东祁连山高寒草甸植被特征和生物多样性对生境的响应
2022-02-15何国兴刘志刚关文昊乔欢欢张德罡韩天虎潘冬荣柳小妮
李 强, 何国兴, 刘志刚, 关文昊, 乔欢欢, 张德罡, 韩天虎, 孙 斌, 潘冬荣, 柳小妮*
(1.甘肃农业大学草业学院/草业生态系统教育部重点实验室/甘肃省草业工程实验室/中-美草地畜牧业可持续发展研究中心,甘肃 兰州 730070; 2. 甘肃省草原技术推广总站, 甘肃 兰州730000)
草地植被特征是草地生产、生态稳定性调控研究的基础[1]。深入了解草地植被的群落组成、结构和功能,才能使草地的生产、生态功能得以全面发挥[2]。祁连山位于青藏高原的东部边缘,与蒙古高原和黄土高原相邻,是我国西北重要的生态安全屏障、水源涵养和生物多样性维持的生态服务功能区,也是全球气候变化最敏感和生态比较脆弱的地区之一[3]。高寒草甸是祁连山的主要草地类型之一,是高寒区主要的畜牧业生产基地,对草地畜牧业发展具有重要的意义,而且对祁连山地区的生态系统稳定性起着举足轻重的作用[4]。
高寒草甸分布于高海拔地区,生态系统脆弱,抗干扰能力差,植被一旦遭受破坏,靠其自然恢复不仅周期长,而且极为困难[5-6]。近年来,高寒草甸草场发生了严重退化,其生态环境日益恶化[7],严重限制高寒区草地畜牧业的发展和该区域农牧民的生活水平。相关学者基于草地退化的成因等问题开展了大量科学研究发现[8-12],高寒草甸生态系统退化的诱导原因是草场承包后,围栏区块化管理过程中不合理的放牧制度[9],即草场承包围栏区块化管理过程未考虑到地形因素(海拔和坡向)对草地生产力资源的影响,不同地形区域生产力资源下等量载牧加速诱发了高寒草地的退化程度[8]。因此,研究不同地形因素(海拔和坡向)下高寒草甸植被群落结构特征对区域草地群落恢复及其草地管理具有重要意义。此外,草地群落物种多样性是表征草地生态系统稳定性和持续性的一个重要指标[13-14],对不同海拔和坡向下草地物种多样性的研究可以更好地认识草地群落的组成、演变和发展[15],对遏制草地退化和草地畜牧业的发展具有重要意义。
鉴于此,本研究以祁连山东段金强河流域的高寒草甸为研究对象,通过不同海拔和坡向下草地植被的调查,分析其群落特征,功能群、生物多样性的差异性,以期实现草地资源的区块化管理、合理利用和生物多样性保护提供科学的数据参考。
1 材料与方法
1.1 研究区概况
试验地点位于甘肃省祁连山东段金强河流域(102°26′31″~102°55′01″E,37°7′23″~37°17′53″N)(图1),海拔在2 600~4 300 m之间,年均温在-0.1℃~0.6℃间,全年≥ 0℃的积温为1 360℃左右,降雨量446 mm左右,主要集中在7-9月,年均蒸发量范围 1 483~1 614 mm,气候寒冷潮湿,水热同期,平均相对湿度55%,年日照时间2 600 h[16]。草地类型为高寒草甸类,土壤类型为亚高山草甸土,山地黑钙土,山地草甸土,土壤含水量(50%~80%),土壤pH值范围在6.94~8.17之间。
图1 研究区域地理位置Fig.1 Geographical location of the study area
1.2 样地设置和样品的采集
于2020年7—8月,在东祁连山金强河流域选取了7个不同海拔(2 800,3 000,3 200,3 400,3 600,3 800和4 000 m),同一海拔设置2个不同坡向(阳坡、阴坡)样地,每个样地各选取3个10 m×10 m的采样区域(表1)。

表1 不同海拔和坡向高寒草甸样地基本信息Table 1 Basic information of alpine meadow sample points at different elevations and aspect
每个采样区域按对角线设3个50 cm×50 cm的样方,即3个重复。测定每个样方中植被的草层高度(cm)和总盖度(%)及不同种植物的高度和分盖度,然后分种采集地上生物量(齐地面刈割,放入样品袋中),称鲜重后,带回实验室于 105℃烘箱中杀青2 h后,60℃下恒重,称重。
1.3 植被含水率和草地生物多样性的计算
植被含水率的计算公式如下:

(1)
草地生物多样性指数计算公式如下[17]:
物种重要值(Important Value,IV):IV=(A+B+C)/3
(2)
物种丰富度R:R=(S—1)/lnN
(3)
Pielou均匀度指数:E=H′/lnS
(4)
(5)
(6)
IV表示物种重要值,A表示物种相对高度,B表示物种相对盖度,C表示物种相对生物量,S表示样方中物种总数,N表示样方中各物种重要值之和;Pi表示物种i的相对重要值,R表示丰富度,E表示物种均匀度,D表示优势度,H表示物种Shannon-Weiner指数。
1.4 数据处理及分析
采用Excel 2007进行数据整理和绘图。在SPSS21.0中用单因素方差分析、T检验和交互效应分析。相关性分析和聚类分析均使用SPSS21.0进行分析。所有数据均为平均值±标准误。
2 结果与分析
2.1 不同海拔和坡向下高寒草甸植被特征
海拔和坡向的交互效应对植被盖度有显著影响(P<0.05)(表2)。随着海拔升高,植被盖度先升高后降低,阳坡和阴坡均在海拔3 200 m处达最大值。除了海拔4 000 m外,同一海拔阴坡植被盖度均高于阳坡,且随着海拔升高,阴坡与阳坡植被盖度之间差异逐渐减小,海拔2 800 m,3 000 m,3 200 m和3 400 m的阴坡与阳坡植被总盖度之间差异显著(P<0.05)。高寒草甸草层高度和地上生物量变化规律与植被盖度类似。

表2 不同海拔和坡向下高寒草甸植被特征Table 2 Vegetation characteristics of alpine grasslands at different altitudes and aspect
2.2 不同海拔和坡向下高寒草甸植物功能群地上生物量变化特征
海拔和坡向交互效应对莎草科、禾本科、豆科和杂类草地上生物量有显著影响(P<0.05)(表3)。随着海拔升高,莎草科地上生物量呈先升高后降低,阳坡和阴坡均在海拔3 200 m处达最大值;同一海拔,阴坡莎草科地上生物量均高于阳坡,除海拔4 000 m阴坡与阳坡莎草科地上生物量差异不显著外,其他海拔阴坡与阳坡莎草科地上生物量差异显著(P<0.05)。豆科地上生物量变化与莎草科类似。
随着海拔升高,除海拔3 600 m阳坡外,禾本科地上生物量降低,海拔2 800 m阳坡和阴坡的禾本科地上生物量均最高(表3);同一海拔,除海拔2 800 m外,阴坡禾本科地上生物量均高于阳坡。随着海拔升高,杂类草地上生物量呈先升高后降低,阳坡在海拔3 400 m达最大值,而阴坡在海拔3 200 m处达最大值。同一海拔,海拔2 800 m和4 000 m阳坡杂类草地上生物量均低于阴坡,其他海拔阳坡杂类草地上生物量均高于阴坡,且除海拔4 000 m外的其他海拔阴坡与阳坡杂类草地上生物量之间差异显著(P<0.05)。
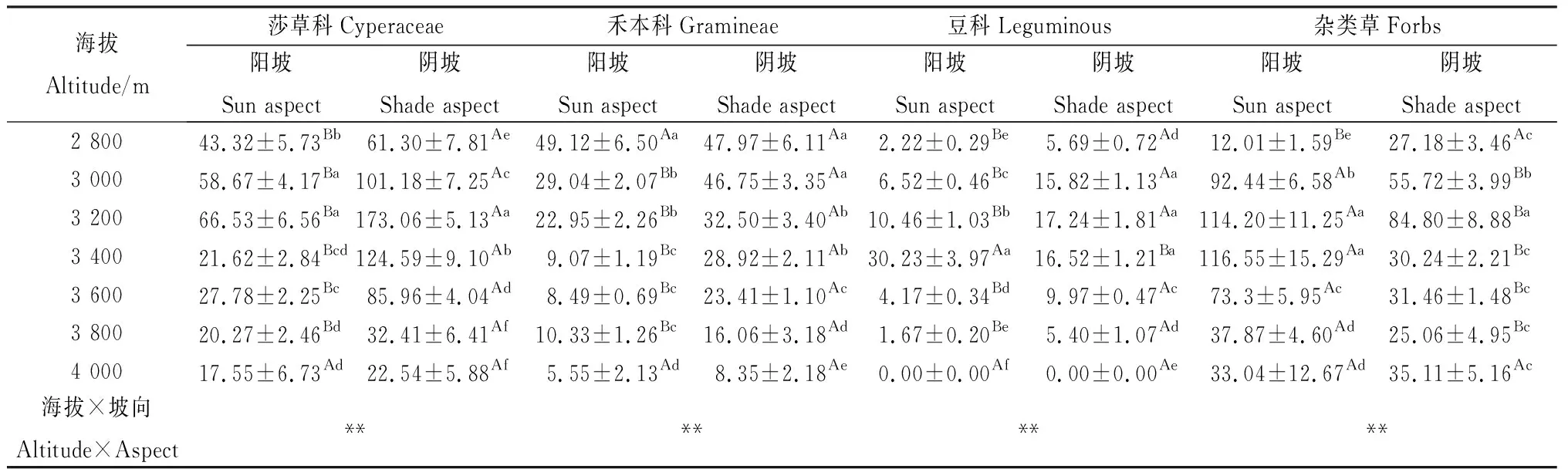
表3 不同海拔和坡向下高寒草甸植物功能群地上生物量的变化特征Table 3 Plant group functional above-ground biomass of alpine grasslands at different altitudes and aspect/g·m-2
从植物功能群地上生物量的组成比例来看(图2),随着海拔升高,阳坡莎草科地上生物量比例先降低后升高,而阴坡莎草科地上生物量比例先升高后降低;禾本科地上生物量比例在阳坡和阴坡均先降低后升高;而豆科地上生物量比例变化趋势与禾本科相反;杂类草地上生物量比例在阳坡呈先升高后降低,在阴坡呈上升趋势。
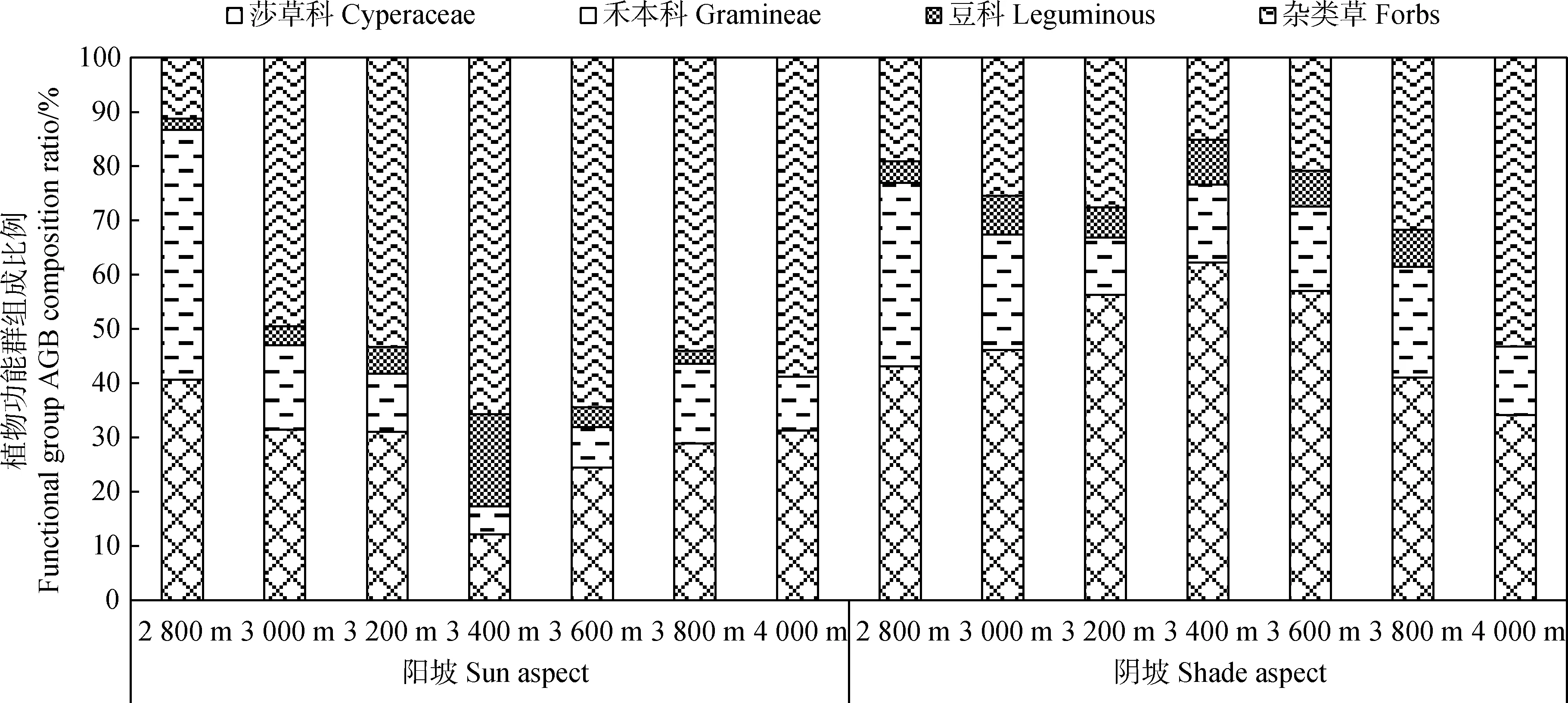
图2 不同海拔和坡向下高寒草甸植物功能群组成比例Fig.2 Plant group functional composition ratio of alpine grasslands at different altitudes and aspect
2.3 不同海拔和坡向下高寒草甸植物含水率
海拔和坡向的交互效应对植被含水率有显著影响(P<0.05)(图3)。随着海拔升高,植被含水率先升高后降低,阳坡和阴坡均在海拔3 200 m处达最大值;同一海拔,除海拔3 600 m外的其他海拔的阴坡植被含水率均高于阳坡,且海拔2 800 m,3 000 m和3 200 m阴坡与阳坡植被含水率之间差异显著(P<0.05)。

图3 不同海拔和坡向下高寒草甸植物含水率Fig.3 Plant water content of alpine grasslands at different altitudes and aspect注:F和Sig代表海拔和坡向交互作用的显著性,当Sig<0.05,即交互效应差异显著;当Sig<0.01,即交互效应极显著。下同Note:F and Sig represent the significance of the interaction between altitude and aspect. Sig<0.05,the interaction effect is significant. Sig<0.01,the interaction effect is extremely significant. The same as below
2.4 不同海拔和坡向下高寒草甸植被群落多样性变化特征
海拔和坡向及其二者的交互效应对Shannon-wiener指数和Simpson指数有显著影响(P<0.05)(图4)。随着海拔升高,Shannon-wiener Index先升高后降低,阳坡在海拔 3 600 m处达最大值,阴坡均在海拔3 200 m处达最大值(图4A);同一海拔,海拔3 200 m以下阴坡Shannon-wiener指数高于阳坡,海拔 3200 m以上阳坡Shannon-wiener指数高于阴坡,除海拔3 200 m和4 000 m外的其他海拔阴坡与阳坡的Shannon-wiener指数之间差异显著(P<0.05)。丰富度指数变化与Shannon-wiener指数基本类似(图4C)。随着海拔升高,除4 000 m梯度外的其它海拔的Pielou指数依次降低(图4B);同一海拔,海拔3 200 m以下阴坡均匀度高于阳坡,海拔3 200 m以上阳坡均匀度高于阴坡。优势度指数变化与Pielou均匀度指数基本类似(图4D)。

图4 不同海拔和坡向下高寒草甸植被生物多样性Fig.4 Vegetation biodiversity of alpine grasslands at different altitudes and aspect
2.5 相关性分析和聚类分析
相关性分析发现,植被盖度与香浓维纳多样性指数、丰富度指数呈极显著正相关,与Pielou均匀度指数呈极显著负相关,与Simpson指数呈显著负相关;草层高度与丰富度呈显著正相关;地上生物量与香浓维纳多样性呈显著正相关,与丰富度指数呈极显著正相关;植被含水率与香浓维纳多样性、丰富度指数呈极显著正相关,与Simpson指数呈极显著负相关。

表4 不同海拔和坡向下高寒草甸植被指标的相关性分析Table 4 Correlation analysis of vegetation indicators in different altitudes and aspect
聚类分析发现(图5),总体呈两类,第一类为:海拔2 800 m阴坡和阳坡、3 600 m阴坡和阳坡,3 800 m阴坡和阳坡4 000 m阴坡和阳坡;第二类为海拔3 000 m的阴坡和阳坡、3 200 m的阴坡和阳坡、3 400 m的阴坡和阳坡。
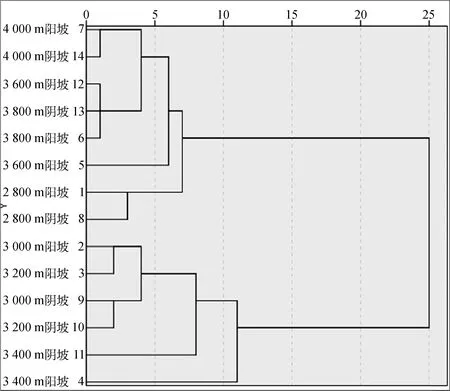
图5 高寒草甸不同海拔和坡向聚类分析Fig.5 Cluster analysis of alpine grasslands at different altitudes and aspect
3 讨论
海拔和坡向是影响高寒草甸植物分布和生长的重要因子,由于海拔和坡向变化导致水热条件的改变形成了环境的异质性分布,进而影响植物群落结构组成差异[18]。金章利等[19]对山地草甸连续3年研究发现,不同海拔草地植被总盖度、草层高度、地上生物量和地下生物量均呈现中海拔>低海拔>高海拔,且不同海拔之间差异显著;王彦龙等[20]研究表明,长江源区高寒草甸地上生物量呈现阴坡大于阳坡的变化趋势。本研究发现,随着海拔的升高,植被总盖度、草层高度、地上生物量呈先升高后降低的趋势;同一海拔上,植被盖度、草层高度、地上生物量基本呈阴坡高于阳坡,这与Busch等[21]和Chick等[22]研究结果一致。随着海拔的升高,平均温度均呈现降低趋势,土壤含水量呈先增加后降低趋势,水热条件的结合促使高寒草甸植被盖度、草层高度和地上生物量呈先升高后降低趋势。当海拔相同时,由于坡向不同导致地面与风向夹角的差异[7],坡向对光、热、水等资源进行了二次分派,从阳坡到阴坡,太阳辐射减少,气温温度降低,蒸发量减少,使阴坡土壤含水量高于阳坡,即草地植被生长条件优于阳坡[6]。本研究还发现,海拔3 200 m为东祁连山金强河流域高寒草甸的典型和中心分布生长区,随着海拔升高的过程中阴坡与阳坡植被盖度、草层高度、地上生物量的差异逐渐缩小,这可能是因为随着海拔的升高,坡向对水热再分派减弱的缘故[21-22]。功能群是植物长期对环境的适应性及某些植物会对生态系统产生相同的作用所形成,并随着外界环境的变化而变化[23]。海拔和坡向对水热的异质性分派,使草地植被功能群地上生物量随着海拔和坡向发生着变化[24]。本研究发现,莎草科、豆科和杂类草地上生物量随海拔升高呈先升高后降低趋势,禾本科地上生物量呈降低趋势;由于从海拔2 800 m到4 000 m,随着水分和热量能级的改变,植物类型发生了变化,依次呈现从旱生型-中旱生型-湿生型的过渡状态,而中旱生型的莎草科、禾本科、豆科等植物生态位较宽,在不同海拔均有分布,即植物对环境的适应性导致植被群落物种组成和结构变化[14]。本研究发现,同一海拔下,阴坡莎草科、禾本科和豆科地上生物量均基本高于阳坡,而杂类草地上生物量阳坡高于阴坡,这与张倩等[25]研究结论类似,但徐长林[26]对坡向对高寒草甸群落植被构成的研究结果与本研究不同,这可能是由于不同科属植物具有不同的适应性、返青期、生长期周期和耐践踏性导致不同坡向植物功能群的差异的缘故[18]。
植物群落物种多样性是生物及其与环境形成的生态复合体以及与此相关的各种生态过程的总和,不仅能够度量群落的组成结构和功能复杂性,而且也能指示环境状况[3]。刘旻霞等[27]对区域物种多样性研究发现,北坡土壤含水量、养分均较高,给植物提供了良好的生长环境,导致北坡的物种多样性高于其他坡向。张倩等[25]研究发现,随着坡向变化,物种丰富度和Shannon-Wiener指数变化一致,其大小均表现为:阴坡>阳坡>半阳坡>半阴坡。本研究中,随着海拔的升高,Shannon-Wiener指数和丰富度指数呈先升高后降低的趋势,Shannon-Wiener指数和丰富度指数基本呈阴坡高于阳坡,而均匀度和优势度指数变化规律与Shannon-Wiener指数基本相反。由于在高海拔和低海拔区域,环境因子不利于优势建群种植物的生长;坡向的改变对小尺度的降雨和地表径流进行分流,不同坡向获取的太阳辐射差异较大,物种组成和原有的生态位发生变化,继而出现新的物种来维持现有的生态平衡,这是生态系统适应环境变化的一种方式[26]。本研究基于聚类分析发现,3 000 m的阴坡和阳坡、3 200 m的阴坡和阳坡、3 400 m的阴坡和阳坡的植被特征、功能群地上生物量和生物多样性指数类似,也就是说在高寒草甸围栏区块化管理过程中,海拔3 000 m和3 400 m可以作为其划分的参考值。
草原是兼有生态和生产两种功能的特殊生态系统[28],任继周先生[29]指出,“草原是一个有生命的开放生态系统单元,应尊重草原自然资源的生态系统特性”。草地承包到户,网围栏区块化管理过程中,部分草场未放牧浪费生产力、部分草场过量载牧导致草场退化,围栏影响了部分野生动物生活习性,矛盾和冲突日益凸显。本试验研究表明,东祁连山金强河流域高寒草甸植被特征、功能群和生物多样性沿海拔和坡向具有显著变化,海拔和坡向共同影响着草地生产力和生物多样性,应依据草地实际生产力状况载畜,调整以山脊线划分家庭牧场的原则,促进草地植被生产力和生物多样性保护耦合协调的可持续发展。
4 结论
高寒草甸植被特征、功能群和生物多样性随着海拔和坡向的改变而变化,同时,海拔与坡向交互作用对植被特征、群落和生物多样性指数的影响差异显著;即海拔和坡向共同影响着高寒草甸草地生产力和生物多样性,海拔3 000 m和3 400 m可以作为东祁连山高寒草甸围栏区块化管理的参考,应避免以山脊作为草场划分的分界线。
