联想和发散的无意识加工会导致更强酝酿效应。Sio等人(2009)将酝酿效应研究中涉及的目标任务划分为3种:创造性问题(例如发散性的结果问题, “如果每个人都失去了读写能力, 会发生什么?”), 言语顿悟问题(例如远距离联想测验)和视觉顿悟问题(例如九点问题)。这些任务或要求尽可能多地激活不同概念, 或要求重构错误或不完备的问题表征。元分析显示相比顿悟问题, 创造性任务更易受到酝酿的积极影响, 说明酝酿效应更多发生在需要广泛激活不同概念的问题上。此外, 研究还显示, 酝酿效应更能提升答案的新颖性而不是有效性(Tsenn et al., 2014; Al-Shorachi et al., 2015), 这也符合无意识思维的发散性特征。
第二是内插任务和目标任务的类型。此理论预期, 如果内插任务和目标任务的类型不同, 则个体在酝酿期将分配更多的认知资源对目标任务进行持续的无意识加工, 酝酿效应也越强。Al-Shorachi等人(2015)发现, 在发散性的想法生成任务之间插入数学测验而不是讲故事任务更有利于酝酿效应。Gilhooly等人(2013)发现, 目标任务类型(语言或空间)和内插任务类型(语言或空间)之间存在交互作用。在酝酿期完成和目标任务性质不同的分心任务更有利于酝酿效应。Gilhooly称这些结果驳斥了注意撤销假说和遗忘固着假说。前者预期内插活动与目标任务的相似性不应该有任何影响, 因为内插任务只需要能转移注意力即可。后者则预期使用与目标任务类似的内插任务更佳, 因为类似的任务将产生更多基于干扰的遗忘。
第三是内插任务的负荷程度和性质。此理论预期认知负荷过高会减弱酝酿效应。Sio等人(2009)发现, 相比于高负荷任务, 低负荷内插任务更有利于言语顿悟问题的酝酿。他们猜测, 这可能是因为低负荷的任务中, 被试的注意力更分散, 更适合检索较远的语义联结。此外, Hao等人(2014)选取反思准备期答案和字谜任务这两个负荷高低不同, 但均可以诱发远距离联想过程的内插任务, 发现二者对发散性言语任务均有同等酝酿效果, 说明内插任务是否有利于无意识的发散思考也是一个关键因素。
最后是酝酿期长度。Yang等人(2012)将被试分为有意识思维组和无意识思维组。对于有意识思维组, 分别给1分钟、3分钟或5分钟的时间持续思考目标任务; 对于无意识思维组被试, 则分别完成1分钟、3分钟或5分钟的分心任务。结果发现, 只有在思维时间为3分钟时, 无意识思维组的表现才优于有意识思维组, 即无意识思维的时间和创造力之间呈现倒U型关系。因此, 太早或者太晚结束酝酿期, 都不利于酝酿效应的产生。
3 心智游移与睡眠的酝酿效应
酝酿效应是受到各类因素(例如内插任务的性质, 准备期的长度等)影响的复杂过程。不同的酝酿理论对它们各有预测, 例如, 注意撤回理论认为酝酿期的长度不会影响到酝酿效应, 而遗忘固着理论则认为酝酿期需要足够长。此外, 有一些因素是很多理论都涉及的, 如酝酿期长度, 准备期长度等, 涉及到它们的实证研究可以借助相关理论加以解释; 而有些因素则只有非常少的理论存在预测, 如有益线索的作用一般只用线索同化理论加以解释。
然而, 近年来酝酿效应的研究领域开始和心智游移、睡眠相结合。这些研究的成果很多难以用上述理论良好地解释, 而更应该结合心智游移和睡眠的本身特点来加以解释。此外, 对于心智游移和睡眠是否能促进酝酿效应的产生, 本身也存在互相矛盾的证据。
3.1 心智游移
心智游移(mind wandering)指与个体当前正在从事的任务无直接关系, 并且不是由个体有意发起的意识体验(Smallwood & Schooler, 2006; Smallwood & Schooler, 2013)。在心智游移时, 个体可能会突然想到一小时后的聚餐, 刚才没能解决的数学问题以及曾经听过的歌曲旋律等无关信息, 此类突然出现在脑海中的画面、声音、语言等, 均属于心智游移。
从理论层面上讲, 心智游移的确可能是酝酿效应的来源之一。执行分心任务时发生的心智游移能帮助个体建立更远的联结, 促进顿悟。例如, Baird等人(2012)让被试从事不寻常用途任务, 之后将被试分配至低负荷任务组、高负荷任务组、休息组、持续工作组。结果发现低负荷组在执行分心任务时, 有更高的心智游移水平, 且在酝酿后的发散性思维任务上表现更好。研究者认为心智游移如激活扩散理论预测的那样, 通过增加无意识联结处理(unconscious associative processing)来产生酝酿效应。除了上述这种无意识层面的解释, 另一种理论观点认为:酝酿期的心智游移让个体有机会在显意识层面处理目标任务(Smallwood & Schooler, 2006)。心智游移的内容往往和自我或当前所关注事物相关(Johannessen & Berntsen, 2010; Watkins, 2010), 例如, Morsella等人(2010)发现短期的任务目标能增加与目标有关的心智游移内容。说明酝酿期的心智游移可以由目标任务驱动, 它有可能服务于目标任务的解决。
然而, 目前却没有稳定的证据表明心智游移是酝酿效应产生的关键, 或者那些在酝酿期有更多心智游移的个体比其他人更能受益于酝酿期。一方面是反对性的证据。例如, Smeekens (2013)发现酝酿期的心智游移无助于顿悟问题的解决。Smeekens等人(2016)在酝酿期测量被试心智游移频率, 也发现心智游移频率与后续发散性思维表现的提高无关。Yamaoka和Yukawa (2017)在特质层面的研究结论混杂, 在创造力的不同指标上有相反的结论。在Yamaoka和Yukawa (2020)的研究中, 他们对比了频繁心智游移的个体与不频繁心智游移的个体, 发现如果严格按照统计学标准, 二者在创造力上没有显著差异, 并称结果可能是由于较少的样本量和总体上较低的心智游移频率。Steindorf等人(2020)将3种酝酿期(执行分心任务时被思维探针打断, 被琐碎的问题打断, 无打扰)和控制组对比, 既没有发现酝酿效应, 也没有发现心智游移有助于发散性思维任务的证据。
另一方面则是支持性的证据。例如, Tan等人(2015)指出Smeekens (2013)的研究中使用的问题难度过大, 存在天花板效应, 酝酿后问题的平均解决率极低, 这些可能是导致效应不明显的原因。因而他们选择难度较低的数字递减任务(NRT; Number Reduction Task)。此任务允许被试通过洞察数字排列的隐藏规律来快速解题。1小时分心任务后, 他们发现, 那些在第二次数字递减任务中成功洞察了隐藏规律的个体, 相比于未洞察者, 在酝酿期有更多心智游移。且他们在随后的创造力测验上的得分也更高。Tan等人认为该结果说明酝酿期的心智游移在顿悟问题解决中发挥了重要作用。Yamaoka和Yukawa (2016)发现在酝酿过程中走神频率更高的被试在之后的不寻常用途任务上表现出更多灵活性和独创性。Leszczynski等人(2017)发现, 酝酿期的心智游移虽然不利于分心任务的表现, 但是却有利于酝酿后的远距离联想任务表现。综上, 酝酿期的心智游移是否促进酝酿后的问题解决, 它与个体的创造力有何关系, 依然是有待解决的问题。
3.2 睡眠
睡眠是每90分钟左右的往复循环, 可以被划分为快速眼动睡眠(REM)和非快速眼动睡眠(NREM), 后者, 即NREM可以被进一步分为4个阶段, 其中阶段3和4是慢波睡眠(SWS)阶段(Stickgold, 2005)。
将睡眠与酝酿结合是近年来的一个热点。经过一段时间的睡眠, 原先没能解决的问题在睡眠后得到了解决, 这种因睡眠而产生的创造力提升也可被视为一种酝酿效应。然而, 目前的实证研究关于睡眠对创造力的促进机制有不同的看法。有研究者认为睡眠的作用在于重组问题表征。Wagner等人(2004)将接触过数字递减任务的被试分配到8小时睡眠组, 夜间清醒组, 日间清醒组, 重测时发现经过睡眠的被试更容易发现隐藏规律, 有研究者认为睡眠通过重构记忆的表征结构来促进酝酿效应。Verleger等人(2013)使用数字递减任务来研究睡眠如何导致顿悟。他们发现一晚的睡眠会使洞察到隐藏规律的参与者, 即顿悟者的数量增加两倍。且在SWS中, 顿悟者的脑电信号有特定的增加。他们认为在睡眠期间, 对记忆的再加工重构了大脑中与任务相关的表征, 这种重构促进了知识的获得。
但也有研究者支持睡眠的激活扩散说。例如, Schönauer等人(2017)使用顿悟任务来研究睡眠是否促进了记忆表征的重组。结果发现, 3小时的睡眠或者清醒酝酿都不能提升解题数量。因而, 他们认为睡眠可能不适合用于解决需要重构问题表征的任务, 而是更多支持需要激活扩散过程的问题解决。与此类似, Sio等人(2013)发现, 和清醒组或控制组相比, 睡眠能促进困难的远距离联想问题的解决, 但不促进简单问题的解决, 其原因可能是睡眠导致的概念网络的激活扩散更有利于困难的, 即需要更广泛激活扩散的, 远距离联想任务的解决。
也有研究者认同睡眠的线索同化理论, 即睡眠的酝酿效应需要一定提示或线索的参与。此类研究采用的范式并不直接检验睡眠组和清醒组的差异, 而是会在睡眠前或睡眠中增加线索提示。例如, Cai等人(2009)发现, 对于那些答案在睡眠前被启动过的远距离联想测验题目(例如, 在酝酿期前被试完成一个类比填空, Chips : Salty; Candy : S___, 答案是Sweet, 由此启动Sweet这个单词, 在酝酿后尝试找出可以和Heart, Sixteen, Cookies分别组合成词组的单词, 即Sweet), 虽然被试对启动项(例如, Sweet)的记忆情况无差异, 但那些进入REM睡眠的被试相较于其他被试(NREM睡眠或清醒)有大约40%的正确率提升。Cai等人认为, 之前被激活的节点在睡眠阶段重新激活, 并无意识地发生扩散。睡眠的作用不仅是巩固对启动项的记忆而是增强启动项的联结网络(associative network)。类似地, Sanders等人(2019)向被试提供谜题, 每个谜题都与不同的声音任意关联。当参与者睡眠时, 有一半与他们没有解决的谜题有关的声音被偷偷呈现出来以重新激活记忆。第二天早上, 参与者解决了31.7%的有提示的谜题(比无提示组提高了55%)。他们认为这种重新激活可能会增强对问题的记忆, 使解决者能够创造性地在问题元素以及相关的知识之间建立新的联系。
然而, 也有一些研究未能发现睡眠的酝酿效应。Brodt等人(2018)采用多种不同类型的任务(谜语, 视觉变化察觉, 字谜), 试图分离睡眠和清醒酝酿的效应。结果发现, 只有在使用谜语任务的时候存在清醒酝酿效应, 但没能发现睡眠促进问题解决的证据。Brodt因此建议将睡眠的功能局限在强化记忆痕迹, 从已编码信息中提取普遍元素上。与其类似, Hołda等人(2020)发现睡眠组对结构不良问题给出的答案并不比清醒组更有创造力。上述研究未能发现睡眠的酝酿效应的原因可能是因为存在一些调节变量, 例如, 被试是小憩或整晚睡眠, 研究的睡眠类型是SWS还是REM, 因变量任务是侧重激活扩散还是表征重构。目前只有少量研究考虑了可能的调节变量的作用, 例如, 例如, Beijamini等人(2021)发现10小时的自然睡眠确实具有酝酿效应(Study 1), 但是其Study 2尝试探索记忆重激活对各类睡眠的影响, 分别在慢波睡眠, 快速眼动睡眠, 清醒状态给予被试听觉层面的提示线索, 结果却没有发现组间差异。
4 酝酿效应的认知神经机制
酝酿效应背后的心理过程至少包括两个认知成分:问题表征的转换和形成远距离联结(Shen et al., 2017; Sandkühler & Bhattacharya, 2008; Weisberg, 2013)。这两个认知成分由不同的脑机制负责。例如, Shen等人(2018)对31项关于创造性思维fMRI研究进行了元分析。他们假设, 酝酿期是由失败驱动的对定势或僵局的打破过程, 在其中, 个体会经历初始错误表征的重构。而随后的明朗期则主要涉及形成新异联系。在此基础上, 研究者依据实验任务的特点将收集到的实验分类, 以揭示创造性思维四阶段的不同的认知神经机制。结果发现, 在酝酿阶段, 前额叶皮层和左侧枕中回的激活可能反映了对定势的打破、抑制无效信息、表征转换、策略转变等过程(Cerruti & Schlaug, 2009; GoeL & Vartanian, 2005; Seyed- Allaei et al., 2017; Shen et al., 2016)。而右侧海马回在明朗期激活, 用于建立远距离的语义或情境记忆联系(Luo & Niki, 2003; Milivojevic et al., 2015)。但是在实际的酝酿过程中, 抛弃错误表征或定势和构建新异联结可能难以彻底分离为两个前后阶段, 反而是酝酿期中两个互相交织的认知成分, 正如Kershaw和Ohlsson (2004)提出, 在顿悟问题解决过程中个体可能遇到多个来源的困难, 在酝酿期, 个体可能同时需要新知识的激活或重构(Sio & Ormerod, 2009)。但对于这些认知成分在人脑中如何体现, 由于目前直接研究酝酿效应神经机制的研究很少, 因此本文将从不同的理论视角出发, 梳理酝酿期的认知成分可能涉及的脑机制, 并且论述心智游移和睡眠的酝酿效应的可能的神经机制(见表2), 以推动后续研究的发展。
4.1 遗忘的视角
酝酿的选择性遗忘理论认为在执行分心任务的过程中, 错误的假设或思维定势被遗忘, 从而让个体可以有一个崭新的视域来看待问题, 或者可以提取正确的信息来解决问题。酝酿期的分心任务可以通过刷新工作记忆来促进遗忘。例如, 研究发现执行分心任务可以阻止海马或海马旁回的记忆编码过程(Mullally & O’Mara, 2013)。这种抑制过程是由前额叶皮层, 尤其是背外侧前额叶调控海马体活动完成的(Anderson & Hulbert, 2020)。其次, 分心任务带来的情境改变也可以促进遗忘。有时即使是想象的情境改变, 也能移除与记忆有关的线索, 促进遗忘发生(Sahakyan et al., 2013)。而这一过程由背外侧前额叶发动, 调控海马体或内侧颞叶记忆系统的活动(Hanslmayr et al., 2012)。例如, Hanslmayr等人(2012)采用fMRI技术, 发现让被试尝试遗忘激活了左侧背外侧前额叶, 也减少了背外侧前额叶−海马体的耦合。综上, 在酝酿期, 大脑前额叶皮层对海马体等记忆系统的调控可能会负责打破定势, 遗忘错误信息, 从而促进问题解决。
4.2 线索同化的视角
酝酿的线索同化理论认为, 陷入僵局的问题在头脑中有更高的表征激活程度, 因而更能与环境中的新线索产生联系, 从而发生顿悟。许多研究指出, 对于在准备期未解决的问题, 存在特定脑区的持续激活。唐晓晨等人(2009)采用事件相关电位方法, 研究在组块破解任务中, 左右脑在解题失败与加工提示信息这两个阶段的活动差异。结果表明, 问题求解失败在右半球额叶诱发了更大的 P150 成分。在提示信息呈现阶段, 对P2成分波幅的分析表明, 右半球的活动则是由准备期是否遭遇失败来调控的。说明陷入僵局增强右脑对问题表征的保持, 并导致被试对提示更加敏感。Gao和Zhang (2014)对个体如何同化无意识线索进行了研究。他们让被试在无意识线索, 有意识线索和无线索三个条件下解决发散性问题, 结果发现给予被试无意识线索可以促进问题解决, 且整个过程依次涉及线索的语义激活(右顶下小叶, N280~340), 新联系的形成(右侧海马旁回, P350~450), 和表征转换(右侧颞上回, P500~760)。沈汪兵等人(2012)据此指出, 参与表征转换的酝酿过程可能涉及右脑额区、顶区、颞区的激活, 且酝酿期存在右半球对问题相关信息的保持增强, 这一大脑状态可能使得个体更善于接受线索所提供的新信息, 从而与已有的认知表征结合形成新的解题方案。
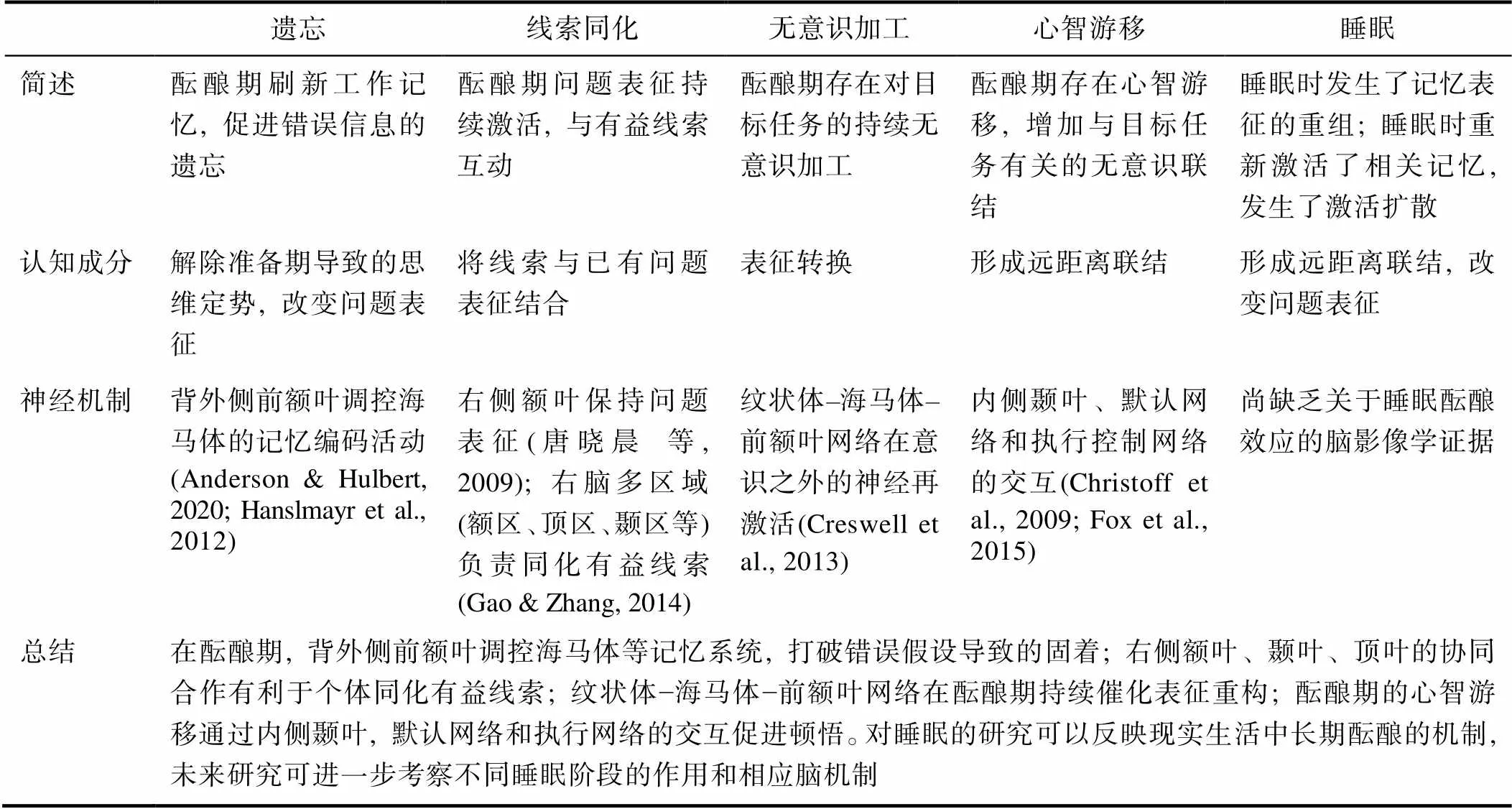
表2 各视角下酝酿期认知神经机制
4.3 无意识加工的视角
无意识加工理论认为在酝酿期存在对问题的持续无意识思考。一些研究者对负责进行酝酿期无意识思考的脑区进行了探索。例如, Darsaud等人(2011)使用数字递减任务考察延迟顿悟的脑机制。实验分为初始训练和再次测试两个部分。研究者将再次测试的第一个block和初始训练的最后一个block的脑成像数据进行比较, 认为二者的差异反映了酝酿期的离线(off-line)加工过程。结果显示, 对于那些在初始训练中未成功解决问题的被试, 在再次测试阶段如果顿悟, 腹内侧前额叶和海马体的激活会显著增强, 说明这些脑区参与内隐规则学习的离线加工。此外, Creswell等人(2013)认为无意识思维的基础是纹状体–海马体–前额叶网络在意识之外的神经再激活(neural reactivation)。他们发现纹状体和前额叶区域在决策信息的编码过程, 以及后续的分心任务中会持续激活, 并可以预测分心任务后的行为表现。这说明无意识的神经再激活可能是酝酿效应的机制。Creswell团队后续的研究检验两分钟的离线处理是否可以促进联想学习。在编码词对后, 被试或先完成两分钟分心任务, 或立即测试词对的记忆情况, 结果显示, 简短、清醒、离线的处理提升了词对记忆。此外, 多体素模式分析发现了背外侧前额叶皮层负责离线加工时记忆表征的再激活(Bursley et al., 2016)。这些脑区的激活可能改变决策选项的记忆表征(Dijksterhuis, 2013), 例如, 酝酿期的无意识思维导致选项的表征更加组织化和极化(Dijksterhuis, 2004; Bos et al., 2011)。综上, 纹状体, 海马体, 前额叶等区域很可能是负责持续催化酝酿期的表征重构。
4.4 心智游移及睡眠酝酿效应的神经机制
4.4.1 心智游移
在认知神经机制方面, 和心智游移关联最密切的是默认网络。默认网络主要包括前额叶、扣带回、颞叶内侧以及顶下小叶等区域(Buckner et al., 2008)。它是在个体在静息态时相对活跃的脑网络, 在执行目标导向任务时反而相对受抑制(Binder et al., 1999; Mazoyer et al., 2001)。
已有研究发现, 默认网络支持着心智游移。例如, Andrews-Hanna (2010)等人的研究发现默认网络在心智游移时有更高的活动强度, 且心智游移的内容可以预测颞叶内侧与默认网络其余部分的功能连接强度。因此, 心智游移促进酝酿的神经机制很可能与默认网络有关。其中需要着重关注的是心智游移和内侧颞叶(MTL)的密切相关。MTL, 尤其是海马体负责思维生成和发动(Fox et al., 2016), 而McCormick等人(2018)的研究表明, 海马体病变对心智游移有影响, 尽管病患在自我生成的思维(self-generated thought)上的频率未受影响, 但是其内容变得更加语义化, 言语化, 更少涉及过去和未来。
但除了默认网络之外, 心智游移还和执行控制网络有关。例如, Morsella等人(2010)发现, 如果在静息前给予个体之后要进行的任务目标, 个体在静息时就会出现关于这个任务的心智游移, 且只有当这个任务可以从心智游移中获益时, 相关的心智游移才会发生, 这提示心智游移可以受目标驱使。认知神经方面的研究发现, 在心智游移前存在默认网络和执行控制网络的激活(Christoff et al., 2009), Fox等人(2015)的元分析也显示, 心智游移与默认网络密切相关, 但也涉及到负责执行控制的脑区, 且在顿悟之前均可观察到这两个网络的激活(Kounios et al., 2008; Kounios et al., 2006)。Golchert等人(2017)发现, 那些经常故意走神的人表现出大脑皮层在额顶叶控制网络的厚度增加, 以及执行脑区和默认脑区之间功能连接增强。上述研究说明心智游移并非是全然不受控制的自发思维, 它很可能通过默认网络和执行系统的交互来促进酝酿效应。
Fox和Beaty (2019)综合考虑了心智游移和创造性思维之间的相似性, 提出心智游移的双过程模型, 认为和创造性思维类似, 心智游移中想法的产生和成型过程也存在生成和评价两个阶段。其中, 负责生成阶段的脑区是内侧颞叶和默认网络, 而负责想法评价阶段的则是执行网络, 或执行网络与默认网络的连接。可以假设, 在酝酿期, 以心智游移为代表的自发思维通过增强默认网络, 内侧颞叶, 执行控制网络之间的信息交换, 生成新颖有效的想法。
4.4.2 睡眠
Stickgold (2005)认为, 睡眠会增加主要的视觉和运动区域的激活, 但边缘系统, 额叶, 顶叶区域也会发生变化, 这表明睡眠可以完全改变大脑使用的策略并允许更多的自动化的任务执行, 以此提升任务表现。但究竟是睡眠的哪一个阶段最为关键, 目前尚未可知。一般来讲, 睡眠是每90分钟左右的往复循环, 可以被划分为快速眼动睡眠(REM)和非快速眼动睡眠(NREM)。NREM可以被进一步分为4个阶段, 其中阶段3和4是慢波睡眠(SWS)阶段(Stickgold, 2005)。Payne (2011)着重讨论了记忆的重新激活对不同阶段睡眠的影响, 认为在SWS期间, 记忆的重新激活会导致原有记忆的增强和稳定, 例如, 通过再现最初学习时出现的记忆线索(如气味)来重新激活记忆, 促进记忆稳定; 而REM期间的再激活会导致大量记忆碎片的分离和重新结合, 这可能会带来对问题的创造性解决方案。
REM被发现确实有助于问题解决。例如, 有研究发现, 尽管接受训练的一周后, 被试在汉诺塔任务上的表现得到提升, 但如果故意减少训练后当晚的REM睡眠, 被试表现就无法提升。Cai等人(2009)也认为, 在REM阶段记忆被重新激活, 并无意识地发生激活扩散。
也有研究发现NREM促进顿悟。例如, Verleger等人(2013)使用数字递减任务, 发现一晚的睡眠可以增加顿悟者的数量, 且那些在睡眠前内隐地察觉了任务规律的个体, 在慢波睡眠中, 10~12 Hz频段的脑电信号有特定的增加。他们认为这一结果支持在慢波睡眠阶段发生的记忆再处理能重构任务相关表征, 促进外显知识获取。
因此, 研究者们可能需要进一步探索不同的睡眠阶段对问题解决的影响及其神经机制, 也需要考虑使用的实验范式对实验结果的影响。例如, Verleger等人(2013)指出, 应当考虑实验使用的目标任务。当使用发散性思维任务, 例如Cai等人(2009)的研究时, REM可能更有利于问题解决, 这背后可能是更直观、综合的右半球模式, 而当使用顿悟任务时, SWS可能更有优势, 这背后可能是更具逻辑性、分析性的左半球模式。此外, 有fMRI研究指出了不同睡眠阶段的认知神经差异。对于涉及规则提取的任务, 在SWS期间给予有关线索, 海马体和尾状核连接增强, 在REM阶段给予有关线索, 运动皮层和小脑的激活增强(Cousins et al., 2016)。
5 讨论与展望
综上所述, 本文对创造性思维中酝酿效应理论和相关研究进行了梳理, 并根据不同的理论取向探讨了影响酝酿效应的诸多因素和可能的内在神经机制。尽管研究者已经积累了大量的证据, 但仍有一些问题亟待解决, 未来可以从以下几个方面进行拓展和深化。
5.1 提高酝酿效应研究的生态效度
目前, 针对酝酿效应的相关研究大多是在传统的实验室条件下完成的, 研究所采用的任务大多是一些经典的创造性问题(例如:远距离联想测验和多用途替代测验), 今后也许可以采用更具生态效度的测量方式, 例如Henok等人(2020)发现, 在酝酿期后与现实的人造物品互动而不是停留在纸面作答上, 更有利于酝酿效应的产生。随着智能设备的更新发展, 也可尝试使用更具生态效度的实验设计。例如, 用一些更便携的设备(例如:智能手机/手表和其他可穿戴设备等)研究更长时程或经验采样1经验采样法(experience-sampling methodology)是指在短时间多次收集个体对生活中经历的事件的即使评估, 是一种有效的纵向研究数据收集方法, 能提升生态效度, 在复杂真实环境下研究关心的变量。的酝酿效应。
酝酿效应的应用价值需要得到重视。例如, 未来研究可以考虑在群体层面检验酝酿效应以提升其应用价值。例如, 有研究发现良好线索与不良线索对个体和小团体的酝酿的影响是不同的(Smith et al., 2010), 相对于个体, 团体可能较少从酝酿中获益, 因为团体更少遗忘不良线索, 且无论是线索良好与否, 团队表现在酝酿后均有上升, 而个体则仅仅在遗忘不良线索后表现提升。此类结果将有助于酝酿效应在群体水平上的应用。最后, 也可以尝试以常人研究的视角, 探索人们在实际生活中对酝酿效应的认识, 以及运用酝酿效应的情况, 特别是酝酿在实际教育和培训领域的被重视以及被利用的情况, 以便在现实中推广和利用酝酿效应来促进创造性思维。例如Frith等人(2021)尝试研究体育锻炼对创造力表现的影响, 结果发现15分钟的跑步机行走可以提升酝酿后的发散性思维表现。Shaw (2020)尝试研究酝酿对数学领域的创造性问题解决的影响。这些研究均有助于酝酿效应在教育和培训领域的实践。
5.2 促进酝酿效应理论的进一步发展
表征转换与激活扩散, 是酝酿效应的两个基本认知成分(Sio et al., 2009)。从这一角度来看, 选择性遗忘理论和激活扩散理论并不互斥, 而是同一过程的两个部分。选择性遗忘促进打破定势, 切换问题表征, 而激活扩散过程则负责建立远距离联结。除此之外, 线索同化理论还强调了外部环境也可能促进酝酿效应, 使得酝酿效果不再是纯粹内部的认知过程。无意识加工理论则是将无意识思维在决策领域的应用扩展到酝酿领域而形成的成果, 借助无意识思维的特征来诠释酝酿效应的实证研究结果。所有这些理论虽有不同, 但却互补。未来的研究可以尝试进一步深化酝酿理论的建设, 以建立整合模型, 深入理解酝酿效应的本质。
有一些研究者尝试将意识和无意识统合起来构建理论, 以解释酝酿效应的机制(Allen & Thomas, 2011; Hélie & Sun, 2010; Yuan & Shen, 2016), 主要有显−隐交互理论(EII; explicit-implicit interaction theory)和酝酿的双过程理论(DPT; dual-process theory)。这些理论的特点在于, 声称酝酿期既存在无意识加工, 也存在有意识加工。例如, Yuan和Shen (2016)认为, 尽管无意识加工理论对发散性思维中的酝酿效应提供了一定解释, 但是在顿悟问题解决或聚合性思维的情况下, 可能有所不足。相对于发散性思维问题, 顿悟问题解决更依赖于通过重构初始问题表征, 约束放松等方法来克服思维僵局。这些过程(如摆脱思维定势)可能需要有意识的认知控制, 因此Yuan等人认为, 在酝酿期同时存在持续的无意识加工和有意识加工。并以这一理论对一些无意识加工理论难以解释的实验结果进行了解释。例如, 无意识思维和创造性表现之间的倒U型关系(Yang et al., 2012):只有在无意识过程和有意识过程间达成平衡时, 才有最佳酝酿效果。
但也存在反对意见, 例如Gilhooly (2017)回应称, 酝酿期的定义就是对目标任务不进行有意识加工, 且大量的实证研究均否认酝酿效应与有意识加工有关系(Gilhooly et al., 2012, 2013), 认为在酝酿期有意识地处理目标任务对创造性问题解决没有帮助。因此有意识思维到底在酝酿期发挥什么样的作用, 以及如何发挥作用尚需进一步探讨。
5.3 提高在实验室中复现酝酿效应的可能性
由于酝酿效应受到大量调节因素的影响, 因此在实验室中若不能严格控制各类因素, 则很可能出现无法观测到酝酿效应的情况。为了提高在实验室中复现酝酿效应的可能性, 今后的研究者至少应当考虑以下几个尚无定论的问题。
使用什么范式更容易获取酝酿效应?目前使用较广泛的两个范式是延迟酝酿范式和立即酝酿范式。Nordgren等人(2011)发现对于复杂决策任务, 延迟酝酿(2分钟有意识思考+2分钟分心任务)的效果好于立即酝酿(了解目标问题后立刻进行4分钟分心任务), 说明将意识与无意识相结合可以得到最优结果。然而, Gilhooly等人(2012)发现立即酝酿(4分钟分心任务+7分钟持续工作)的效应好于延迟酝酿(5分钟有意识工作+4分钟分心任务+2分钟继续工作)2但这一结论的得出可能是由于在因变量计算上, 延迟酝酿组涵盖了5分钟准备期工作的成果, 并不是纯粹的酝酿后效应。。建议未来研究考虑进行元分析, 比较两种范式获取的效应量差异。
根据实验范式和因变量测量任务, 权衡合适的准备期长度。Sio等人(2009)对延迟酝酿的元分析显示:对于视觉任务, 准备期越长, 被试更可能意识到转换策略或改变问题表征的必要性, 酝酿效应也越大。Strick等人(2011)对无意识与决策的元分析也显示, 呈现目标问题的时间应当足够短, 避免过早地做判断, 立即酝酿才有效应。较长的准备期既可能允许被试穷尽一个问题空间, 从而更可能在第二次尝试时转换问题表征, 但也可能使得固着或定势被建立和增强, 阻碍无意识加工。一种可能的情况是, 对于立即酝酿范式, 应当有较短的准备期长度, 但对于延迟酝酿范式, 至少在使用视觉任务时, 需要足够长的准备期长度。在具体的实验设计中, 建议研究者审慎选取范式和因变量测量任务, 权衡准备期长度, 以获取最大的酝酿效应。
此外, 酝酿效应的产生是否一定需要被试拥有针对目标任务的内隐目标?Snyder等人(2004)利用延迟酝酿范式检测无意识加工的作用, 在实验中没有告知被试在酝酿之后还要回来解决先前的任务, 但是还是出现了酝酿效应; 而Bos等人(2008)的使用立即酝酿范式的研究则显示, 如果被试不知道在分心任务之后还会第二次解决目标任务, 则酝酿效应不会存在, 即无意识思维是目标依存性的。因此, 在实验室环境下, 被试是否需要知道自己在分心任务后会回到目标任务, 或者说是否需要保持一个内隐的目标, 是一个尚未解决的问题。建议今后的研究者对此问题展开研究, 以提高在实验室中复现酝酿效应的可能性。
5.4 发掘促进酝酿效应的积极因素
如何提升酝酿的效果是在实践上极重要的问题。未来的研究可以尝试将酝酿效应和冥想结合。作为一种广受欢迎的实践项目, 冥想训练被认为可以有效促进酝酿效应(Deshmukh, 2013)。例如, Ren等人(2011)让被试在冥想(间隔一定次数的呼吸后举手报告)之后, 再次尝试解决冥想前无法解决的顿悟问题, 结果发现相较于控制组, 冥想组的被试解题率更高。并且发现冥想时的α波占比(与精神放松有关)和冥想后的顿悟负相关。说明在酝酿期进行冥想, 可能使被试保持在一个警醒且专注的状态, 从而促进顿悟。情境改变也可能是促进酝酿产生的积极因素。Smith等人(2020)提出酝酿的情境依存固着假设(context-dependent fixation hypothesis), 认为当个体处于和错误信念无关的新情境下, 固着就可以被打破。考虑到目前选择性遗忘理论对遗忘的理解停留在消退说或干扰说的层面, 但遗忘却并不仅仅是消退或干扰。遗忘也可能是来自于情境的转变。缺乏情境线索的支持, 相关记忆也将更难被提取(Anderson & Hulbert, 2020)。目前的研究大多强调来自分心任务的干扰能促进对误导线索的遗忘, 却忽视了情境的转变也能促进遗忘, 增强酝酿效应。建议未来研究积极寻找提升酝酿效果的积极因素。
5.5 进一步研究心智游移, 睡眠的酝酿效应
对于心智游移, 未来研究需要解决的首要问题是确定心智游移和酝酿之间的关系。酝酿期的心智游移是否有利于问题解决?目前存在互相矛盾的结论, 因此建议今后的研究者探索可能的调节变量。例如, 例如, 过往研究大多强调心智游移的频率, 而忽视心智游移的具体内容的分析, 近年来脑成像研究和行为实验才逐渐开始注意心智游移时具体的意识内容(Andrews-Hanna et al., 2010; Killingsworth & Gilbert, 2010)。对心智游移主题内容的分析可能会揭示一些调节变量, 例如, 是否与目标任务有关的心智游移更能促进酝酿效应的产生?其次, 建议今后的研究者开展对心智游移的认知神经机制研究, 探索默认网络和执行控制网络在酝酿期如何发挥作用。对这一主题的研究将揭示酝酿效应的认知神经机制, 确定酝酿期的认知活动是否全然是自发的无意识思维, 还是存在目标导向的成分, 从而推动酝酿效应理论的进一步发展。
对于睡眠, 未来研究需要解决的首要问题是确定睡眠是否能, 以及如何能促进创造性问题解决。目前有研究者认同睡眠的表征重构说, 也有人认可激活扩散说, 也有人认可线索同化说。研究者需要确定各种理论分别适用于何种情况, 也即寻找调节变量。例如, 被试是整晚睡眠还是仅仅小憩片刻?研究的对象的哪一个睡眠阶段?使用的因变量是侧重表征重构还是激活扩散?其次, 目前很多实验都采用了目标记忆重激活的范式, 建议未来研究明确这一范式下得出的结论和直接对比睡眠组和控制组所得的结论有何差异。最后, 目前非常缺乏对睡眠促进创造力这一主题的脑影像学研究, 建议研究者开展相关实验, 揭示睡眠促进创造力的认知神经机制。
5.6 关注酝酿的非认知层面
酝酿不是纯粹冰冷的认知过程, 而是涉及到复杂的社会因素和情绪因素。之后的研究应当尝试关注群体水平的酝酿效应, 例如, Zhou等人(2019)发现, 在群体性的想法生成活动中, 接触到他人的想法会造成评价恐惧, 增加负面情绪, 而酝酿期可以减少接触他人想法的负面效应, 促进团体创造力。Breslin (2019)的研究显示, 在团队创造过程中, 酝酿期最好可以让全员都进入休息, 而不是只让一部分人休息。关注社会层面的调节因素, 例如, Zhang等人(2018)发现, 尽管很多研究显示提供外部奖励不利于创造力, 但在酝酿期前给予奖励通知反而会促进酝酿效应。关注酝酿过程的情绪因素, 例如Hao等人(2015)发现在酝酿期诱发积极情绪, 相比于诱发中性或负性情绪, 产生最大的酝酿效应。建议今后的研究更加关注群体酝酿, 社会层面的调节因素, 不同情绪对酝酿的影响等, 从这一新视角出发积累新的实证证据。
沈汪兵, 刘昌, 罗劲, 余洁. (2012). 顿悟问题思维僵局早期觉察的脑电研究.(7), 924–935.
唐晓晨, 庞娇艳, 罗劲. (2009). 顿悟中的蔡格尼克效应: 左右脑在解题失败与提示信息加工时的活动差异.(22), 3464–3474.
Allen, A. P., & Thomas, K. E. (2011). A dual process account of creative thinking.,(2), 109–118.
Al–Shorachi, E., Sasasmit, K., & Gonçalves, M. (2015). Creativity intervention: Using storytelling and math problems as intervening tasks for inducing incubation. In(pp. 081–090).
Andrews-Hanna, J. R., Reidler, J. S., Huang, C., & Buckner, R. L. (2010). Evidence for the dsefault network's role in spontaneous cognition., 322−335.
Baird, B., Smallwood, J., Mrazek, M. D., Kam, J. W. Y., Franklin, M. S., & Schooler, J. W. (2012). Inspired by distraction: Mind wandering facilitates creative incubation.,(10), 1117–1122.
Bargh, J. A., Raymond, P., Pryor, J. B., & Strack, F. (1995). Attractiveness of the underling: An automatic power→ sex association and its consequences for sexual harassment and aggression.,(5), 768.
Beijamini, F., Valentin, A., Jäger, R., Born, J., & Diekelmann, S. (2021). Sleep facilitates problem solving with no additional gain through targeted memory reactivation.,, 30.
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S. F., Rao, S. M., & Cox, R. W. (1999). Conceptual processing during the conscious resting state: A functional MRI study., 80−83.
Bos, M. W., Dijksterhuis, A., & van Baaren, R. B. V. (2008). On the goal–dependency of unconscious thought.(4), 1114–1120.
Bos, M. W., Dijksterhuis, A., & van Baaren, R. B. V. (2011). The benefits of “sleeping on things”: Unconscious thought leads to automatic weighting.,(1), 4–8.
Bowden, E. M., & Jung–Beeman, M. (2003). Normative data for 144 compound remote associate problems.,(4), 634–639.
Brandmeyer, T., & Delorme, A. (2020). Meditation and the wandering mind: A theoretical framework of underlying neurocognitive mechanisms., 1745691620917340.
Breslin, D. (2019). Off‐Task social breaks and group creativity.,(4), 496–507.
Brodt, S., Pöhlchen, D., Täumer, E., Gais, S., & Schönauer, M. (2018). Incubation, not sleep, aids problem-solving.,(10), zsy155.
Buckner, R. L., Andrews–Hanna, J. R., & Schacter, D. L. (2008). The brain’s default network anatomy, function, and relevance to disease., 1−38.
Bursley, J. K., Nestor, A., Tarr, M. J., & Creswell, J. D. (2016). Awake, offline processing during associative learning.,(4), e0127522.
Cai, D. J., Mednick, S. A., Harrison, E. M., Kanady, J. C., & Mednick, S. C. (2009). REM, not incubation, improves creativity by priming associative networks.,(25), 10130–10134.
Cerruti, C., & Schlaug, G. (2009). Anodal transcranial direct current stimulation of the prefrontal cortex enhances complex verbal associative thought.(10), 1980–1987.
Chiang, N. C., & Chen, M. L. (2017). Benefits of incubation on divergent thinking.(3), 282–291.
Choi, H., & Smith, S. M. (2005). Incubation and the resolution of tip-of-the-tongue states.(4), 365–376.
Christoff, K., Gordon, A. M., Smallwood, J., Smith, R., & Schooler, J. W. (2009). Experience sampling during fMRI reveals default network and executive system contributions to mind wandering.8719–8724.
Cousins, J. N., El-Deredy, W., Parkes, L. M., Hennies, N., & Lewis, P. A. (2016). Cued reactivation of motor learning during sleep leads to overnight changes in functional brain activity and connectivity.(5), e1002451.
Creswell, J. D., Bursley, J. K., & Satpute, A. B. (2013). Neural reactivation links unconscious thought to decision-making performance.,(8), 863–869.
Darsaud, A., Wagner, U., Balteau, E., Desseilles, M., Sterpenich, V., Vandewalle, G., … Maquet, P. (2011). Neural precursors of delayed insight.(8), 1900–1910.
Deshmukh, V. D. (2013). Cognitive pause-and-unload hypothesis of meditation and creativity.(3), 217.
Dijksterhuis, A. (2004). Think different: The merits of unconscious thought in preference development and decision making.,(5), 586.
Dijksterhuis, A. (2013). First neural evidence for the unconscious thought process.,(8), 845.
Dijksterhuis, A., & Meurs, T. (2006). Where creativity resides: The generative power of unconscious thought.,(1), 135–146.
Dijksterhuis, A., & Nordgren, L. F. (2006). A theory of unconscious thought.,(2), 95–109.
Fox, K. C., Andrews-Hanna, J. R., & Christoff, K. (2016). The neurobiology of self-generated thought from cells to systems: Integrating evidence from lesion studies, human intracranial electrophysiology, neurochemistry, and neuroendocrinology.,, 134–150.
Fox, K. C., & Beaty, R. E. (2019). Mind-wandering as creative thinking: Neural, psychological, and theoretical considerations.,, 123–130.
Fox, K. C., Spreng, R. N., Ellamil, M., Andrews-Hanna, J. R., & Christoff, K. (2015). The wandering brain: Meta-analysis of functional neuroimaging studies of mind-wandering and related spontaneous thought processes., 611–621.
Frith, E., Ponce, P., & Loprinzi, P. D. (2021). Active or inert? An experimental comparison of creative ideation across incubation periods.,(1), 5–14.
Gao, Y., & Zhang, H. (2014). Unconscious processing modulates creative problem solving: Evidence from an electrophysiological study., 64–73.
Gilhooly, K. J. (2016). Incubation and intuition in creative problem solving.,, 1076.
Gilhooly, K. J. (2017). Response: Commentary: Incubation and intuition in creative problem solving.,, 465.
Gilhooly, K. J., Georgiou, G., & Devery, U. (2013). Incubation and creativity: Do something different.,(2), 137–149.
Gilhooly, K. J., Georgiou, G. J., Garrison, J., Reston, J. D., & Sirota, M. (2012). Don’t wait to incubate: Immediate versus delayed incubation in divergent thinking.,(6), 966–975.
Goel, V., & Vartanian, O. (2005). Dissociating the roles of right ventral lateral and dorsal lateral prefrontal cortex in generation and maintenance of hypotheses in set-shift problems.(8), 1170–1177.
Golchert, J., Smallwood, J., Jefferies, E., Seli, P., Huntenburg, J. M., Liem, F., … Margulies, D. S. (2017). Individual variation in intentionality in the mind-wandering state is reflected in the integration of the default-mode, fronto-parietal, and limbic networks.,, 226–235.
Haarmann, H. J., George, T., Smaliy, A., & Dien, J. (2012). Remote associates test and alpha brain waves.,(2), 5.
Hanslmayr, S., Staudigl, T., & Fellner, M. C. (2012). Oscillatory power decreases and long-term memory: The information via desynchronization hypothesis., 74.
Hao, N., Ku, Y. X., Liu, M. G., Hu, Y., Grabner, R. H., & Fink, A. (2014). Enhancing verbal creativity via brief interventions during an incubation interval.,(1), 30–38.
Hao, N., Liu, M. G., Ku, Y. X., Hu, Y., & Runco, M. A. (2015). Verbal divergent thinking facilitated by a pleasurable incubation interval.,(3), 286.
Henok, N., Vallée-Tourangeau, F., & Vallée-Tourangeau, G. (2020). Incubation and interactivity in insight problem solving.,(1), 128–139.
Hołda, M., Głodek, A., Dankiewicz-Berger, M., Skrzypińska, D., & Szmigielska, B. (2020). Ill-defined problem solving does not benefit from daytime napping.,, 559.
Hélie, S., & Sun, R. (2010). Incubation, insight, and creative problem solving: A unified theory and a connectionist model.,(3), 994.
Johannessen, K. B., & Berntsen, D. (2010). Current concerns in involuntary and voluntary autobiographical memories., 847−860.
Kershaw, T. C., & Ohlsson, S. (2004). Multiple causes of difficulty in insight: The case of the nine-dot problem., 3–13.
Killingsworth, M. A., & Gilbert, D. T. (2010). A wandering mind is an unhappy mind., 932.
Kounios, J., Fleck, J. I., Green, D. L., Payne, L., Stevenson, J. L., Bowden, E. M., & Jung-Beeman, M. (2008). The origins of insight in resting-state brain activity.281–291.
Kounios, J., Frymiare, J. L., Bowden, E. M., Fleck, J. I., Subramaniam, K., Parrish, T. B., & Jung-Beeman, M. (2006). The prepared mind: Neural activity prior to problem presentation predicts subsequent solution by sudden insight.882–890.
Leszczynski, M., Chaieb, L., Reber, T. P., Derner, M., Axmacher, N., & Fell, J. (2017). Mind wandering simultaneously prolongs reactions and promotes creative incubation.,(1), 1–9.
Luo, J., & Niki, K. (2003). Function of hippocampus in “insight” of problem solving.(3), 316–323.
Madjar, N., Shalley, C. E., & Herndon, B. (2019). Taking time to incubate: The moderating role of ‘what you do’ and ‘when you do it’ on creative performance.,(3), 377–388.
Mazoyer, B., Zago, L., Mellet, E., Bricogne, S., Etard, O., Houde, O., … Tzourio-Mazoyer, N. (2001). Cortical networks for working memory and executive functions sustain the conscious resting state in man., 287−298.
McCormick, C., Rosenthal, C. R., Miller, T. D., & Maguire, E. A. (2018). Mind-wandering in people with hippocampal damage.,(11), 2745−2754.
Meyer, D. E., & Schvaneveldt, R. W. (1971). Facilitation in recognizing pairs of words: Evidence of a dependence between retrieval operations.,(2), 227.
Milivojevic, B., Vicente-Grabovetsky, A., & Doeller, C. F. (2015). Insight reconfigures hippocampal-prefrontal memories.(7), 821–830.
Morsella, E., Ben-Zeev, A., Lanska, M., & Bargh, J. A. (2010). The spontaneous thoughts of the night: How future tasks breed intrusive cognitions., 641−650.
Moss, J., Kotovsky, K., & Cagan, J. (2007). The influence of open goals on the acquisition of problem-relevant information.,(5), 876.
Moss, J., Kotovsky, K., & Cagan, J. (2011). The effect of incidental hints when problems are suspended before, during, or after an impasse.,(1), 140.
Mullally, S. L., & O'Mara, S. M. (2013). Suppressing the encoding of new information in memory: A behavioral study derived from principles of hippocampal function.,(1), e50814.
Nordgren, L. F., Bos, M. W., & Dijksterhuis, A. (2011). The best of both worlds: Integrating conscious and unconscious thought best solves complex decisions.,(2), 509–511.
Payne, J. D. (2011). Sleep on it!: Stabilizing and transforming memories during sleep.,(3), 272–274.
Poincaré, H. (1910). Mathematical creation., 321–335.
Ren, J., Huang, Z. H., Luo, J., Wei, G. X., Ying, X. P., Ding, Z. G., … Luo, F. (2011). Meditation promotes insightful problem-solving by keeping people in a mindful and alert conscious state.(10), 961–965.
Ritter, S. M., & Dijksterhuis, A. (2014). Creativity—the unconscious foundations of the incubation period.,, 215.
Sahakyan, L., Delaney, P. F., Foster, N. L., & Abushanab, B. (2013). List-method directed forgetting in cognitive and clinical research: A theoretical and methodological review., 131–189.
Sanders, K. E., Osburn, S., Paller, K. A., & Beeman, M. (2019). Targeted memory reactivation during sleep improves next-day problem solving.,(11), 1616–1624.
Sandkühler, S., & Bhattacharya, J. (2008). Deconstructing insight: EEG correlates of insightful problem solving.(1), e1459.
Schönauer, M., Brodt, S., Pöhlchen, D., Breßmer, A., Danek, A. H., & Gais, S. (2018). Sleep does not promote solving classical insight problems and magic tricks.,, 72.
Seifert, M. C., Meyer, D. E., Davidson, N., Patalano, A. L., & Yaniv, I. (1995). Demystification of cognitive insight: Opportunistic assimilation and the prepared-mind perspective. In R. J. Sternberg & J. E. Davidson (Eds.),(pp. 65–124). Cambridge, MA: MIT Press.
Segal, E. (2004). Incubation in insight problem solving.,(1), 141–148.
Seyed-Allaei, S., Avanaki, Z. N., Bahrami, B., & Shallice, T. (2017). Major thought restructuring: The roles of different prefrontal cortical regions.(7), 1147–1161.
Shaw, S. (2020).(Unpublished doctoral dissertation). University of California, Los Angeles.
Shen, W. B., Tong, Y., Li, F., Yuan, Y., Hommel, B., Liu, C., & Luo, J. (2018). Tracking the neurodynamics of insight: A meta-analysis of neuroimaging studies.,, 189–198.
Shen, W. B., Yuan, Y., Liu, C., Zhang, X., Luo, J., & Gong, Z. (2016). Is creative insight task-specific? A coordinate- based meta-analysis of neuroimaging studies on insightful problem solving., 81–90.
Shen, W. B., Yuan, Y., Liu, C., & Luo, J. (2017). The roles of the temporal lobe in creative insight: An integrated review.(4), 321–375.
Sio, U. N., Kotovsky, K., & Cagan, J. (2017). Interrupted: The roles of distributed effort and incubation in preventing fixation and generating problem solutions.,(4), 553–565.
Sio, U. N., Monaghan, P., & Ormerod, T. (2013). Sleep on it, but only if it is difficult: Effects of sleep on problem solving.,(2), 159–166.
Sio, U. N., & Ormerod, T. C. (2009). Does incubation enhance problem solving? A meta-analytic review.,(1), 94.
Sio, U. N., & Ormerod, T. C. (2015). Incubation and cueing effects in problem-solving: Set aside the difficult problems but focus on the easy ones.,(1), 113–129.
Smallwood, J., & Schooler, J. W. (2006). The restless mind., 946–958.
Smallwood, J., & Schooler, J. W. (2013). The restless mind.(S), 130–149.
Smeekens, B. A. (2013).. The University of North Carolina at Greensboro.
Smeekens, B. A., & Kane, M. J. (2016). Working memory capacity, mind wandering, and creative cognition: An individual-differences investigation into the benefits of controlled versus spontaneous thought.,(4), 389.
Smith, C. M., Bushouse, E., & Lord, J. (2010). Individual and group performance on insight problems: The effects of experimentally induced fixation.,(1), 91–99.
Smith, S. M. (1995). Getting into and out of mental ruts: A theory of fixation, incubation, and insight. In R. J. Sternberg & J. E. Davidson (Eds.),(pp.121–149). Cambridge, MA: MIT Press.
Smith, S. M., & Beda, Z. (2020). Old problems in new contexts: The context-dependent fixation hypothesis.,(1), 192.
Smith, S. M., & Blankenship, S. E. (1989). Incubation effects.,(4), 311– 314.
Smith, S. M., & Blankenship, S. E. (1991). Incubation and the persistence of fixation in problem solving., 61–87.
Smith, S. M., & Dodds, R. A. (1999). Incubation. In M. A. Runco & S. R. Pritzker (Eds.),(pp. 39–43). San Diego, CA: Academic Press.
Smith, S. M., Sifonis, C. M., & Angello, G. (2012). Clue insensitivity in remote associates test problem solving.,(2), 3.
Snyder, A., Mitchell, J., Ellwood, S., Yates, A., & Pallier, G. (2004). Nonconscious idea generation.,(3_suppl), 1325–1330.
Steindorf, L., Hammerton, H. A., & Rummel, J. (2020). Mind wandering outside the box—About the role of off-task thoughts and their assessment during creative incubation..
Stickgold, R. (2005). Sleep-dependent memory consolidation.,(7063), 1272–1278.
Storm, B. C., & Angello, G. (2010). Overcoming fixation: Creative problem solving and retrieval-induced forgetting.(9), 1263–1265.
Strick, M., Dijksterhuis, A., Bos, M. W., Sjoerdsma, A., van Baaren, R. B., & Nordgren, L. F. (2011). A meta-analysis on unconscious thought effects.,(6), 738–762.
Tan, T. T., Zou, H., Chen, C. S., & Luo, J. (2015). Mind wandering and the incubation effect in insight problem solving.,(4), 375–382.
Tsenn, J., Atilola, O., McAdams, D. A., & Linsey, J. S. (2014). The effects of time and incubation on design concept generation.,(5), 500–526.
Verleger, R., Rose, M., Wagner, U., Yordanova, J., & Kolev, V. (2013). Insights into sleep's role for insight: Studies with the number reduction task.,(4), 160.
Wagner, U., Gais, S., Haider, H., Verleger, R., & Born, J. (2004). Sleep inspires insight.,(6972), 352– 355.
Wallas, G. (1926).New York, NY: Harcourt Brace.
Watkins, E. R. (2010). Level of construal, mind wandering, and repetitive thought: Reply to McVay and Kane. (2010)., 198−201.
Weisberg, R. W. (2006).. John Wiley & Sons.
Weisberg, R. W. (2013). On the “demystification” of insight: A critique of neuroimaging studies of insight.(1), 1–14.
Yamaoka, A., & Yukawa, S. (2016). Mind-wandering enhances creative problem solving.,(5), 506–512.
Yamaoka, A., & Yukawa, S. (2017). The relationship between mind-wandering or awareness and creativity., 151–162.
Yamaoka, A., & Yukawa, S. (2020). Does mind wandering during the thought incubation period improve creativity and worsen mood?.,(5), 1785–1800.
Yang, H., Chattopadhyay, A., Zhang, K., & Dahl, D. W. (2012). Unconscious creativity: When can unconscious thought outperform conscious thought?.,(4), 573–581.
Yaniv, I., & Meyer, D. E. (1987). Activation and meta-cognition of inaccessible stored information: Potential bases for incubation effects in problem solving.(2), 187–205.
Yuan, Y., & Shen, W. (2016). Commentary: Incubation and intuition in creative problem solving.,, 1807.
Zhang, Z., Zhang, W. T., Wu, X. F., Tan, T. T., & Luo, J. (2019). Incubation optimizes the promoting effects of rewards on creativity.,(2), 271–272.
Zhong, C. B., Dijksterhuis, A., & Galinsky, A. D. (2008). The merits of unconscious thought in creativity.,(9), 912–918.
Zhou, X., Zhai, H. K., Delidabieke, B., Zeng, H., Cui, Y. X., & Cao, X. (2019). Exposure to Ideas, Evaluation Apprehension, and Incubation Intervals in Collaborative Idea Generation.,, 1459.
The incubation effect of creative thinking
LI Ziyi, ZHANG Ze, ZHANG Ying, LUO Jing
(School of psychology, Capital Normal University, Beijing 100048, China)
Incubation effect refers to the phenomenon that when people are confronted with an unsolvable problem, they temporarily put it aside and switch to complete other irrelevant tasks instead, which is beneficial to the ultimate solution of the original problem. In recent years, researchers have conducted extensive studies on incubation effect of creative thinking and its influences, and put forward many theories to explain incubation effect. The representative theories include “selective forgetting theory”, “spreading activation and cue assimilation theory” and “unconscious work theory”. They explain the mechanism of incubation effect from different perspectives and predict different influences and phenomena respectively. Related research of mind wandering and sleep further shed light on the mechanism behind incubation effect. Cognitive neuroscience research has shown that representation change and spreading activation, as two core cognitive processes during incubation period, involve cooperation of multiple brain regions, including prefrontal cortex, striatum, hippocampus, etc. Future research should deepen theoretical construction, reveal the internal mechanism of incubation effect with cognitive neuroscience method and intelligent monitoring devices, so as to understand the nature of incubation effect of creative thinking in a more comprehensive and profound way.
incubation effect, unconscious thought, creativity
B842
2021-06-03
* 国家自然科学基金(31671124, 31871093)、科技创新能力建设-基础科研基金(047-19530050133, 025- 185305000)、北京市科委北京大脑计划资助。
罗劲, E-mail: luoj@psych.ac.cn
